











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
La enzima acetolactato sintasa (ALS) actúa como diana de varias familias de herbicidas como las sulfonilureas. Su mecanismo de acción se basa en la inhibición de la enzima ALS en la ruta de la biosíntesis de aminoácidos ramificados (leucina, isoleucina y valina) (Zhou et al., 2007). La inhibición del enzima ALS puede inducir un cierto nivel oxidativo a través de la producción de especies reactivas de oxígeno (ROS), especialmente peróxido de hidrógeno (H2O2), como respuesta al estrés abiótico (Zabalza et al., 2007; Eceiza et al., 2023). Sin embargo, las plantas han desarrollado mecanismos para hacer frente al estrés oxidativo y el glutatión es una de las moléculas que elimina H2O2. Además de su participación activa en el proceso de desintoxicación de ROS (Noctor et al., 2012), también sirve como sustrato para las glutatión-Stransferasas (GSTs). Las GSTs son un grupo de enzimas multifuncionales que catalizan la desintoxicación directa de compuestos xenobióticos, a través de su conjugación con moléculas de glutatión reducido (GSH) (Gaafar et al., 2022). Por tanto, son capaces de metabolizar ciertos herbicidas y esta conjugación enzimática se ha descrito como un tipo de mecanismo de resistencia non-target-site (NTS, independiente del mecanismo de acción), si bien los mecanismos NTS en la mayoría de los casos de resistencia a inhibidores de ALS descritos hasta el momento, están relacionados con las enzimas citocromo P450 (Rigon et al., 2020). También se ha descrito la implicación de las GSTs como mecanismo NTS frente a inhibidores de ALS (Gaines et al., 2020).
Algunas especies de malas hierbas son más propensas a desarrollar resistencia, como la especie objeto de nuestro estudio Amaranthus palmeri S. Watson que, debido a sus atributos ecológicos y biológicos, presenta poblaciones resistentes a múltiples herbicidas entre ellos los inhibidores de ALS (Ward et al., 2013).
El objetivo de este estudio es mejorar el conocimiento de los procesos metabólicos involucrados en los mecanismos de resistencia NTS a inhibidores de ALS en A. palmeri. Se ha utilizado una población sensible y una resistente y se ha analizado y comparado la fisiología y el metabolismo del glutatión en respuesta a los inhibidores de ALS para determinar la implicación de las GSTs en la resistencia NTS en A. palmeri.
MATERIAL Y MÉTODOS
Material vegetal y tratamientos
Las semillas de las poblaciones de A. palmeri sensibles y resistentes a inhibidores de ALS fueron proporcionadas por la Dra. Mithila Jugulam (Kansas State University, Manhattan, Kansas, EEUU). La población resistente presenta resistencia NTS a sulfonilureas basada en las enzimas del P450 (Nakka et al., 2017a). Las plantas se crecieron en cultivo hidropónico puro con aireación forzada según Fernández-Escalada et al. (2016). Se contaba con 16 contenedores entre ambas poblaciones con aproximadamente 7- 8 plantas. A día 21 de crecimiento de las plantas en estadio de desarrollo definido como BBCH 14.35 y 48 horas antes de aplicar el herbicida, se aplicó un inhibidor de la enzima GST (NBD-Cl) en una dosis de 270 g m.a./ha. Se utilizó como herbicida nicosulfurón 4% (p/v) a una dosis 5 veces la recomendada, que es 60 g m.a./ha, y que fue aplicado de manera individual o con el inhibidor. Asimismo, se dejaron plantas sin tratar (controles) y plantas solo tratadas con el inhibidor. Se realizaron dos repeticiones de cada tratamiento. A los siete días de la aplicación del herbicida se congelaron hojas todas las plantas (-80 ºC) para la realización de las determinaciones analíticas.
