










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkFinisterra - Revista Portuguesa de Geografia
versão impressa ISSN 0430-5027
Finisterra no.113 Lisboa abr. 2020
https://doi.org/10.18055/Finis17881
ARTIGO
Do século XIX ao XXI: Estudo comparativo da vegetação primitiva pelo olhar de Saint-Hilaire e fitofisionomias atuais
From the 19th to the 21st century: a comparative study of primitive vegetation by Saint-Hilaire and current phytophysiognomies
Du XIXème au XXIème siecles: etude comparative de la vegetation primitive selon Saint-Hilaire aux vegetations actuelles
Del siglo XIX al XXI: estudio comparativo de la vegetación primitiva a través de la vista de Saint-Hilaire y las fitofisionomias actuales
Jussiara Dias dos Santos1, Danielle Piuzana Mucida2, Anne Priscila Dias Gonzaga3, Luciano Cavalcante de Jesus França4, Eduarda Soares Menezes5
1Bolseiro, Universidade Federal dos Vales do Jequitinhonha e Mucuri, Rua Engenheiro Alberto Giesbrecht, 307A, Diamantina, 39100-000, Minas Gerais (MG), Brasil. E-mail: jussisantosgeo123@gmail.com
2Professora Associada II, Universidade Federal dos Vales do Jequitinhonha e Mucuri, Diamantina, Minas Gerais, Brasil. E-mail: danielle.piuzana@ufvjm.edu.br
3Professora Adjunta III, Universidade Federal dos Vales do Jequitinhonha e Mucuri, Diamantina, Minas Gerais, Brasil. E-mail: diaspri@gmail.com
4Doutorando em Engenharia Florestal, Universidade Federal de Lavras, Lavras, Minas Gerais, Brasil. E-mail: lucianodejesus@florestal.eng.br
5Doutoranda em Ciência Florestal, Universidade Federal dos Vales do Jequitinhonha e Mucuri, Diamantina, Minas Gerais, Brasil. E-mail: eduarda_menezs@hotmail.com
RESUMO
O objetivo deste trabalho foi comparar os domínios vegetacionais descritos por Saint-Hilaire, em 1817, às classificações fitofisionómicas atuais para a região entre Diamantina, Serro e Minas Novas, Minas Gerais, Brasil. Elaborou-se um banco de dados a partir da análise de suas obras da vegetação para o trajeto percorrido e realizou-se etapas de campo. Foram elaborados mapas de vegetação atual e cobertura da terra de três caminhos percorridos pelo naturalista. Os domínios vegetacionais matas virgens, catingas, carrasqueiros, carrascos e campos naturais foram descritos para a área de estudo e compararam-se às fitofisionomias atuais são denominadas Florestas Estacionais Semidecíduas e Decíduais, Caatinga Arbórea Aberta; Cerrado Denso, Típico, Ralo e Rupestre; Campo Sujo, Limpo e Rupestre, respectivamente. O naturalista descreveu áreas antropizadas como Capoeiras e Campos Artificiais. As comparações entre descrições antigas e atuais mostraram-se viáveis e relevantes. Estas informações contribuem com dados ecológicos, biológicos e geográficos a subsidiarem investigações científicas e estratégias para a conservação dos recursos naturais dos biomas brasileiros.
Palavras-chave: Cerrado; ecótono; Mata Atlântica; viajante naturalista.
ABSTRACT
The objective of this work is to compare the vegetation domains described by Saint-Hilaire at 1817 to current phytophysionomic classifications and ecotone areas for the region between Diamantina, Serro and Minas Novas, Minas Gerais, Brazil. A database was elaborated from the analysis of his works with localities, species and descriptions of the vegetation strata for the trajectory he traveled and steps he covered in the field. Current vegetation and land cover maps were elaborated using the ArcGIS 10.3.1 software. Three paths were defined from the analysis of the traveller’s works. Vegetation domains such as virgin forest, catingas, carrasqueiros, carrascos and natural grasslands, described by Saint Hilaire for Minas Gerais were described in the study area and compared to the current phytophysiognomies: Semi-deciduous Seasonal Forests, Deciduous Seasonal Forests; Open Arboreal Caatinga; Dense, Typical, Ralo and Rupestrian Cerrado; Dirty, Clean and Rupestrian Grasslands respectively. The naturalist describes anthropic domains called capoeira and artificial grasslands whose vegetation is at different stages of ecological succession. Comparisons between old and current descriptions have proved to be feasible and relevant. This information contributes with ecological, biological and geographical data, subsidizing studies and strategies for the conservation of the natural resources of Biomes.
Keywords: Cerrado; ecotone; Atlantic Forest; naturalist traveler.
RÉSUMÉ
L’objectif de cette étude était de comparer les types de végétations décrites par Saint-Hilaire aux classifications actuelles et aux zones écotones de la région comprise entre Diamantina, Serro et Minas Novas dans l’État de Minas Gerais au Brésil. A partir de l’analyse des Suvres du naturaliste, une base de données des lieux qu’il a couvert a été élaborée et des études de terrain ont eu lieu sur les parcours qu’il a parcouru. Des cartes de la végétation et de la couverture terrestre actuelles ont été réalisées à l’aide du logiciel ArcGIS 10.3.1. Trois voies que le naturaliste a parcourues ont été définies à partir de l’analyse des travaux. Des types de végétation tels que les forêts vierges, catingas, carrasqueiros, carrascos et prairies naturelles, décrites par Saint Hilaire pour l’État de Minas Gerais se trouvent dans la zone d’étude et ont été corrélées aux classifications actuelles appelées: forêts saisonnières semi-décidues; forêts saisonnières décidues; Caatinga Arboréale Ouverte; Cerrado Denso, typique, Ralo et Rupestre; Champ Sujo, Limpo et Rupestre, respectivement. Le naturaliste décrit des domaines humanisés appelés Capoeiras et prairies artificielles, dont la végétation se trouve à différents stades de la succession écologique. Les comparaisons entre les descriptions anciennes et actuelles se sont révélées réalisables et pertinentes. Ces informations contribuent comme données écologiques, biologiques et géographiques, à soutenir la recherche scientifique et les stratégies pour la conservation des ressources naturelles des biomes.
