











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Digitaria sanguinalis (L.) Scop. (Ds) es una mala hierba anual con una distribución cosmopolita que puede provocar pérdidas importantes de rendimiento en diferentes cultivos. Para el control de esta mala hierba se utilizan diversos métodos, pero sin duda el uso de herbicidas es el más empleado. La recurrencia de este procedimiento está comportando la aparición de biotipos resistentes a herbicidas (Zhao et al., 2023). El manejo de esta especie, además, se ve dificultado por la posibilidad de la presencia de varias cohortes a lo largo del período de cultivo (Leguizamón & Lovato Echeverría, 2014).
Con la situación indicada, la perspectiva del control biológico de la mala hierba adquiere interés. Son numerosos los organismos que pueden interaccionar con la mencionada especie de la mala hierba (Xia et al., 2020). Entre los hongos patógenos (Krupska, 2012) se encuentra una Ustilaginal (Us - Ustilago syntherismae (Schwein.) Peck). En el inicio de la primavera, el hongo presente en el suelo puede infectar a las plántulas y penetrar en sus tejidos internos. Al desarrollarse la planta con la presencia interna del hongo se pueden producir las siguientes situaciones (Mas & Verdú, 2020): a) las hifas del hongo pueden quedar bloqueadas en alguna zona de la base del tallo principal (plantas asintomáticas que presentaran inflorescencias normales); b) las hifas del hongo se desarrollan más o menos sincrónicamente con el/los tallo/s del individuo (planta que presentará soros en lugar de inflorescencias, totalmente castrada); y c) las hifas del hongo pueden colonizar algún(nos) hijuelo(s), pero no todos (planta que presentará inflorescencias con espiguillas y soros). Las plantas totalmente castradas presentan soros (masas de esporas envueltas por un peridio) que se comportan como unidades de dispersión (Piepenbring et al., 1998). Esta característica abre una ventana a la expectativa del uso del hongo como agente de control de la maleza.
Tanto las plantas asintomáticas como las parcialmente castradas presentan un grado de resistencia, fenotípicamente cualitativa o cuantitativa, respectivamente (Nicks et al., 2015). En campo, no obstante, sólo pueden detectarse a maduración las parcialmente castradas (Tabla 1). Desde el punto de vista de la evolución de la enfermedad, la presencia de plantas parcialmente castradas supone una característica poblacional muy interesante debido a que estos individuos aseguran la producción de semillas y teliósporas.
El objetivo del trabajo es presentar este set de plantas parcialmente castradas y su aptitud para formar tanto soros como inflorescencias con espiguillas.
Tabla 1 Expresión fenotípica de las plantas de Digitaria sanguinalis en su interacción biótica con el hongo Ustilago syntherismae (Verdú & Mas, 2020)
Foto de individuo con espiguillas
|
Foto de individuo con soros
|
Foto de individuos con espiguillas y soros |
| Plantas sanas (1) Plantas asintomáticas (2) | Plantas enteramente castradas (PEC) | Plantas parcialmente castradas (PPC) |
| (1) Pueden presentar resistencia fenotípica cualitativa (2) Pueden presentar resistencia fenotípica cuantitativa | Plantas susceptibles | Pueden presentar resistencia fenotípica cuantitativa |
MATERIALES Y MÉTODOS
El estudio se realizó en una parcela situada en Caldes de Montbui (Barcelona, 41˚36’ N y 2˚10’ E) con un historial agrícola con cultivos de maíz, cebada y girasol, principalmente (Verdú & Mas, 2015). En esta comunicación se presentan los datos correspondientes al período 2009-2017. Al inicio de cada campaña (en mayo) se dispusieron de manera regular y se marcaron en número variable según el año diversos cuadrados de 0.25 m2, con la finalidad de seguir la población de plantas de D.sanguinalis. Semanalmente, se hizo un trabajo de escarda en el interior de los cuadrados para retirar las plántulas/plantas de otras especies. Al final de la campaña (octubre/noviembre) se recolectaron las plantas de cada cuadrado. Para cada individuo se anotó su fenotipo en relación la enfermedad fúngica. Inicialmente se consideraron tres tipos (Tabla 1): sanas (incluye las plantas asintomáticas), parcialmente castradas (PPC, soros e inflorescencias con espiguillas en el mismo individuo), y con presencia absoluta de carbón (soros). En cuatro de los años (2011-2014) del período mencionado se registraron el número de inflorescencias con espiguillas y de soros de cada individuo de los tres tipos de plantas. Para relacionar la frecuencia de plantas parcialmente castradas con la densidad de plantas maduras se utilizó un modelo lineal generalizado con la distribución binomial negativa. El procedimiento GENMOD (SAS, 2013) se utilizó para estimar el modelo. Los estadísticos básicos de las variables se calcularon con el procedimiento UNIVARIATE (SAS, 2013).
