











 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUÇÃO
A biodiversidade funcional - refere-se aos elementos da biodiversidade à escala da exploração agrícola ou da paisagem, que fornecem serviços dos ecossistemas que apoiam a produção agrícola sustentável e podem proporcionar benefícios para o ambiente regional e global e para o público em geral (ELN-FAB, 2012) - é uma importante mais-valia nos sistemas agrícolas ao beneficiar diretamente o agricultor.
Esta biodiversidade funcional pode ser a base de estratégias de gestão do ecossistema que potenciem a componente da limitação natural, reduzindo a necessidade de implementar outras medidas de proteção das culturas, em particular o uso de pesticidas. Diversos estudos mostram que a conservação desta componente da fauna entomológica dos ecossistemas agrícolas apresenta resultados promissores, nomeadamente uma redução clara da pressão de pragas e do uso de pesticidas (ELN-FAB, 2012).
No entanto, os sistemas monoculturais, a intensificação agrícola e o abandono rural são, hoje, responsáveis por grande pressão na biodiversidade dos agroecossistemas, com a consequente perda dos referidos serviços do ecossistema (Kremen e Miles, 2012; Barnes, 2020). Assim, importa conhecer a biodiversidade funcional ao nível da cada agroecossistema, de modo a permitir avaliar a sua importância e contribuir para a sua conservação e promoção, nomeadamente ao nível da proteção da cultura.
A cultura da aveleira não tem expressão significativa, em Portugal: ocupava em 2017 uma área de 357 ha, com uma produção anual de 307 t, com maior importância na região Centro (INE, 2019). No entanto, encontra no território nacional condições favoráveis à sua produção, pelo que representa uma espécie fruteira com interesse e relativamente à qual importa estruturar conhecimento que permita obter um produto de qualidade com base em sistemas de produção sustentáveis.
A biodiversidade de artrópodes associados à cultura da aveleira em Portugal, em particular no que se refere à fauna auxiliar, é pouco conhecida, resumindo-se a listagens das pragas mais relevantes (AliNiazee, 1998; Silva et al., 2005).
Destas, apenas algumas provocam estragos que podem assumir importância económica, como o balanino, Curculio (Curculio) nucum L. (Coleoptera: Curculionidae), afídeos Myzocallis (Myzocallis) coryli Goeze e Corylobium avellanae Schrank (Hemiptera: Aphididae), ácaro-dos-gomos, Phytoptus avellanae Nalepa (Acari: Prostigmata: Eriophyidae) e mineira-da-aveleira, Oberea linearis (L.) (Coleoptera: Cerambycidae) (Silva et al., 2005; Vieira, 2013). De facto, os estragos provocados pelas diversas espécies de artrópodes fitófagos, em pomares velhos, podem atingir cerca de 50% da colheita nas cultivares de casca mais fina, como ’Cosford’ e ‘Daviana’, em particular devido à ação do balanino, ou uma redução na ordem dos 20%, em pomares não tratados e nas cultivares mais sensíveis como ‘Dawton’ e ‘Daviana’, por ação do ácaro-dos-gomos (Silva et al., 2005).
Alguns estudos realizados em países produtores de avelã, indicam que existe um potencial importante associado à fauna auxiliar nesta cultura. Por exemplo, Walton et al. (2009) mostraram que larvas de sirfídeos e crisopídeos dos géneros Chrysopa e Hemerobius têm um papel importante na predação de afídeos e cochonilhas, e diversos outros entomófagos generalistas como carabídeos (predadores do pupas do lepidóptero Cydia latiferreana (Walsingham) e do curculionídeo Curculio occidentis L.), bichas-cadela (como Forficula auricularia L.) e aranhas (como Pimoa altioculata (Keyserling)) que são predadoras de ovos e larvas de diversas espécies de insetos, coccinelídeos (diversas espécies do género Adalia são predadores de afídeos, cochonilhas, outros pequenos insetos e ácaros, sendo bastante eficientes quando existem elevadas densidades de pragas).
Aos longo dos últimos 30 anos, inúmeros estudos realizados em pomares de aveleiras na Turquia (principal produtor mundial) permitiram contruir uma listagem de mais de 150 espécies de insetos auxiliares e 70 espécies de ácaros predadores, com um papel importante na luta biológica contra pragas nesta cultura (Ozman-Sullivan, 2017).
Relativamente ao controlo do cerambicídeo Oberea linearis L. através da luta biológica, Tuncer (2009) identificou diversos insetos parasitóides com interesse - Apanteles spp., Macrosentrus spp., Meteorus spp., Itoplectis spp. e Pristomerus vulnerator (Panzer) - podendo atingir 100% de taxa de parasitismo larvar em alguns anos.