Mots clés: Cerrado; ecotone; Forêt Atlantique; voyageur naturaliste.
RESUMEN
El objetivo de este trabajo fue comparar los dominios de vegetación descritos por Saint-Hilaire, en 1817, con las clasificaciones fito-fisionómicas actuales para la región entre Diamantina, Serro y Minas Novas, Minas Gerais, Brasil. Se elaboró una base de datos basada en el análisis de sus trabajos sobre vegetación para la ruta recorrida y se llevaron a cabo etapas de campo. Se prepararon mapas de la vegetación actual y de la cobertura del suelo de tres caminos recorridos por el naturalista. Los dominios de vegetación de bosques vírgenes, catingas, carrasquizos, carrascos y campos naturales se describieron para el área de estudio y, en comparación con las fito-fisionomías actuales, se denominan Bosques Semideciduos y Caducifolios estacionales, Caatinga Arbórea Abierta; Cerrado Denso, Típico, Drenaje y Roca; Campo Sucio, Limpio y Rupestre, respectivamente. El naturalista describió áreas antropizadas como Capoeiras y Campos Artificiales. Las comparaciones entre descripciones antiguas y actuales demostraron ser factibles y relevantes. Esta información contribuye con datos ecológicos, biológicos y geográficos para apoyar investigaciones científicas y estrategias para la conservación de los recursos naturales de los biomas brasileños.
Palabras clave: Cerrado; ecotono; Bosque Atlántico; viajero naturalista.
I. Introdução
Com a vinda da Família Real para o Brasil, em 1808, a Coroa Portuguesa tornou-se menos restritiva quanto à entrada de estrangeiros no território, permitindo maior número de expedições científicas, em relação aos séculos anteriores (Mucida, Gontijo, Morais, & Fagundes, 2019). Com isso, naturalistas estrangeiros tiveram permissão para transitarem por regiões estratégicas da colónia (Ferreira Neto, 2003). No entanto, esse trânsito continuou condicionado à autorização da Coroa portuguesa.
O olhar analítico de naturalistas sobre a paisagem do Brasil no século XIX é apresentado como livros, nomeadamente literaturas de viagem. Estas obras consistem em relatos de viagem, resultados de expedições, cujas sequências normalmente acompanham rotas realizadas nas viagens. Estas literaturas compreendem descrições e análises sobre flora, fauna, geomorfologia, aspetos antropológicos, sociais, dentre outros, e são importantes fontes primárias de informação (Lopes, Milagres, Piuzana, & Morais, 2011).
No que concerne à vegetação, no início do século XIX, os principais Biomas do Brasil foram identificados em trabalhos de Martius que delineou o primeiro esboço da vegetação da colónia (Kury, 2001; Gonzaga, Piuzana, Machado, & Morais, 2018). Outro exemplo de classificação vegetacional é encontrado em trabalho de Saint-Hilaire, no contexto de Minas Gerais, em obras como o Quadro geográfico da vegetação primitiva na Província de Minas Gerais (Saint-Hilaire, 2011 [1837]).
Estudos de naturalistas como de Saint-Hilaire e Martius foram precursores para identificar fitofisionomias (Coutinho, 2006), e auxiliaram no entendimento de ecótonos, áreas onde diferentes floras se interpenetram, constituindo as transições florísticas ou contactos edáficos e climáticos (IBGE, 2012; Milan & Moro, 2017).
As descrições da vegetação de Saint-Hilaire na porção centro-nordeste do Estado de Minas Gerais, subsidiaram o entendimento das formações florestais, savânicas e campestres do Bioma Cerrado (Walter, 2006) bem como de vegetações estépicas do bioma Caatinga e florestas da Mata Atlântica (IBGE, 2012). Desta forma, as análises e entendimento deste tipo de acervo, especialmente de obras traduzidas, são fonte de informação primária, especialmente na região central do estado, devido a sua riqueza em ouro, diamante, ferro, além da diversidade ecológica e botânica (Mucida et al., 2019).
Neste sentido, este estudo teve como objetivo comparar descrições dos domínios vegetacionais descritos por Auguste de Saint-Hilaire, às classificações atuais do ecótono Cerrado-Mata Atlântica-Caatinga por caminhos entre Diamantina, Serro e Minas Novas, região centro-nordeste do Estado de Minas Gerais, Brasil.
II. Materiais e métodos
1. Caracterização da área de estudo
Este trabalho abrange caminhos que conectam Diamantina, Serro e Minas Novas, mesorregião do Vale do Jequitinhonha, Minas Gerais (fig. 1). Os caminhos foram percorridos por Saint-Hilaire entre abril e novembro de 1817. Diamantina e Serro localizam-se na Serra do Espinhaço Meridional. Já o percurso até Minas Novas, a passar por Guanhães e Peçanha, localiza-se a leste da serra.
