











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Una de las frases más recurrentes dentro del mundo del estudio de las especies invasoras es: “las invasiones biológicas son la segunda causa de extinción de especies y pérdida de biodiversidad a lo largo del planeta después de la destrucción de hábitats” (Mack & D’Antonio, 1998). Aunque es evidente y conspicuo el efecto en la diversidad y biodiversidad de las especies invasoras, ciertamente, esta aseveración no deja de ser una referencia circular en la que no existen datos suficientes para considerarlo un impacto tan dramático como el propuesto. Básicamente, tal como han puesto de manifiesto algunos autores (Theodoropoulus, 2003), estaríamos ante una extrapolación de casos particulares, especialmente en islas, donde verdaderamente el impacto de las exóticas ha podido ser dramático a nivel de provocar cambios de ecosistemas, como es el caso de la introducción de Myrica faya Aiton en Hawaii (Vitousek et al., 1987) o el de depredadores vertebrados que tanto impacto han podido tener en la fauna de los ecosistemas insulares, más vulnerables cuanto más aislados.
La construcción de nuevas carreteras se ha convertido en una de las formas más importantes de dispersar especies, aumentando la homogeneización de las comunidades vegetales, especialmente en la frontera, donde el efecto perturbador del corredor antropogénico (la carretera o sendero) es más evidente (Arévalo et al., 2008; Arteaga et al., 2009). Este proceso destruye hábitats nativos y crea nuevos hábitats para especies con mayor adaptación a entornos perturbados por la actividad humana. Esta homogeneización puede llevar a la extinción local de muchas especies y a la sobredispersión de especies introducidas y generalistas, resultando en una falta de biodiversidad global a diferentes niveles (McKinney & Lockwood, 2001).
En este estudio, hemos seleccionado 12 especies (“dirty dozen”) de las invasoras de Canarias, de las más agresivas, las cuáles usaremos como indicadoras de vulnerabilidad de los distintos hábitats. La distribución de estas especies a lo largo de las carreteras de costa a cumbre (de 0 m a 2450 m el mayor gradiente que hemos encontrado), nos indicará qué hábitats son más vulnerables.
Nuestra hipótesis de partida es que los ecosistemas altamente ligados a la agricultura (en explotación o abandono), son los más adecuados no solo para la producción agrícola, sino los más ricos en especies, y por tanto los más adecuados para el establecimiento y dispersión de especies exóticas.
MATERIALES Y MÉTODOS
Zona de estudio
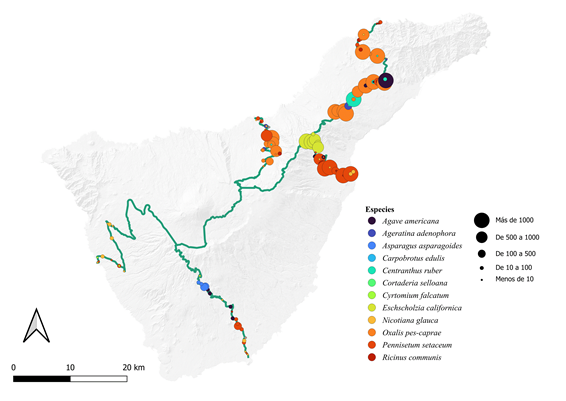
En nuestro caso hemos utilizado la Isla de Tenerife en las Islas Canarias, y de ella hemos seleccionado cinco carreteras desde la costa hasta la máxima altitud que llegan a alcanzar. El muestreo se ha realizado durante los meses de noviembre y diciembre de 2023. De las cinco, dos de ellas parten de la zona suroeste de la isla, dos de la zona norte, y una última recorre prácticamente toda la dorsal de la isla que separa barlovento de sotavento. Las plantas seleccionadas que alcanzaron mayor cota altitudinal, en cada transecto, fueron: Nicotiana glauca Graham (748 m en el transecto a), Oxalis pes-caprae L. (1489 m en el transecto c) y Eschscholzia californica Cham. in Nees (1500 m en el transecto b, 1620 m en el transecto e y 1719 en el transecto d) (Figura 2).
Metodología
Se recorrieron las cinco carreteras seleccionadas desde la menor cota altitudinal posible hasta la mayor altitud. Inicialmente se iba parando y realizando anotación de la presencia de alguna de las especies que tenemos seleccionadas, cada 100 metros de altitud, pero si entre el trayecto encontrábamos alguna, también se anotaban. En todos los casos recogíamos los siguientes datos: densidad de la especie entre 5 clases (menos de 10, entre 10 y 100, de 100 a 500, de 500 a 1000 y más de 1000) y estado fenológico (vegetativo, flor, fruto, flor/fruto).
