











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
El glifosato y los inhibidores de la acetolactato sintasa (ALS) son herbicidas que inhiben la síntesis de aminoácidos como mecanismo de acción: el glifosato inhibe la síntesis de aminoácidos aromáticos y los inhibidores de la ALS inhiben la síntesis de aminoácidos ramificados. La 5-enolpiruvil-siquimato-3-fosfato sintasa (EPSPS) y la ALS son enzimas diana muy estudiadas, por lo que el mecanismo de acción de estos herbicidas es bien conocido desde hace tiempo, y son de los herbicidas más usados a nivel mundial. Sin embargo, su modo de acción, conocido como la secuencia completa de eventos que culmina provocando algún daño en la planta que eventualmente puede ser la muerte, no está completamente descifrado.
Hasta la fecha, los estudios dirigidos a conocer el modo de acción del glifosato y los inhibidores de la ALS apuntan a que, a pesar de inhibir enzimas diferentes en rutas metabólicas diferentes, la respuesta fisiológica de la planta a ambos herbicidas es similar. Entre estos efectos comunes están la acumulación de carbohidratos (Orcaray et al., 2010), la disminución de proteína soluble (Zulet et al., 2013) y el estrés oxidativo (Eceiza et al., 2022, 2023). Una familia de enzimas que se ha hipotetizado que puede tener un papel relevante en la respuesta fisiológica a estos herbicidas es la familia de las glutatión S-transferasas (GSTs). Las GSTs son una familia muy amplia de enzimas que, usando el glutatión como sustrato, realizan varias funciones: pueden actuar como antioxidantes, transportadoras... Sin embargo, la función más conocida de las GSTs es la detoxificación de compuestos xenobióticos por conjugación con el glutatión (Noctor et al., 2012). Varios herbicidas pueden ser detoxificados por la GST, incluyendo algunos inhibidores de la ALS (Zhang et al., 2021), mientras que la detoxificación del glifosato por este mecanismo no se ha descrito.
El uso masivo del glifosato y los inhibidores de la ALS ha acarreado efectos medioambientales y económicos poco deseables, como la evolución de biotipos de malas hierbas resistentes. Una de las especies que más biotipos resistentes han desarrollado es Amaranthus palmeri S. Watson, una mala hierba con potencial invasor en la que se han detectado poblaciones resistentes al glifosato, a inhibidores de la ALS, y resistentes múltiples a ambos grupos de herbicidas (Barco-Antoñanzas et al., 2022). En el presente estudio, se han usado poblaciones sensibles, resistentes y resistentes múltiples (todas ellas por mecanismos TS) de Amaranthus palmeri para conocer la implicación de las GSTs en el modo de acción del glifosato y de los inhibidores de la ALS.
MATERIALES Y MÉTODOS
Material vegetal y tratamientos
La resistencia al glifosato de las poblaciones resistente al glifosato (GLR) y resistente múltiple (RM) venía dada por una amplificación génica de la EPSPS (Fernández-Escalada et al., 2016; Barco-Antoñanzas et al., 2022). Las poblaciones resistentes a inhibidores de ALS (AIR) y RM contenían mutaciones en el gen ALS que confiere resistencia cruzada a varias materias activas pertenecientes al grupo de inhibidores de la ALS (Torra et al., 2020; Barco-Antoñanzas et al., 2022; Eceiza et al., 2023). Las plantas fueron crecidas en cultivo hidropónico puro con aireación forzada (Fernández-Escalada et al., 2016). A los 19-22 días de edad se aplicaron los distintos tratamientos (Tabla 1). Se eligieron los inhibidores de ALS que habían seleccionado dichas resistencias. Los herbicidas se aplicaron con un aerógrafo con regulación de presión y volumen. En todas las poblaciones se utilizaron como control plantas sin tratar. Se muestrearon las hojas a los tres días en el caso del glifosato y de siete días en el caso del nicosulfurón.
Tabla 1 Tratamientos aplicados a las distintas poblaciones
| Población | Descripción | Herbicida aplicado | Dosis aplicada Kg ia/ha |
| GLS | Sensible de referencia para GLR | Glifosato | DR = 0,84 kg ha-1 3 DR = 2,52 kg ha-1 |
| GLR | Resistente a Glifosato | ||
| AIS | Sensible de referencia para AIR | Nicosulfurón | DR = 0,06 kg ha-1 3 DR = 0,18 kg ha-1 |
| AIR | Resistente a nicosulfurón | ||
| GLS | Sensible de referencia para RM | Glifosato + Piritiobac | 0,84 kg ha-1 + 0,09 kg ha-1 |
| RM | Resistente múltiple |
Actividad y expresión génica de la GST
La actividad GST se midió espectrofotométricamente como la conjugación del 1-cloro-2,4-dinitro-benceno a 340 nm, como descrito anteriormente (Miyata et al., 2004). En cuanto a la expresión génica, se calculó el nivel de transcripción relativo para genes que codifican GSTs de las subfamilias Phi y Tau mediante PCR cuantitativa (qPCR), con primers diseñados a partir de la secuencia de Amaranthus hypochondriacus y con una eficiencia de entre 90 y 110%.
Análisis estadístico
Para los análisis estadísticos, las plantas se separaron por poblaciones. Las diferencias entre los controles de las plantas resistentes y sus sensibles de referencia (GLR y GLS; AIR y AIS; RM y GLS) se evaluaron mediante la prueba t de Student (p < 0,05). Las diferencias entre tratamientos dentro de una misma población se evaluaron mediante ANOVA de una vía, seguida de una prueba posthoc HSD (p < 0,05).