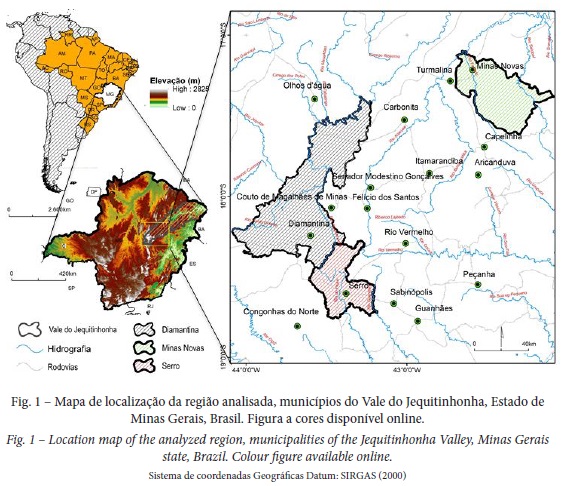
Entre Diamantina e Serro, a área caracteriza-se pelo domínio do Supergrupo Espinhaço que se constitui como planalto, onde afloram rochas das formações basais do Supergrupo Espinhaço (Saadi, 1995). Trata-se de uma região divisora de bacias hidrográficas (São Francisco, Doce e Jequitinhonha), confirmando seu caráter de teto orográfico regional (Saadi, 1995). Entre Serro, Guanhães e Peçanha predominam rochas do embasamento (Bloco Guanhães ou Complexo Ortognáissico de Guanhães) caracterizado por ortognaisses (Noce, Pedrosa-Soares, & Alkmim, 2007). Já entre Minas Novas, Capelinha, Turmalina, Itamarandiba perdominam de rochas metassedimentares neoprotezoicas do Grupo Macaúbas (formações Serra do Catuni, Chapada Acauã) com formações estruturadas de chapadas e grotas (Saadi, 1995).
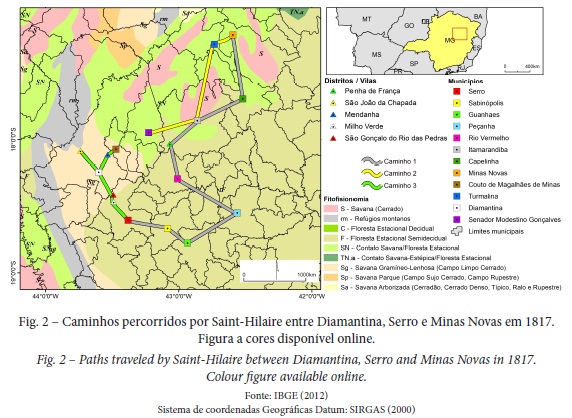
A vegetação encontrada na área de estudo confere-lhe caráter de ecótono; a leste, nas imediações do Serro predomina a vegetação do bioma Mata Atlântica. A oeste, na região de Diamantina, o bioma Cerrado é preponderante, com grande influência das formações campestres. Já em direção a Minas Novas, há transição do Cerrado para Florestas Estacionais Deciduais com influência do bioma Caatinga (fig. 2) (IBGE, 2012).
2. Levantamento de dados
As informações bibliográficas e documentais foram obtidas a partir da base de dados primários das literaturas de viagem. Foram consultadas obras traduzidas para o português do viajante Auguste de Saint-Hilaire, no percurso da região da Serra do Espinhaço e Alto Vale do Jequitinhonha, com destaque para: Saint-Hilaire (2000 [1830]), Saint-Hilaire, (2004 [1833]) e Saint-Hilaire (2011 [1837]). Para o estudo comparativo das vegetações utilizou-se Ribeiro e Walter (2008) e IBGE (2012).
A partir da leitura e análise das obras supracitadas elaborou-se um ficheiro de dados com as localidades percorridas por Saint-Hilaire entre Diamantina, Serro e Minas Novas. As informações priorizadas foram nomes científicos, populares da flora e o tipo de uso do solo à época. As identidades botânicas encontradas foram atualizadas pela lista de espécies da Flora do Brasil (Flora do Brasil, 2018). Excursões a campo foram realizadas com a finalidade de reconhecer marcos geográficos, comparar as características de estratos da vegetação e espécies ocorrentes.
A confeção de uma base cartográfica foi realizada a partir do levantamento de limites territoriais dos municípios referentes aos trajetos de Saint-Hilaire, cujos dados públicos encontram-se disponibilizados no Instituto Prístino (2018) e IBGE (2018). A base de dados da vegetação foi obtida junto ao Ministério de Meio Ambiente (MMA, 2002). O processamento foi realizado pelo software ArcGIS 10.3.1, a partir da sobreposição dos limites (shapefile), compilação com os planos de informações das fitofisionomias e cobertura da terra (ano base 2014). Os caminhos foram elaborados considerando o conceito de distância euclidiana (Ávila, 2001), e as delimitações espaciais vetoriais ao longo de vias de acesso ocorreram a partir das descrições feitas pelo naturalista em 1817.
III. Resultados
1. Os Caminhos percorridos por Saint-Hilaire
Saint-Hilaire percorreu ao longo de 1817, um extenso trajeto da Mesorregião do Jequitinhonha. No Alto Jequitinhonha, nas proximidades de Diamantina, percorreu rotas de forma a visitar importantes serviços de diamantes. As rotas percorridas ao Baixo Jequitinhonha tiveram por objetivo, além análise das paisagens, o estudo das populações de índios Botocudos.
O trajeto de Saint-Hilaire pela área de estudo ocorreu entre 08 de abril e 12 de novembro de 1817 e foi subdividido em (fig. 2): Caminho 1, percorrido em abril, que teve início na sede municipal do Serro (Vila do Príncipe), a passar por Sabinópolis (Vila das Correntes), Guanhães, Peçanha (Passanha), Paulistas, Rio Vermelho, Penha de França, Itamarandiba (São João Batista), Angelândia (Alto dos Bois), Capelinha, Minas Novas (Vila do Fanado). O Caminho 2 iniciado em Minas Novas em meados de julho de 1817, a passar por Itamarandiba, Turmalina (Piedade), Senador Modestino Gonçalves (Vila do Araçuaí). E o Caminho 3 que teve início em novembro de 1817 com trajeto de São João da Chapada (São João do Barro), Diamantina (Arraial de Tijuco), Mendanha, Couto de Magalhães de Minas (Rio Manso), Bandeirinha, São Gonçalo do Rio das Pedras (Rio das Pedras), Capivari (Ribeirão Capivari), Milho Verde e Serro.