Para los datos de caracterización de la zona tomábamos datos de altitud, orientación de la carretera tomando como origen el punto de partida de la carretera a nivel del mar, pendiente en la zona donde se encontraba la especie, naturaleza del sustrato (roca, piedra y arena), vegetación, y cobertura arbórea (así como altura y especie dominante en caso de zona forestal). También se caracterizaba el tipo de comunidad vegetal y el nivel de perturbación. Finalmente se anotaban observaciones generales y de la población, y se tomaban fotografías del ambiente y de la planta.
RESULTADOS Y DISCUSIÓN
De las especies seleccionadas, las 12 han aparecido en el estudio, indicando una presencia dominante en los ecosistemas de la isla. Por otro lado, la especie que ha aparecido a mayor altitud ha sido Eschscholzia californica, una especie caracterizada por ambientes de alta montaña. Aún así no pasó de los 1720 metros de altitud. La dominante a menor altitud (10 metros) fue Nicotiana glauca, característica componente del matorral costero e incluso suele aparecer en el cinturón halófilo costero. Tres especies tan sólo aparecieron una vez, en algunos casos ligados a la jardinería (Carpobrotus edulis (L.) N. E. Br. in E. Phillips, Cortaderia selloana (Schult. et Schult. f.) Asch. et Graebn. y Puchlea ovalis (Pers.) DC.).
Representando la altitud media de aparición de las especies, podemos comprobar que la mayoría aparece a la altitud de 200 y 800 metros. Ello vendría a indicar que el fuerte filtro ambiental que supone la altitud es un condicionante importante para la dispersión de las especies exóticas. Esto ya ha sido demostrado en multitud de trabajos previos (Pauchard et al., 2009), y aunque muchos indican una relación con los procesos asociados al calentamiento global (Iseli et al., 2023), todo indica que puede ser más un proceso de aumento de llegada de propágulos (Arévalo et al., 2017).
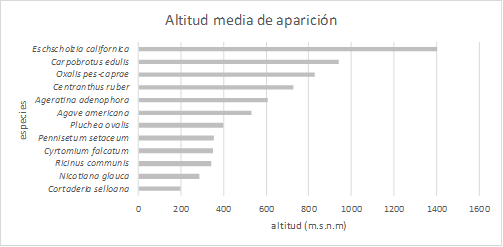
Figura 1 Altitud media (en metros sobre el nivel del mar) de aparición de las especies estudiadas a lo largo de las 5 carreteras estudiadas.
La descripción de la distribución y abundancia de las especies a lo largo de las carreteras quedan reflejadas en la Figura 2. De ellas se pueden extraer varias conclusiones. Las carreteras con orientación suroeste presentan menor abundancia de especies. Las dos carreteras identificadas como tales (a y b), sobre todo la más orientada al oeste, además de poca abundancia también presenta una riqueza menor. Ello indica que las condiciones ambientales son menos adecuadas, al ser una zona que en las partes bajas podríamos decir que guardan condiciones desérticas (menos de 200 mm de precipitación al año), además de elevadas temperaturas con respecto a otra parte de la isla.
Las carreteras que salen de norte o sureste (c, d y e) muestran abundancias muchos mayores de todas las especies que aparecieron, además de una mayor presencia de especies. Las condiciones ambientales de mayor humedad y de temperaturas menos extremas en el periodo estival, ofrecen unas condiciones más apropiadas para la colonización de las islas.
Con respecto al tipo de comunidad, dada la localización de aparición, podemos decir que la más vulnerable a este tipo de colonizaciones coincide directamente con el matorral termófilo, siendo también relativamente vulnerables las zonas de matorral costero y tabaibal cardonal. Sin embargo, como hemos podido ver anteriormente, por encima de los 1700 metros la presencia de especies introducidas empieza a ser ocasional y con abundancias bastante menores.
La comunidad vegetal más común que ha aparecido es el herbazal y el matorral, siendo menos común las comunidades de sustitución en suelos agrícolas como las de zonas boscosas. Muchos herbazales y matorrales también podrían estar relacionados con abandonos lejanos en el tiempo de la actividad agrícola (hecho que ha afectado a más del 90% de los cultivos de Canarias desde los años 60).
CONCLUSIONES
Podemos considerar, desde un punto de vista de aplicación de estos resultados, que no todos los ecosistemas de la isla responden igual a la llegada de propágulos de especies invasoras. Debemos relacionar la dispersión de estas especies no sólo con el tipo de ecosistema, que en cierto modo explica la presencia de las mismas, sino también con el nivel de perturbaciones. Es necesario indicar que los mayores niveles de perturbación se localizan en zonas de costa y zonas de medianías, cuya vegetación natural corresponden con el matorral costero y el tabaibal cardonal, y a mayor altitud, el bosque termófilo, coincidiendo en el primer y segundo caso al desarrollo turístico y en el tercero, a la agricultura.
Por todo ello, es necesario realizar el seguimiento más exhaustivo de estas zonas, ya que las podemos identificar como las que permitirán el establecimiento de las especies más agresivas en la isla, sino indicar también que cualquier nivel de perturbación será en estas zonas donde tenga un impacto más directo en el favorecimiento de las especies introducidas.