Determinaciones analíticas
El contenido del glutatión y sus relacionados se midió mediante electroforesis capilar equipada con un detector de fluorescencia inducida por láser (Zulet et al., 2015). Se determinó el contenido de cisteína (Cys), γ-glutamil-cisteína (GCC), glutatión reducido (GSH) y glutatión oxidado (GSSG). La determinación de la actividad enzimática se llevó a cabo utilizando el método descrito por Peragón & Amores-Escobar (2018) midiendo la variación a 340 nm del sustrato 1-cloro-2,4-dinitrobenceno (CDNB) que se conjuga con la GSH. Se calculó el nivel de transcripción relativo para genes que codifican GSTs de la subfamilia Phi (GSTFs) mediante PCR cuantitativa (qPCR), con cebadores diseñados a partir de la secuencia de Amaranthus hypochondriacus y con una eficiencia de entre 90 y 110%.
Análisis estadístico
Las posibles diferencias entre los valores medios basales (plantas sin tratar) de las plantas sensibles y resistentes se compararon con la prueba t de Student. Para comparar las diferencias entre tratamientos dentro de la misma población se realizó la prueba ANOVA de una vía, seguida de la prueba post hoc de comparaciones múltiples HSD Tukey o T3 Dunnet test según había o no homogeneidad de varianzas. En todos los casos, los análisis estadísticos fueron realizados a un nivel de significancia del 5 % (p-value ≤ 0.05). Los análisis estadísticos y representaciones gráficas se realizaron con RStudio versión 4.3.1.
RESULTADOS Y DISCUSIÓN
Se evaluaron los contenidos de los metabolitos relacionados con el glutatión (Figura 1). Cuando la población sensible es tratada con nicosulfurón, se observa un incremento en la síntesis de los precursores del glutatión, del GSH y del glutatión total en presencia del herbicida, pero este incremento también es significativo en presencia del inhibidor de las GSTs aplicado 48h antes de la aplicación del herbicida. La población resistente presentó unos valores basales de Cys, GSH y ratio GSH/contenido total superiores a la sensible y en ella también se detectó un incremento significativo del contenido de GSH y glutatión total en presencia del herbicida y el inhibidor.
Las hojas de las plantas resistentes presentaron un mayor nivel basal de la actividad GST (Figura 2). En la población sensible, se observó un incremento significativo en la actividad GSTs tras la aplicación individual, a pesar de no ser significativo tras la aplicación conjunta del herbicida e inhibidor de GSTs también se observa un incremento de la actividad GST (Figura 2), mientras que en la resistente no se detectaron cambios.
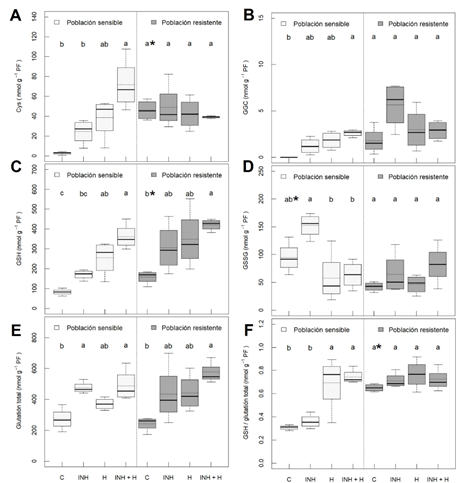
Figura 1 Contenido de glutatión y sus relacionados de dos poblaciones de Amaranthus palmeri (sensible y resistente a nicosulfurón) con diferentes tratamientos ( n = 4) ; plantas sin tratar (C), inhibidor de GSTs (NBD-Cl 270 g m.a./ha) (INH), nicosulfurón (300 g m.a./ha) (H) e inhibidor de GST con nicosulfurón (INH + H). A) Contenido cisteína (Cys), B) contenido de γ-glutamilcisteína (GCC), C) contenido de glutatión reducido (GSH), D) contenido de glutatión oxidado (GSSG), E) contenido del glutatión total (GSH + 2*GSSG) y F) ratio entre el glutatión reducido y el glutatión total (GSH/ glutatión total). Las diferentes letras se refieren a diferencias significativas entre tratamientos de la misma poblacion. Los * se refieren a diferencias significativas entre las plantas sin tratar. Los análisis estadísticos fueron realizados a un nivel de significancia del 5 % (p-value ≤ 0.05).