Neste sentido, parece ser importante conhecer a diversidade e a abundância de artrópodes na cultura da aveleira, pelo papel que podem desempenhar na redução dos estragos causados pelas diversas pragas da cultura. Com este objetivo, monitorizou-se, em 2018, ao longo de toda a campanha cultural, um pomar de aveleiras conduzido em modo de produção biológico, na região Centro, de modo a conhecer a biodiversidade de artrópodes nesta cultura e disponibilizar essa informação para apoiar a tomada de decisão na proteção da cultura.
MATERIAL E MÉTODOS
Campo de ensaio
O campo de ensaio consistiu num pomar de aveleiras, em Modo de Produção Biológico desde 2005, com uma área de ≈5500m2, pertencente à Direção Regional de Agricultura e Pescas do Centro e localizado na Estação Agrária de Viseu. A plantação das aveleiras foi realizada em 1989, num solo tipo Al - aluviossolos modernos de textura média a ligeira, derivados de granito, com um compasso de 5 m x 3 m e um sistema de condução em vaso. No campo estão instaladas 15 cultivares de aveleira, cada uma com 6 árvores em cada um de 3 talhões. A envolvência do pomar é constituída a poente por uma bordadura de aveleiras, a nascente por um pomar de macieiras, a norte por um prado e a sul por um souto. A rega é efetuada através de um sistema de micro-aspersão. O solo é revestido com vegetação espontânea natural permanente (enrelvamento natural).
O campo de ensaio foi dividido em 3 talhões (Figura 1), seguindo o alinhamento das 3 repetições de cultivares instaladas, identificadas de 1 a 3, com cerca de 90 árvores cada, tendo-se excluído as árvores de bordadura de cada talhão.
Colheita de artrópodes
O ensaio decorreu de maio a agosto de 2018, sempre que possível com monitorizações quinzenais, pois devido à elevada precipitação que ocorreu durante aquele período nem sempre foi possível realizar as monitorizações previstas.
A amostragem foi realizada de forma aleatória, com recurso à técnica das pancadas, tendo-se utilizado 100 pancadas por talhão, 2 pancadas por árvore.
Os artrópodes foram recolhidos em saco de plástico e imobilizados de imediato com recurso a algodão embebido em acetato de etilo. Posteriormente e já em laboratório, foi realizada a triagem com recurso a microscópio estereoscópico e os artrópodes guardados em frascos, devidamente identificados com data de recolha e talhão. De seguida, os artrópodes capturados foram separados e contabilizados por morfoespécies e identificados até ao táxon possível, seguindo Booth et al. (1990), Triplehorn e Johnson (2005) e Dotson et al. (2011). O uso de morfoespécies como alternativa à classificação taxonómica tem sido utilizado como para superar as dificuldades de identificação, em particular quando se trata de estudos de biodiversidade (Derraik et al., 2010). Baseia-se na separação dos indivíduos por taxum com base em características morfológicas facilmente reconhecíveis (Derraik et al. 2002).
As morfoespécies foram agrupadas em grupos funcionais baseados na informação taxonómica e hábitos alimentares: predadores (organismos que se alimentam de presas), parasitóides [organismos que em estados imaturos se desenvolvem à custa de um organismo de outra espécie (hospedeiro) conduzindo à sua morte], fitófagos (organismos que se alimentam de plantas), detritívoros (organismos que se alimentam de detritos, matéria vegetal morta) e “indiferentes” (organismos sem ação de interesse) (Carlos e Torres, 2009; Coutinho, 2007).
Índices de Biodiversidade
A biodiversidade do agroecossistema em estudo foi avaliada com recurso a diversos índices de biodiversidade, de modo a obter resultados comparáveis em relação à diversidade e riqueza com outras comunidades de insetos ou ao longo do tempo (Campos et al., 2008). Estes baseiam-se na riqueza em espécies, dominância e a equitabilidade de espécies (Quadro 1).
RESULTADOS E DISCUSSÃO
Capturaram-se 1557 artrópodes pertencentes a 9 ordens da classe Insecta e à ordem Araneae da classe Arachnida, sendo a classe Insecta a mais representativa com cerca de 88% dos artrópodes capturados.
Contabilizando por talhão, verificou-se que no talhão 1 se registou maior número de capturas, com 658 artrópodes, seguindo-se o talhão 2 com 473 e o talhão 3 com 426. Ao longo do tempo, houve também variação do número de indivíduos quer por data quer por talhão (Figura 2). No dia 24 de julho, a quantidade de indivíduos capturados foi mais elevada em todos os talhões, o que está relacionado com o aumento da população de afídeos que ocorreu até aquela data, originando um aumento de alimento disponível e um acréscimo da população de indivíduos pertencentes ao grupo predadores e parasitóides.
