Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Desde la antigüedad, la vid se ha cultivado en la cuenca mediterránea en régimen de secano. En los últimos años, sin embargo, el uso de riego se ha hecho cada vez más habitual en los viñedos mediterráneos, siendo la estrategia predominante la aplicación de riego deficitario en las líneas del viñedo durante el periodo de mayor demanda de agua, paliando así los efectos perjudiciales que el estrés hídrico podría generar sobre el cultivo (Tomás et al., 2012). En el contexto actual de cambio climático, se espera que la superficie dedicada a viñedos en regadío siga creciendo, ya que el aumento de temperaturas y la reducción de las precipitaciones pueden comprometer seriamente la viabilidad de este cultivo en el área mediterránea (Santos et al., 2020). Más allá del aumento de la huella hídrica, la conversión de viñedos de secano en regadío podría tener otros efectos no deseados que deben ser analizados en detalle, como su efecto sobre las comunidades arvenses. En este sentido, se ha demostrado que los sistemas de riego pueden alterar tanto la composición como los rasgos funcionales de las comunidades arvenses presentes en cultivos leñosos (Juárez-Escario et al., 2017). En los viñedos, concretamente, se ha observado en líneas regadas una mayor presencia de malas hierbas perjudiciales para el viñedo (ej., Sonchus asper), viéndose potenciados aquellos rasgos funcionales asociados con una mayor capacidad competitiva de la planta, como la altura o el área foliar (Guerra et al., 2021, 2022). En 2021 se implantó un nuevo ensayo en viñedo con el fin de analizar el efecto de tres estrategias de riego sobre diferentes variables agroambientales. Nuestra hipótesis de partida es que la presencia de especies más competitivas podría ser mayor cuanto mayor sea la dosis de riego aplicada, sobre todo en las líneas del viñedo. En este trabajo presentamos los resultados de la primera campaña de muestreos, durante la cual se ha examinado el efecto de dichas estrategias sobre la composición e identidad funcional de las comunidades arvenses.
MATERIALES Y MÉTODOS
El ensayo se llevó a cabo en la finca experimental El Socorro (IMIDRA; Colmenar de Oreja, Madrid), en una parcela de 0.5 ha dedicada íntegramente a un viñedo en espaldera de la variedad Petit Verdot. El clima de la región es mediterráneo continental semiárido, con una temperatura media anual de 13,7 °C y una precipitación anual de 421 mm (datos de la estación meteorológica de El Socorro para el periodo 2001-2021). El ensayo se estableció en 2021, siguiendo un diseño experimental de bloques al azar con tres repeticiones y tres estrategias de riego, manteniendo en cada estrategia diferentes coeficientes de cultivo (KC) en comparación con la evapotranspiración de referencia (ET0): Kc de 0.2 (riego insuficiente), 0.4 (dosis convencional en nuestra área de estudio) y 0.8 (riego excesivo). El periodo de riego comenzó a principios de junio y finalizó a finales de septiembre, ajustando semanalmente los aportes de riego por goteo para cada estrategia. El manejo del suelo en los tres tratamientos fue el mismo: laboreo en las calles del viñedo y una combinación de laboreo con intercepa y glifosato en las líneas. Con el fin de evaluar el impacto del riego sobre las comunidades arvenses, en 2023 se llevaron a cabo cuatro de muestreos de vegetación en dos momentos diferentes del año: primavera (abril, mayo) y verano (julio, agosto). En cada uno de los 9 plots experimentales (3 bloques × 3 tratamientos) se establecieron 6 puntos de muestreo (3 en la calle, en la línea). Se distinguió entre calle y línea al considerar que podrían existir diferencias en la composición de las comunidades arvenses según la posición que ocupan dentro del viñedo, tal y como observamos en Guerra et al. (2022). En cada punto de muestreo se colocó un marco rectangular de 33 x 66 cm, estimando el porcentaje de cobertura de cada especie presente dentro del marco. Para realizar el tratamiento y análisis de los datos, se seleccionaron en primer lugar aquellas especies con una presencia significativa en la parcela de estudio (porcentaje de cobertura relativa superior a un 1% en alguna de las dos épocas de muestreo). Para dichas especies, a partir de bases de datos como TryDatabase (Kattge et al., 2020), se recopiló información sobre tres rasgos vegetativos relacionados con la capacidad de las especies para competir, captar recursos y/o persistir ante el manejo agrícola: forma vital de Raunkiaer (RLF), altura de la planta (PH) y área foliar (LA). A partir de estos datos se analizó la identidad funcional, calculando para ello la media ponderada de la comunidad o CWM (Garnier et al., 2004) para cada rasgo individual. Para examinar si había diferencias significativas en la composición de las comunidades arvenses según los factores estudiados, se ejecutó un análisis PERMANOVA (Anderson, 2001) tal y como se describe en detalle en Guerra et al. (2022). Paralelamente, se construyeron modelos lineales para analizar la distribución de las especies más abundantes y la cobertura total de especies arvenses en función de los diferentes factores. Además del tratamiento (estrategia de riego), en todos los modelos construidos se usaron como variables predictoras la posición dentro del viñedo, el bloque, la época de muestreo y el efecto combinado del tratamiento con estas tres últimas variables (ej. tratamiento × bloque). Para seleccionar el modelo que se ajustaba mejor a los datos analizados se empleó el criterio de información de Akaike (AIC). La significación estadística de los modelos se computó mediante test ANOVA, empleando un test de Tukey como prueba post-hoc para la comparación por pares. Todos los análisis estadísticos se realizaron en R (R Core Team, 2020).