Las GST Phi y Tau participan en la desintoxicación de herbicidas (Evans et al., 2017). En este estudio se abordó el nivel de expresión génica de dos GSTs de tipo Phi, GSTF2.4 y GSTF2.5. En nuestros resultados no se observaron diferencias significativas para ambas GSTFs ni entre tratamientos (Figura 3, A y B) ni entre poblaciones. Al no haber detectado modificaciones en los niveles de transcritos de estas GSTs se puede concluir que las variaciones detectadas de actividad GST (Figura 2) estarían originadas por cambios postraduccionales o por otro tipo de GSTs.
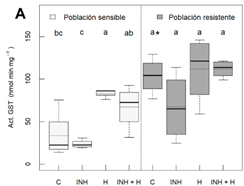
Figura 2 Actividad enzimática GST en dos poblaciones de Amaranthus palmeri (sensible y resistente a nicolsulfurón) con diferentes tratamientos (n = 4); plantas sin tratar (C), inhibidor de GSTs (NBD-Cl 270 g m.a./ha) (INH), nicosulfurón (300 g m.a./ha) (H) e inhibidor de GSTs con nicosulfurón (INH + H). Las diferentes letras se refieren a diferencias significativas entre tratamientos de la misma poblacion. Los * se refieren a diferencias significativas entre las plantas sin tratar. Los análisis estadísticos fueron realizados a un nivel de significancia del 5 % (p-value ≤ 0.05).
Los resultados indican que en la población sensible se induce la síntesis de glutatión en respuesta al estrés oxidativo producido por el herbicida, tal y cómo se ha descrito en otras especies (Zabalza et al., 2007). La inducción de la actividad enzimática GST detectada en la población sensible indica un papel de dichas enzimas en la respuesta fisiológica de la población al herbicida. De hecho, el efecto del herbicida sobre este parámetro se mantiene en presencia del inhibidor de las GSTs.
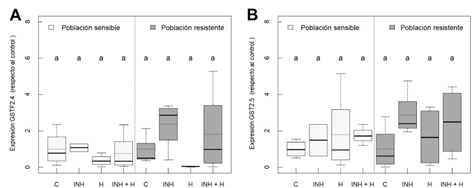
Figura 3 Expresión génica de GSTF2.4 (A) y GSTF2.5 (B) en dos poblaciones de Amaranthus palmeri (sensible y resistente a nicosulfurón) con diferentes tratamientos (n = 4); plantas sin tratar (C), inhibidor de GSTs (NBD-Cl 270 g m.a./ha) (INH), nicosulfurón (300 g m.a./ha) (H) e inhibidor de GSTs con nicosulfurón (INH + H). Las diferentes letras se refieren a diferencias significativas entre tratamientos de la misma poblacion. Los análisis estadísticos fueron realizados a un nivel de significancia del 5 % (p-value ≤ 0.05).
En el caso de la población resistente anteriormente, se detectó resistencia NTS al herbicida atrazina por metabolización de dicho herbicida (Nakka et al., 2017b). Sin embargo, en el caso del nicosulfurón, los resultados de este estudio no confirman la implicación de las enzimas GST en la metabolización de dicho herbicida, ya que a pesar de haber una alta actividad basal en comparación con las plantas sensibles, el nicosulfurón no induce esta actividad.
Estos análisis ayudarán a comprender el metabolismo del glutatión y la implicación de las GSTs en la respuesta de las poblaciones sensibles y resistentes NTS a inhibidores de ALS. El conocimiento de los aspectos fisiológicos de las poblaciones resistentes así como el estudio de los mecanismos de resistencia NTS contribuirá a avanzar en el manejo de dichas poblaciones.