2. Descrição da Vegetação no Século XIX por Saint-Hilaire: um Quadro Geral
Ao percorrer a área de estudo, Saint-Hilaire classificou a vegetação por nomes específicos tais como: matas virgens; catingas; carrasqueiros; carrascos; campos naturais, artificiais e capoeiras. Pela narrativa, foi possível separar as vegetações por meio da estratificação vertical e horizontal, assim como pela caracterização da vegetação pela sua composição florística. Sendo assim diferenciamos estruturalmente a vegetação da área de estudo em: matas virgens, que correspondem às plantas mais altas e densas florestas, com mais de 2m de altura; as catingas constituem-se por indivíduos com cerca de 1,5 a 1,8m e diferenciam-se das primeiras por serem caducifólias; carrasqueiros possuem 1,5m e os Carrascos, que por sua vez, são constituídos por indivíduos de cerca de 1m de altura.
Além disso, os campos subdividem-se em artificiais ou naturais (Saint-Hilaire, 2011[1837]). O naturalista descreveu os primeiros como aqueles decorrentes de processos de queimadas e cultivos sucessivos que contribuíam para a expansão de espécies exóticas, como o capim-gordura (Melinis minutiflora) e as samambaias (Pteridium caudata). Já os campos naturais correspondem ao Cerrado Sentido Restrito, Rupestre, Campos Limpo, Sujo e Rupestre, nativos do bioma Cerrado (Ribeiro & Walter, 2008).
2.1. Matas Virgens
O domínio das matas virgens foi relatado nas proximidades de Serro, Peçanha, Angelândia, São Gonçalo do Rio Preto, São Gonçalo do Rio das Pedras e Couto de Magalhães de Minas. Sobre esta vegetação o naturalista afirmou: “As matas pareceram-me apresentar no conjunto da vegetação mais ou menos semelhante à das florestas do Rio de Janeiro; as árvores, todavia, têm, em geral, menos vigor [...]” (Saint-Hilaire, 2000 [1833], p. 134). Uma espécie coletada e frequentemente presente na narrativa foi o jatobá ou jatobá-da-mata (Hymenaea courbaril L.), pertencente à família Fabaceae. Esta espécie é encontrada em Florestas Estacionais do Bioma Cerrado e porções da Floresta Ombrófila da Mata Atlântica (Flora do Brasil, 2018).
Neste sentido, as matas virgens correlacionam-se às formações arbóreas de ocorrência em dois Biomas (Cerrado; Mata Atlântica) e vinculam-se às Florestas Estacionais Semidecíduas (IBGE, 2012; Flora do Brasil, 2018). Cabe destacar que esta fitofisionomia apresenta ampla ocorrência em áreas de ecótono dos biomas descritos para a região (fig. 2, fig. 3A).

2.2. Catingas
Catingas consistem em vegetações intermediárias entre Matas Virgens e Carrasqueiros. São descritas nas proximidades de Minas Novas, Chapada do Norte e Francisco Badaró. Segundo registos da literatura de viagem de Saint-Hilaire (2000 [1833]) ocorrem da seguinte forma: “As catingas acabavam de perder sua folhagem [15/07/1817]. As gramíneas e quase todas as outras plantas que crescem entre as árvores desse bosque estavam torradas pelo ardor do sol” (Saint-Hilaire, 2000 [1833], p. 279). Segundo o naturalista “Se as catingas perdem as suas folhas anualmente, isso vem de que o solo em que elas crescem é, por natureza, mais suscetível de se dessecar do que as terras em que se alteiam as matas virgens” (Saint-Hilaire, 2000, p. 240).
O naturalista relata que as catingas também eram passíveis de desflorestação como as matas virgens, entretanto, para o cultivo de algodão. As áreas, após alguns anos de plantio, eram abandonadas por diminuição de produtividade dando origem aos campos artificiais e capoeiras (fig. 2, fig. 3B). Dentre espécies citadas por Saint-Hilaire para o domínio das catingas estão: Emburana (Commiphora leptophloeos (Mart.) J.B.Gillett). Entretanto, segundo revisão técnica realizada por Juliana de Paula-Souza em Saint-Hilaire (2011, p. 38) a descrição feita pelo naturalista para a espécie C. leptophloeos ainda pode se referir à Amburana cearensis (Allemão) A.C.Sm, também conhecida como Emburana, de ocorrência nas Florestas Estacionais Deciduais da área em estudo (Flora do Brasil, 2018). Outra espécie relatada é a Barriguda (Chorisia ventricosa), basiónimo de Ceiba ventricosa (Nees e Mart.) Ravenna. Ressalva-se que a descrição da planta pelo naturalista também se assemelha às características taxonómicas de Cavanillesia umbellata Ruiz & Pav. espécie típica de áreas com influência do Bioma Caatinga, como a região de Minas Novas e Francisco Badaró.
A Emburana (A. cearenses) e a Barriguda (C. umbellata) apresentam ampla distribuição em áreas ecotonais entre Caatinga e Cerrado, mas com maior aproximação ao primeiro e, frequentes nas Florestas Estacionais Deciduais (Flora do Brasil, 2018). Assim sendo, considerando os aspetos estruturais, florísticos e ecológicos, pode-se inferir que a catinga referida por Saint-Hilaire é atualmente classificada como Florestal Estacional Decidual.
2.3. Carrasqueiros
Os carrasqueiros, na visão do naturalista, desenvolvem-se na região de Minas Novas e são caracterizadas por espécies da família Fabaceae, subfamília Mimosoideae. Em passagem pela região, Saint-Hilaire referiu-se à vegetação:
“Cerca de uma légua [4,2km] de Vila do Fanado (Minas Novas) [...] quando se começa a descer, a terra, sempre vermelha, torna-se, ao mesmo tempo, pedregosa e parece extremamente seca; a vegetação muda completamente de aspeto. Ali, arbustos da altura de seis pés [1,8m] têm os caules retos, bastante delgados e ramificados desde a base; são aproximados uns dos outros; seus ramos se confundem, e o conjunto apresenta exatamente o aspecto das moitas de nossas florestas. Entre esses arbustos, um único apresentava então flores, o chamado flor-de-toda-gente, cujas corolas brancas embalsamavam o ar com o seu doce perfume.” (Saint-Hilaire, 2000 [1833], p. 219).