RESULTADOS Y DISCUSIÓN
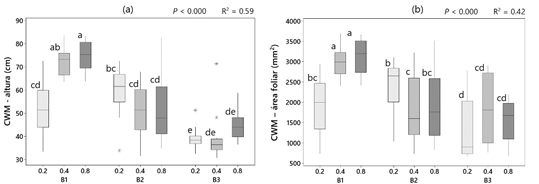
Durante los muestreos de vegetación se identificaron un total de 43 especies, mostrando 14 de ellas una presencia significativa en alguno de los dos periodos de muestreo (Tabla 1). Cirsium arvense (L.) Scop (CIRAR) y Convolvulus arvensis L. (CONAR) fueron las dos especies más abundantes tanto en los muestreos de primavera como en los de verano, tratándose en ambos casos de geófitos de difícil control. Ausente en los muestreos de primavera, cabe destacar también la presencia notable de Solanum nigrum L., mala hierba habitual en cultivos hortícolas de regadío, pero rara en viñedos. El análisis PERMANOVA no evidenció un efecto significativo de la estrategia de riego sobre la composición, la cual se vio afectada fundamentalmente por efectos de bloque y época de muestreo (Tabla 2). No obstante, el mejor modelo se obtuvo al considerar el efecto combinado de tratamiento y bloque. Estos resultados podrían deberse a la presencia mayoritaria de CIRAR y CONAR, las cuáles se expanden fundamentalmente a partir de órganos subterráneos de origen radicular un patrón espacial agregado. En el caso de CIRAR, al diferenciar entre bloques sí se observó un efecto del tratamiento en el bloque 1, siendo significativamente más abundante cuanto mayor es el Kc (Figura 1b). La presencia de SOLNI también se vio afectada por el tratamiento, alcanzando los mayores porcentajes de cobertura en Kc = 0.8 (Figura 1d). En CONAR, sin embargo, no se observó un efecto del tratamiento pero sí de la posición, siendo significativamente más abundante en las líneas del viñedo (Figura 1c), tal y como se señaló en Guerra et al. (2022). En general, al considerar el porcentaje total de especies arvenses el efecto del bloque se diluye y cobra mayor significación tanto el tratamiento como la posición dentro del viñedo. Así, el porcentaje de arvenses fue mayor cuanto mayor fue el Kc, y más alto en las líneas que en las calles del viñedo (Figura 1a).