Figura 2 Número de artrópodes capturados ao longo do ensaio (2018), no pomar de aveleiras da DRAP-C, EA Viseu.
As ordens mais abundantes (Figura 3) foram Hemiptera (54%), com destaque para a grande presença de indivíduos das famílias Aphididae (subordem Sternorryncha), Anthocoridae e Miridae da subordem Heteroptera, e Coleoptera (18%) que apresentou maior número de morfotipos, em particular da família Coccinellidae. É de referir também a presença da ordem Araneae, com 11% de indivíduos capturados. As restantes ordens apresentaram valores residuais. Por data, como já foi referido anteriormente, sobressaiu o dia 24 de julho e, em especial, a ordem Hemiptera (Figura 4).

Figura 3 Distribuição por ordens do total de artrópodes capturados (2018) no pomar de aveleiras da DRAP-C, EA Viseu.
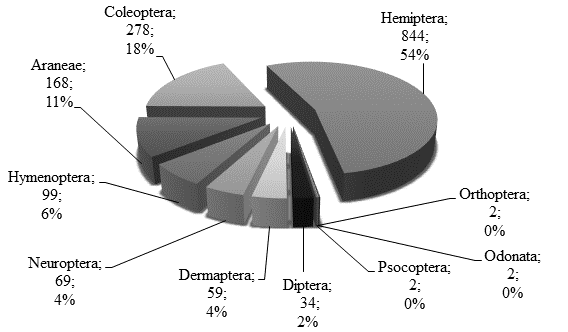
Figura 4 Distribuição por ordens do total de artrópodes capturados ao longo do ensaio (2018) no pomar de aveleiras da DRAP-C, EA Viseu.
Quanto à separação por tipos funcionais, constatou-se que predominam dois grandes grupos: os predadores e parasitóides (66% dos indivíduos capturados) e os fitófagos (33%) (Figura 5). Os restantes, detritívoros e “indiferentes”, representaram um número residual. Por data destaca-se, mais uma vez, o dia 24 de julho com a captura de uma grande quantidade de indivíduos pertencentes ao grupo predadores e parasitóides (Figura 6). Esta situação poderá estar relacionada com a conjugação de fatores ocorrida nessa altura, nomeadamente no que se refere às condições meteorológicas (Figura 6) ocorridas e ao aumento da disponibilidade de alimento (afídeos).

Figura 5 Distribuição dos artrópodes capturados (2018), por grupo funcional, no pomar de aveleiras da DRAP-C, EA Viseu.
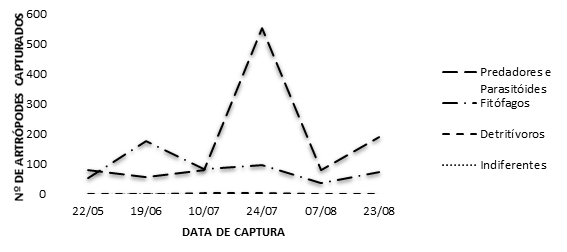
Figura 6 Distribuição dos artrópodes capturados ao longo do tempo (2018) no pomar de aveleiras da DRAP-C, EA Viseu, por grupo funcional, e condições meteorológicas no mesmo período (Fonte destes valores: Estação Meteorológica da Estação Agrária).
Analisando os índices de biodiversidade por morfoespécies (Quadro 2), constatou-se que a riqueza específica variou entre 18 e 21 morfoespécies, revelando um valor de diversidade relativamente baixo quando comparado com estudos realizados em outros países (Ozman-Sullivan, 2017). Em temos de riqueza do ecossistema, o índice de Margalef apresentou-se, ao longo de todo o período de observação, dentro do intervalo de variação que indica a presença de uma diversidade média (superior a 2 e inferior a 5), tendo variado entre 3,09 no dia 24 de julho e 4,10 no dia 22 de maio.
Relativamente à abundância proporcional-dominância, o índice de Simpson foi inferior a 0,33 ao longo de todo o período de monitorização, o que indica uma baixa probabilidade de dois indivíduos serem da mesma espécie, ou seja, reduzida dominância e maior diversidade.
O índice de Shannon-Wiener, que permite avaliar a abundância proporcional-equitabilidade, variou entre 1,79 no dia 24 de julho e 2,48 no dia 22 de maio, que foram sempre inferiores aos valores de lnS (3,04 e 2,89 nas datas referidas, respetivamente), o que indica equitabilidade média - não há dominância, mas as espécies também não estão igualmente representadas.