RESULTADOS Y DISCUSIÓN
La media de PPC representa un porcentaje muy bajo (2.3%, entre 0 y 6.5% en el período estudiado) del total de plantas presentes en la población que llegaron a la madurez (con espiguillas i/o soros) al final de las campañas estudiadas. La variabilidad en el número observado de PPC en cada campaña se muestra en la Figura 1. En general el tamaño poblacional, número de plantas de los distintos fenotipos, está muy condicionado por la precipitación incidente en el período, y muy en particular la que se registra en el inicio de la primavera y en el inicio del otoño (Verdú & Mas, 2015; Mas & Verdú, 2020).
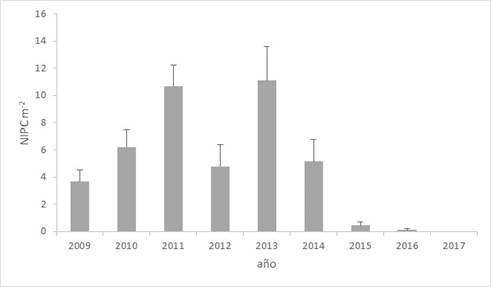
Figura 1 Densidad de individuos parcialmente castrados (NIPC m-2) por Ustilago syntherismae observada en la población de Digitaria sanguinalis estudiada en Caldes de Monbui (Barcelona) al final de la campaña (octubre/noviembre). Las barras representan el error estándar de la media.
El número de PPC presentes en la población está relacionado con el número total de individuos que alcanzan la madurez reproductiva al final de la campaña. Tal y como se puede observar en la Figura 2, el modelo que relaciona estas dos variables pierde fiabilidad a partir de un valor de densidad de plantas superior a 100 individuos/0.25 m2 (en gran medida debido a que se dispone de pocos datos de densidades elevadas).
Las PPC aseguran la entrada en la población de teliósporas que presumiblemente tienen un menor grado de infectividad que el que presentan las de las PEC. Por otro lado, las espiguillas formadas por las PPC tienen semillas que previsiblemente presentaran un mayor grado de resistencia a la infección por el hongo. Este material puede ser de interés para el estudio del genoma del hongo y de la planta implicados en los procesos de infección y de resistencia.
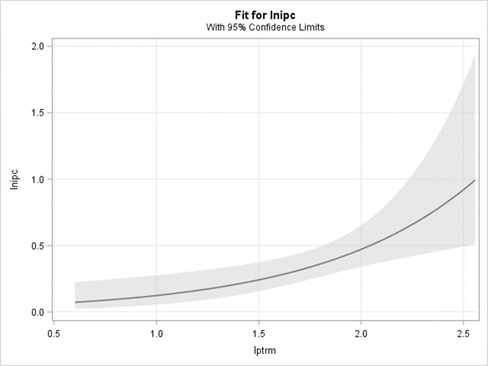
Figura 2 Variación en la densidad de individuos de Digitaria sanguinalis parcialmente castrados (lnipc=log10[nipc*0.25m-2]) por Ustilago syntherismae en función de la densidad de plantas totales reproductivamente maduras (lptrm=log10[ptrm*0.25m-2]) al final de la campaña, en el período 2009-2017 en una parcela de Caldes de Montbui, Barcelona). La función ajustada fue dibujada a partir de los parámetros estimados en un modelo linear generalizado con los límites de confianza del 95%.
CONCLUSIONES
Las plantas parcialmente castradas representan la expresión fenotípica de un cierto grado de resistencia a la infección por el hongo.
Las plantas enteramente castradas son una fuente de esporas que pueden ser usadas en laboratorio para realizar infecciones forzadas y disponer de una cantidad importante de este material.
Sería muy interesante la detección de plantas asintomáticas en campo ya que de esta manera se podrán recolectar espiguillas con semillas con un cierto grado de resistencia a la enfermedad. Y ello permitiría el estudio de las bases genéticas de esta resistencia.