Tabla 1 Abundancia relativa (según época de muestreo) y rasgos funcionales de aquellas especies arvenses con una presencia significativa en la parcela de estudio. RLF, forma de vida de Raunkiaer (Th, terófito; Th/H, terófito, ocasionalmente hemicriptófito; G, geófito); PH, altura de la planta; LA, área foliar
| ABUNDANCIA RELATIVA (%) | RASGOS FUNCIONALES | |||||
| PRIMAVERA | VERANO | RLF | PH (cm) | LA (mm2) | ||
| BROMA | 3.43 | 0.08 | Th | 21.5 | 132.8 | |
| CIRAR | 29.04 | 37.19 | G | 84.3 | 3718.2 | |
| CONAR | 36.76 | 34.99 | G | 38.2 | 722.4 | |
| CRUBO | 6.99 | 0 | Th/H | 30 | 3974.3 | |
| CRUPY | 3.67 | 0 | Th/H | 65 | 854.8 | |
| DIPER | 1.3 | 3.03 | Th | 42.8 | 2296 | |
| EPHPT | 0 | 7.39 | Th | 8 | 1169.5 | |
| FUMOF | 1.07 | 0.2 | Th | 26.4 | 526 | |
| HORMU | 1.2 | 0.06 | Th | 32.4 | 322.2 | |
| LACSE | 0.91 | 1.51 | Th | 82.2 | 2631.1 | |
| PAPRH | 6.81 | 0 | Th | 43 | 2124.9 | |
| SOLNI | 0 | 10.97 | Th | 46 | 1450.3 | |
| SONAS | 0 | 2.46 | Th/H | 60.6 | 4279.4 | |
| VERHE | 1.98 | 0.12 | Th | 13 | 137.8 | |
Las especies arvenses han sido referenciadas según su código EPPO (2023): BROMA, Bromus madritensis L.; CIRAR, Cirsium arvense (L.) Scop.; CONAR, Convolvulus arvensis L.; CRUBO, Carduus bourgeanus Boiss. & Reut.; CRUPY, Carduus pycnocephalus L.; DIPTE, Diplotaxis erucoides (L.) DC.; EPHPT, Euphorbia prostrata Aiton; FUMOF, Fumaria officinalis L.; HORMU, Hordeum murinum L.; LACSE, Lactuca serriola L.; PAPRH, Papaver rhoeas L., SOLNI, Solanum nigrum L.; SONAS, Sonchus asper (L.) Hill; VERHE, Veronica hederifolia L.
Tabla 2 Resumen de los modelos construidos para el análisis PERMANOVA y para cada variable respuesta: % TOTAL, % cobertura total de especies arvenses; CIRAR, % cobertura de Cirsium arvense; CONAR, % cobertura de Convolvulus arvensis; SOLNI, % cobertura de Solanum nigrum; CWM PH, CWM para el rasgo "altura"; CWM LA, CWM para el rasgo "área de la hoja"; CWM RLF (G), CWM para el rasgo "Forma de vida de Raunkiaer - Geófitos". Se indica el nivel de significación para todos los modelos (***, P < 0.001; **, P < 0.01; *, P < 0.05; ns, P > 0.05). Para los modelos que mostraron el mejor ajuste, se indica además el coeficiente de determinación del modelo (R2)
| PERMANOVA | % TOTAL | CIRAR | CONAR | SOLNI | CWM PH | CWM LA | CWM RLF (G) | |
|---|---|---|---|---|---|---|---|---|
| Modelo1 | ns | *** | * | ns | * | ns | ns | ns |
| Modelo2 | * | *** | ns | *** | ** | * | ** | ns |
| Modelo3 | *** | ns | *** | *** | ns | *** | *** | *** |
| Modelo4 | *** | ** | * | ns | *** | ns | ns | * |
| Modelo5 | ns | 0.38 *** | ns | 0.22 *** | * | ns | * | ns |
| Modelo6 | 0.34 *** | ** | 0.62 *** | *** | ns | 0.59 *** | 0.42 *** | 0.21 *** |
| Modelo7 | *** | *** | ** | ns | 0.22 *** | ns | ns | ns |
Modelo1, tratamiento ~ ; Modelo2, posición ~ ; Modelo3, bloque ~ ; Modelo4, época de muestreo ~ ;
Modelo5, tratamiento × posición ~ ; Modelo6, tratamiento × bloque ~ ; Modelo7, tratamiento × época de muestreo ~
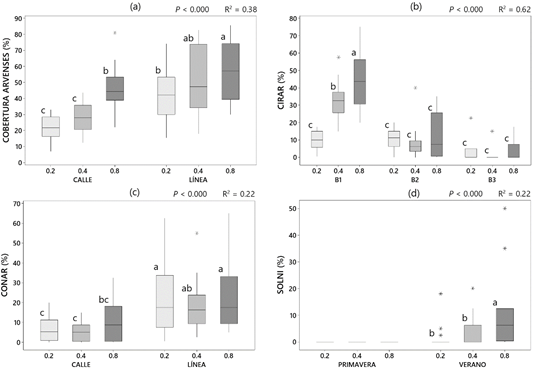
Figura 1 Efecto de los diferentes factores modelizados sobre el porcentaje de cobertura del total de especies arvenses (a), y sobre Cirsium arvense (b), Convolvulus arvensis (c) y Solanum nigrum (d). Letras diferentes sobre los diagramas de cajas indican diferencias significativas entre grupos.