O índice de Pielou foi, sempre, superior a 0,50, o que pode indicar que, em termos de diversidade, a distribuição das espécies é relativamente uniforme, em particular nos dias 22 de maio e 7 de agosto (0,82 e 0,81 respetivamente).
Quadro 2 Índices de biodiversidade da comunidade de artrópodes, por morfoespécies, ao longo do tempo (2018), no pomar de aveleiras da Estação Agrária de Viseu
| Índices de Biodiversidade | Data | |||||
| 22/05 | 19/06 | 10/07 | 24/07 | 07/08 | 23/08 | |
| Abundância (N) | 131 | 234 | 166 | 648 | 117 | 261 |
| Riqueza específica (S) | 21 | 20 | 21 | 21 | 18 | 21 |
| Índice de Margalef (I) | 4,10 | 3,48 | 3,91 | 3,09 | 3,57 | 3,59 |
| Índice de Simpson (l) | 0,12 | 0,19 | 0,14 | 0,33 | 0,13 | 0,15 |
| Índice de Shannon-Wiener | 2,48 | 2,22 | 2,38 | 1,79 | 2,35 | 2,22 |
| Índice de Pielou | 0,82 | 0,74 | 0,78 | 0,59 | 0,81 | 0,73 |
A mesma análise foi realizada considerando os índices de biodiversidade por ordem (Quadro 3). Neste caso, em termos de riqueza do ecossistema, observa-se uma riqueza específica entre 6 e 9 ordens e um índice de Margalef inferior a 2, o que indica baixa diversidade ao nível das ordens. Em termos de abundância proporcional-dominância, o índice de Simpson apresenta valores inferiores a 0,50, o que indica uma baixa probabilidade de dois indivíduos serem da mesma ordem, ou seja, reduzida dominância.
Os valores do índice de Shannon-Wiener são baixos, o que indica uma equitabilidade baixa, ainda que com uma distribuição das espécies mediamente uniforme - índice de Pielou superior, mas próximo de 0,50.
Quadro 3 Índices de diversidade da comunidade de artrópodes, por ordens, ao longo do tempo (2018), no pomar de aveleiras da Estação Agrária de Viseu
| Índices de Biodiversidade | Data | |||||
| 22/05 | 19/06 | 10/07 | 24/07 | 07/08 | 23/08 | |
| Abundância (N) | 131 | 234 | 166 | 648 | 117 | 261 |
| Riqueza específica (S) | 6 | 6 | 9 | 9 | 7 | 7 |
| Índice de Margalef (I) | 1,03 | 0,92 | 1,56 | 1,24 | 1,26 | 1,08 |
| Índice de Simpson (l) | 0,30 | 0,45 | 0,33 | 0,49 | 0,31 | 0,29 |
| Índice de Shannon-Wiener | 1,38 | 1,08 | 1,39 | 1,16 | 1,51 | 1,45 |
| Índice de Pielou | 0,77 | 0,60 | 0,63 | 0,53 | 0,78 | 0,75 |
Embora a informação obtida seja bastante relevante, seria de esperar maior diversidade e abundância da fauna artrópode, em particular por se tratar de um pomar em modo de produção biológico. Importa relembrar que apenas foram contabilizados os artrópodes susceptíveis de captura com a técnica das pancadas, o que representa apenas uma parte da comunidade de artrópodes presente e se refere apenas a um ano.
Este é um estudo preliminar. É importante continuar a realizar outros trabalhos idênticos, com o intuito avaliar a evolução em anos diferentes, em pomares com outros itinerários técnicos e modos de produção, bem como em outras regiões. Também é essencial prosseguir na identificação das espécies encontradas, avaliação do seu papel funcional e contabilização dos prejuízos causados pelas espécies fitófagas.
CONCLUSÕES
O presente estudo permitiu realizar uma primeira avaliação da biodiversidade de artrópodes na cultura da aveleira, num pomar em modo de produção biológico. Se, por um lado houve alguma variação na abundância de indivíduos ao longo do período de observação, verificou-se um máximo e o mínimo de capturas respetivamente a 24 de julho e 7 de agosto.
Relativamente à avaliação da diversidade, verificou-se uma riqueza ao nível dos morfotipos e ordens relativamente baixa, mas uma equitabilidade média e sem dominância de nenhum grupo, o que pode ser interessante em termos de equilíbrio do ecossistema.
A abundância de insetos auxiliares, principalmente das ordens Coleoptera e Hemiptera-Heteroptera, demonstra um potencial considerável da limitação natural, em particular para a manutenção das populações de afídeos em níveis não prejudiciais à cultura, aspeto bastante importante num pomar conduzido em modo de produção biológica.