Considerando os aspetos florísticos descritos pelo naturalista para os carrasqueiros, atribuir equivalência com a atual fitofisionomia é impreciso, haja vista que na descrição, Saint-Hilaire cita apenas ocorrência da família Fabaceae, que apresenta ampla distribuição no território brasileiro, presente nos biomas Caatinga, Cerrado e Mata Atlântica.
No entanto, é possível inferir, considerando juntos os aspetos florísticos e estruturais e a localização geográfica (Minas Novas), que esta vegetação deve possuir carácter ecotonal entre Cerrado e Caatinga, aproximando-se de tipos vegetacionais de menor porte, como a Caatinga Arbustiva Aberta, fitofisionomia dominada pelo gênero Calliandra, uma Mimosoideae (Andrade-Lima, 1981).
2.4. Carrascos
Os carrascos caracterizam-se como vegetação que desenvolvem-se na forma de “florestas anãs, baixas e raquíticas” (Saint-Hilaire, 2000, p. 232). Nestas florestas de pequeno porte podem ser encontradas plantas do gênero Hyptis spp., famílias de Fabaceae e Malvaceae, além de espécies de rosa-do-campo (Kielmeyera rosea Mart. & Zucc.) pertencente à família Calophyllaceae que, para o naturalista, caracterizam o domínio vegetacional. Na área de estudo essa vegetação foi encontrada entre Penha de França, Angelândia, Capelinha, Minas Novas, Turmalina, Itamarandiba e Diamantina. Nas palavras de Saint-Hilaire:
“Encontrei no meio dos carrascos a mesma mimosa, os mesmos Hyptis, finalmente, a composta de folhas urze, que chamam alecrim-do-campo, e que nessa ocasião estava coberta de flores. Nessa época florescia ainda a encantadora malvácea [...] rosa-do-campo; e reconheci que ela devia ser considerada uma das espécies características dos carrascos” (Saint-Hilaire, 2000 [1833], p. 293).
Pelo porte e densidade descritos e espécies mais citadas pelo naturalista é possível associar os carrascos as formações savânicas do Cerrado, a variar entre Cerrado Denso, Cerrado Típico, Cerrado Ralo e Cerrado Rupestre (fig. 2, fig. 3C). Essa correlação relaciona-se à presença de alumínio e elevada acidez do solo, que condiciona o baixo porte da vegetação, inclinadas, tortuosas, com ramificações irregulares e retorcidas (Ribeiro & Walter, 2008).
2.5. Campos Naturais
Saint-Hilaire (2000) descreveu o gênero Hyptis e famílias Myrtaceae, Convolvulaceae, Verbenaceae, Melastomataceae e Ericaceae como plantas predominantes nas áreas abertas de campos naturais. O naturalista relata em Penha de França (distrito de Itamarandiba), bem como entre Diamantina e Bandeirinha (povoado de Diamantina):
“Entre Tijuco e Bandeirinha o terreno é árido e arenoso e não apresenta senão campos, compostos de plantas herbáceas. Apesar da extrema secura encontrei em flor cerca de 30 plantas que ainda não possuía. Eram, entre outras, 2 ou 3 belas Melastomatáceas, 2 Ericaceas, o Ionudum Ianatum ASH., várias Polygala, enfim a encantadora Deulexia muscosa A. S. Hil., que se assemelha a um musgo por suas pequenas folhas e seus caules estendidos sobre o chão” (Saint-Hilaire, 2004 [1830], p. 39).
Os campos naturais correlacionam-se aos mosaicos de Campos Limpos, Campos Sujos e Campos Rupestres do bioma Cerrado, além de configurarem-se em ambientes de ecótonos entre Cerrado e Mata Atlântica (Ribeiro & Walter, 2008; IBGE, 2012), como no caso do distrito de Penha de França (fig. 2, fig. 3D).
2.6. Campos artificiais e capoeiras
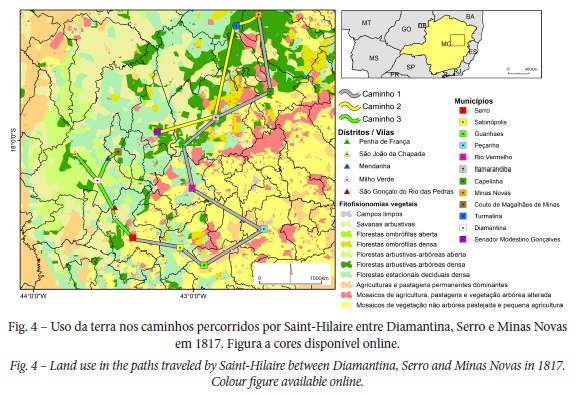
Os campos artificiais e capoeiras são descritos entre Serro, Rio Vermelho e Minas Novas após retirada de matas virgens, catingas e carrasqueiros para o plantio de tabaco, cana-de-açúcar, milho, feijão e algodão. Neste sentido: “Para formar os campos [artificiais] de algodoeiros, só se prepara a terra pela queima das matas que o cobrem. Ao mesmo tempo que as sementes de algodoeiro, costuma-se, também, plantar milho” (Saint-Hilaire, 2000 [1833], p. 235). Desta forma, os campos artificiais constituem-se por plantas pioneiras que se desenvolvem em ambientes degradados em estágios iniciais de sucessão ecológica, e associadas a diversas fitofisionomias savânicas ou florestais (IBGE, 2012). O naturalista cita, por toda a área de estudo, espécies invasoras como samambaias ou fetos (Pteridium caudatum) e capim-gordura (Melinis minutiflora): “Por todos os lados é rodeada por colinas áridas e despidas, absolutamente impróprias para a cultura. Sobre algumas, entretanto, ainda cresce o capim-gordura; em outras encontra-se em companhia de malváceas e o saccharum” (Saint-Hilaire, 2000 [1833], p. 135). Esta invasão biológica é expressiva em toda a Reserva da Biosfera da Serra do Espinhaço (Mucida et al., 2019) como cuja porção setentrional é abarcada por este trabalho (fig. 3E,F e fig. 4).