RESULTADOS Y DISCUSIÓN
La actividad GST basal fue similar en las poblaciones GLS y GLR y en las poblaciones AIS y AIR. Además, en las poblaciones sensibles, se incrementó de forma proporcional a la dosis de glifosato o nicosulfurón (Figura 1). Debido a la falta de evidencias de que la GST pueda degradar el glifosato y a la similitud de la respuesta a ambos herbicidas, es probable que el incremento de la actividad GST no esté implicando una metabolización de los herbicidas, sino que podría ser una respuesta relacionada con el estrés oxidativo causado por estos en las poblaciones sensibles. Esta hipótesis ha sido propuesta anteriormente (Miteva et al., 2010) e iría en línea con estudios previos sobre estrés oxidativo inducido por glifosato o nicosulfurón en estas poblaciones (Eceiza et al., 2022, 2023).
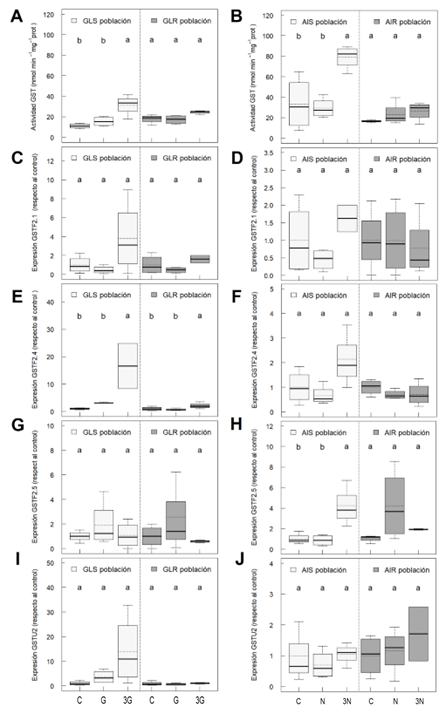
Figura 1 Actividad GST (A, B); expresión génica de GST Phi: GSTF2.1 (C, D), GSTF2.4 (E, F) y GSTF2.5 (G, H); y expresión génica de GST Tau: GSTU2 (I, J), en poblaciones de Amaranthus palmeri sensible a glifosato (GLS), resistente a glifosato (GLR), sensible a inhibidores de ALS (AIS) y resistente a inhibidores de ALS (AIR). Tratamientos con herbicidas: sin tratar (control, C), dosis recomendada de glifosato (G) o nicosulfurón (N), y 3 veces la dosis recomendada de glifosato (3G) o nicosulfurón (3N). Letras diferentes indican diferencias significativas entre tratamientos en cada población (ANOVA, HSD Tukey, p < 0,05).
Las subfamilias de GST Phi y Tau son las más abundantes e importantes en plantas. Se caracterizan por ser inducidas en respuesta a estreses ambientales bióticos y abióticos, como el generado por los herbicidas y, sobre todo en el caso de las Phi, por conferir protección ante el estrés oxidativo (Lee et al., 2014). Se midió la expresión génica de las GST Phi (GSTF2.1, GSTF2.4, GSTF2.5) y Tau (GSTU2). Por lo general, las expresiones génicas de las GST Phi tendieron a incrementarse acorde a la dosis de glifosato o nicosulfurón (Figura 1). Por todo lo dicho, es probable que el incremento de actividad GST se deba, mayoritariamente, a un incremento de las GST Phi en respuesta al estrés oxidativo inducido por ambos herbicidas en las poblaciones sensibles (Eceiza et al., 2022, 2023).
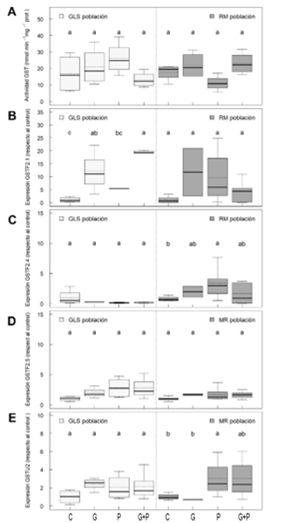
Figura 2 Actividad GST (A); expresión génica de GST Phi: GSTF2.1 (B), GSTF2.4 (C) y GSTF2.5 (D); y expresión génica de GST Tau: GSTU2 (E), en poblaciones de Amaranthus palmeri sensible a glifosato e inhibidores de ALS (GLS) y resistente múltiple a ambos herbicidas (RM). Tratamientos con herbicidas: sin tratar (control, C), dosis recomendada de glifosato (G) o piritiobac (P), y combinación de dosis de glifosato y piritiobac (G+P). Letras diferentes indican diferencias significativas entre tratamientos en cada población (ANOVA, HSD Tukey, p < 0,05).
Para corroborar este efecto de los herbicidas en la actividad y expresión GST, la población GLS (que también se comprobó como sensible a los inhibidores de la ALS) se comparó con la población RM (Figura 2), resistente a glifosato y al inhibidor de la ALS piritiobac por mecanismos target-site (Barco-Antoñanzas et al., 2022). Al igual que en las poblaciones GLR y AIR, los herbicidas no indujeron un incremento de la actividad GST en esta población, tendencia que sí se observó en la población GLS (Figura 2A). Nuevamente, las expresiones génicas tendieron a aumentar con los herbicidas en ambas poblaciones.
CONCLUSIÓN
Tanto el glifosato como los inhibidores de la ALS provocaron, en las plantas sensibles, un incremento de la actividad GST y de expresión génica, sobre todo de las Phi GSTs proporcional a la dosis de herbicida. En las plantas resistentes target-site, en cambio, los ligeros cambios en las expresiones génicas no se tradujeron en mayor actividad GST, evidenciando que la inducción de esta actividad enzimática es un efecto de la inhibición de la EPSPS o la ALS.