Saint-Hilaire criticava as derrubadas e queimadas de matas, uma vez que não havia finalidade à madeira explorada. O naturalista defendia, ainda, que os mineiros deveriam “fazer o uso dos arados e dos fertilizantes, e não mais teriam necessidade de destruir suas matas (...)” (Saint-Hilaire, 2000 [1833], p. 92).
De acordo com o IBGE (2012) espécies invasoras como as samambaias (Pteridium sp.) apresentam maior predominância na primeira fase de sucessão natural do sistema da vegetação secundária (tratos antrópicos). Já as fases da sucessão natural: segunda, terceira, quarta e quinta são denominadas respectivamente como: capoeirinha, capoeira rala e capoeira e capoeirão (IBGE, 2012).
IV. Discussão
1. A singularidade ecotonal da região
A singularidade ecotonal existente na região deve ser considerada como resposta aos aspetos ambientais e florísticos. No caso dos aspetos ambientais, a heterogeneidade geológica, geomorfológica, edáfica e climática criou especificidades abióticas que refletem nas bióticas, especialmente na vegetação. Tais características, somadas aos contatos com os diferentes domínios vegetacionais, acarretam em alta diversidade florística, que não explica-se apenas pelos gradientes verticais (Ribeiro & Freitas, 2010). As variações ambientais associadas ao componente ecotonal dos biomas Cerrado, Mata Atlântica e Caatinga, formaram os mosaicos vegetacionais existentes na região (Coutinho, 2006). Este complexo vegetacional é traduzido em vários trechos da literatura de viagem de Saint-Hilaire para a região, que destaca a biodiversidade vinculada à geodiversidade. De facto, áreas transicionais já foram identificadas como especialmente ricas em biodiversidade e considerado prioritário para ações de conservação (Reis, Bueno, Rocha, Santos, & Gorgens, 2019).
O período do ano em que Saint-Hilaire percorreu os três caminhos compreendeu duas estações climáticas bem definidas (verão chuvoso e inverno seco). Assim, não se pode descartar que variáveis ambientais podem ter influenciado coletas e descrições da vegetação ao longo do percurso do naturalista em 1817. Espécies de vegetação aberta ou bordas de florestas, geralmente associam-se a temperaturas mais elevadas, logo, variações climáticas ao longo do tempo implicam na ocorrência de grupos de espécies (Huey et al., 2009; Closel & Kohlsdorf, 2012). Neste sentido, dependendo do período do ano em que o naturalista tenha percorrido os trajetos da região, grupos taxonómicos podem ter sido coletados ou podem ter sido negligenciados, por não apresentarem estruturas (flores e/ou frutos) que possibilitassem sua correta identificação.
2. Análise dos domínios vegetacionais e uso do solo
Saint-Hilaire apropriou-se de terminologias em suas literaturas de viagem comuns nos processos de classificação e mapeamento da vegetação (Oliveira-Filho & Fluminhan-Filho, 1999). A expressão mata virgem ainda é amplamente utilizada no senso comum, às vezes como sinónimo de floresta primária (Carvalho, 2010).
A compreensão da estrutura e a dinâmica populacional das espécies é útil para predizer padrões de ocorrência e abundância, fornecendo subsídios para estratégias de conservação e recuperação em ambientes perturbados. Dentre as diversas tipologias observadas pelo naturalista, destacam-se as Florestas Estacionais, que embora sejam representadas por fragmentos, são fundamentais para entender a abrangência dessa vegetação na região, tanto em nível de comunidade quanto em espécies individuais (Oliveira, Medeiros, Moser, Pinheiro, & Olsen, 2011; Trautenmüllee et al., 2019).
As áreas de Florestas Estacionais vêm sofrendo desde o século XIX com o processo de degradação, principalmente com as queimadas, até os dias atuais (Mucida et al., 2019). Somado a isso, a região apresenta elevada fragilidade ambiental potencial (França, 2018), tendo indicativos de maiores probabilidades de ocorrência de degradação.
Quanto às catingas e carrasqueiros, vinculados ao bioma Caatinga, pode-se afirmar que fatores climáticos, compartimentação topográfica, edáficos e antrópicos são responsáveis por suas diferenças fisionómicas e florística (Andrade-Lima, 1981; Alves, Jackson, Araújo, & Santos, 2009). A passagem do naturalista pelo domínio das Caatingas da região ocorreu em meses mais secos. Ele narra, frequentemente, a deciduidade da vegetação de porte arbóreo como uma característica diferencial entre as formações florestais da região (matas virgens e catingas) o que ressalta a relevância desta condição ecológica. Este fator foi balizador na equivalência das catingas às Florestas Estacionais Deciduais ou Matas Secas.
O naturalista associa domínios vegetacionais ao uso do solo. Ambientes de matas virgens, catingas e carrasqueiros vinculam-se a áreas de cultura, ou a ampliação destas. O surgimento de capoeiras, por exemplo, evidencia a prática do sistema de pousio na agricultura da região (Lopes, 2012). Mucida et al. (2019) argumentam que Saint-Hilaire enfatiza que queimadas, cultivos e mineração contribuíam para a expansão do capim-gordura e samambaias nestas áreas degradadas. De certa forma, o naturalista preocupava-se com a fixação humana na terra, como forma de impulsionar a exploração mais racional dos recursos naturais, mas de maneira a diminuir a pressão antrópica exercida nesses ambientes, devido ao mau uso da ocupação da terra.
A associação dos carrascos à fitofisionomias do Cerrado (Cerrado Sentido Restrito, variando em Cerrado Denso, Cerrado Típico, Cerrado Ralo e Cerrado Rupestre) deu-se pelo porte, densidade e florística descrita. Essa comparação relaciona-se à presença de alumínio e elevada acidez do solo que condicionam árvores baixas, inclinadas, tortuosas (Ribeiro & Walter, 2008). Brandão, Pignal, Romaniuc, Grael, e Fagg (2012) correlacionaram plantas nativas do Cerrado Sentido Restrito em áreas classificadas pelo naturalista como carrascos, devido a presença das famílias botânicas: Verbenaceae, Rubiaceae, Bignoneaceae, Malpighiaceae, Melastomataceae, Myrtaceae e Apocynaceae.
Espécies das famílias Verbenaceae e Rubiaceae são reconhecidas por Saint-Hilaire por seu uso medicinal na forma de chás (Saint-Hilaire, 2014 [1837]) ou como uso potencial em cosméticos, como o Chá-de-Pedestre (Lantana pseudothea), o gervão (Starchytarpheta jamaicensis), poya-do-campo (Richardia scabra) e a malva-do-campo (Kielmeyera spiciosa), muitas delas, revisitadas em estudos como Brandão et al. (2012) e Mügge, Paula-Souza, Melo, e Brandão (2016).
Os ambientes naturais destas espécies vêm a sofrer degradação desde 1970 com a inserção da monocultura de eucalipto e da mineração (Brandão et al., 2012). A ocupação e o uso do Cerrado, geralmente, associam-se à retirada da cobertura vegetal ou possíveis queimadas (Menezes, Mucida, França, Aguilar, & Pereira (2019), interferindo nos diferentes ciclos biogeoquímicos e no equilíbrio dinâmico dos ecossistemas, fundamentais na distribuição da vegetação na paisagem (Martins, Marques, Santos, & Marques, 2015). Neste sentido, infere-se que tais processos antrópicos, há mais de 200 anos delinearam mudanças ecológicas significativas.
Habitats preferenciais no estabelecimento das espécies são marcados por fatores ambientais que interferem positiva ou negativamente sobre a presença das espécies em determinado local (Finger & Oestreich Filho, 2014). Dependendo das condições ecológicas encontradas, essas espécies podem ocasionar um impacte negativo ao ecossistema e tornarem-se invasoras: espécies exóticas em ecossistema natural ou antrópico, que desenvolve altas taxas de crescimento, reprodução e dispersão (Matos & Pivello, 2009).
As espécies encontradas nos campos naturais, especialmente da Serra do Espinhaço, representam uma rica biodiversidade. Estima-se que um quarto das novas espécies descritas no Brasil entre 1997-2002 encontram-se nesse território e seguramente muitas espécies ainda não foram descobertas (Rapini, Ribeiro, Lamberti, & Pirani, 2008). Além disso, as formações campestres do Cerrado (campos sujo, limpo e rupestre) e formações savânicas possuem baixo grau de resiliência (Negreiros, Fernandes, Berbara, Rodarte, & Barbosa, 2011).
Já campos artificiais e as capoeiras correspondem a vegetação em estágio de sucessão ecológica com graus diferenciados de desenvolvimento. Transformações importantes estão a ocorrer no conjunto de espécies, com o aumento populacional e possível imigração de espécies adaptadas a ambientes florestais e provável extinção das não adaptadas. Cabe lembrar que o estado de fragmentação da paisagem na região pode dificultar a chegada de novas espécies, impedindo ou atrasando esse processo (Pinheiro & Durigan, 2009).
Dessa forma, tal discussão torna-se importante para o subsídio da criação de Unidades de Conservação, devido a diversidade encontrada na região, além de parte dos seus caminhos estarem vinculados ao Património Histórico e Cultural (Gonzaga et al., 2018). Ribeiro e Freitas (2010) apontam que a conservação em países com elevada diversidade, como o Brasil e mais especificamente na Serra do Espinhaço, contribui para que não haja perda da flora autóctone de espécies que a ciência desconhece.
V. Considerações finais
A partir das análises da vegetação e descrições botânicas contidas em obras de Saint-Hilaire, em seu trajeto Diamantina-Serro-Minas Novas, foi possível comparar domínios vegetacionais descritos pelo naturalista às fitofisionomias atuais.
Os mapas gerados neste estudo agregaram na geovisualização das características analisadas e subsidiaram a delimitação do modelo dos trajetos realizados por Saint-Hilaire, sendo fundamentais para estudos de análises em escala de paisagens. As limitações cartográficas encontram-se na ausência de dados públicos mais detalhados para a região, bem como sobre organização fitofisionómica, logo, devem ser tomadas como uma aproximação generalista das realidades de campo.
Na região de estudo, as áreas mais elevadas, compreendidas na Serra do Espinhaço Meridional, são moldadas sobre os quartzitos e a descrição florístico-estrutural da vegetação registra fitofisionomias campestres e savânicas do Cerrado. Entre Diamantina e Serro há transição de fitofisionomias do Bioma Cerrado para Mata Atlântica, como Florestas Estacionais Semideciduais. Do Serro a Minas Novas, há transição dos Biomas Mata Atlântica, Cerrado e Caatinga. Dessa forma, o trajeto Saint-Hilaire permite visualizar domínios vegetacionais diversos, que formam verdadeiro mosaico vegetacional de alta relevância de diversidade taxonómica, fitofisionómica e conservacionista, importantes ferramentas para as análises de paisagem voltadas para o campo da geografia, ecologia e fitogeografia local e regional
Esses ambientes, apesar de heterogêneos e únicos em termos de diversidade e endemismo, passam por processos degradatórios vinculados ao extrativismo mineral ou a atividades agropecuárias desde o século XVIII até os dias atuais. Desta forma, parte da vegetação observada e registrada por Saint-Hilaire encontra-se desfigurada, restando resquícios provenientes da sucessão ecológica dos ambientes.
As comparações entre descrições de naturalistas do século XIX com a literatura atual é viável e de aplicabilidade relevante, uma vez que contribuem em levantamentos de informações ecológicas, biológicas e geográficas regionais e nacionais, como evolução temporal, subsidiando estudos e estratégias para a conservação dos recursos naturais dos biomas.
REFERÊNCIAS BIBLIOGRÁFICAS
Alves, A., Jackson, J., Araújo, M. A., & Santos, S. N. (2009). Degradação da caatinga: uma investigação ecogeográfica [Degradation of Caatinga: An Investigation Geographical Ecology]. Caatinga, 22(3), 126-135.
Andrade-Lima, D. A. (1981). The caatinga dominium. Revista Brasileira de Botânica, 4(1), 149-153. [ Links ]
Ávila, G. E. (2001). Geometria e Fundamentos [Geometry and Fundamentals]. Revista do Professor de Matemática, 45.
Brandão, M. G. L., Pignal, M. Romaniuc, S., Grael, C. F. F., & Fagg, W. (2012). Useful Brazilian plants listed in the field books of the French naturalist Auguste de Saint-Hilaire (1779-1853). Journal of Ethnopharmacology, 143(2), 488-500. DOI: 10.1016/j.jep.2012.06.052. [ Links ]
Carvalho, E. B. (2010). No fundo da mata virgem: a complexidade de um elemento mítico no imaginário ocidental sobre a natureza [Deep inside the virgin forest: the complexity of a mythical element in the western imaginary about nature]. Revista Tempo e Argumento, 2(2), 135-153.
Closel, M. B., & Kohlsdorf, T. (2012). Climatic changes and fossoriality: implications for the subterranean herpetofauna. Revista da Biologia, 8, 19-24. DOI: 10.7594/revbio.08.04. [ Links ]
Coutinho, L. M. (2006). O Conceito de Bioma [The Biome Concept]. Acta Botânica brasílica, 20(1), 13-23.
França, L. C. J. (2018). Fragilidade Ambiental Potencial da bacia hidrográfica do rio Jequitinhonha, Minas Gerais, Brasil [Environmental Fragility Potential of the river Jequitinhonha basin, Minas Gerais, Brazil]. (Dissertação de Mestrado). Retrieved from http://acervo.ufvjm.edu.br/jspui/handle/1/1585 [ Links ]
Ferreira Neto, M. C. N. (2003). O olhar analítico de um observador vigilante [The analytical look of a watchful observer]. História Social, 10, 149-179.
Finger, Z., & Oestreich Filho, E. (2014). Efeitos do solo e da altitude sobre a distribuição de espécies arbóreas em remanescentes de Cerrado sensu stricto [Effects of the soil and altitude on the distribution of tree species in remainders of sensu stricto Cerrado]. Advances in Forestry Science, 1(1), 27-33. DOI: 10.34062/afs.v1i1.1226.
Flora do Brasil 2020. (2018). Jardim Botânico do Rio de Janeiro [Rio de Janeiro Botanical Garden] Retrieved from http://floradobrasil.jbrj.gov.br/ [ Links ]
Gonzaga, A. P. D., Piuzana, D., Machado, E. L. M., & Morais, M. S. (2018). (Re)visitando a Reserva da Biosfera da Serra do Espinhaço com Martius e Spix (1818-2018) [(Re)visiting the Espinhaço Range Biosphere Reserve with Martius and Spix (1818-2018)]. Diamantina: UFVJM. [ Links ]
Huey, R. B., Deutsch, C. A., Tewksbury, J. J., Vitt, L. J., Hertz, P. E., Álvarez Pérez, H. J., & Garland, J., T. (2009). Why tropical forest lizards are vulnerable to climate warming. Proceedings of the Royal Society B: Biological Sciences, 276(1664), 1939-1948. DOI: 10.1098/rspb.2008.1957. [ Links ]
Kury, L. (2001). Viajantes-naturalistas no Brasil oitocentista: experiência, relato e imagem [Scientific travellers in eighteenth-century Brazil: their experience, reports and graphics representations]. História, Ciências, Saúde-Manguinhos, 8(Suppl.), 863-880. DOI: 10.1590/S0104-59702001000500004.
Instituto Brasileiro De Geografia e Estatística. (IBGE). (2018). Bases cartográficas e malha digital [Cartographic bases and digital mesh]. Retrieved from https://mapas.ibge.gov.br/bases-e-referenciais/bases-cartograficas/malhas-digitais [ Links ]
Instituto Brasileiro De Geografia e Estatística. (IBGE). (2012). Manual Técnico da Vegetação Brasileira [Technical Manual of the Brazilian vegetation]. Rio de Janeiro: IBGE.
Instituto Prístino. (2018). Atlas Digital Geoambiental [Geoenvironmental Digital Atlas]. Retrieved from https://institutopristino.org.br/atlas/ [ Links ]
Lopes, F. A., Milagres, A. R., Piuzana, D., & Morais, M. S. (2011). Viajantes e Naturalistas do Século XIX: A Reconstrução do Antigo Distrito Diamantino na Literatura de Viagem [Travelers and Naturalists of the 19th Century: The Reconstruction of the Ancient Diamantine District in Travel Literature]. Caderno de Geografia, 21(36), 66-84.
Lopes, Q. I. (2012). Entre capoeiras e matas virgens: a prática agrícola extensiva em Minas Gerais Setecentista [Between capoeiras and virgin forests: the extensive agricultural practice in eighteenth century Minas Gerais]. Revista de História Econômica & Economia Regional Aplicada, 7(13).
Martins, K. G., Marques, M. C. M., Santos, E., & Marques, R. (2015). Effects of soil conditions on the diversity of tropical forests across a successional gradient. Forest Ecology and Management, 349, 4-11. DOI: 10.1016/j.foreco.2015.04.018. [ Links ]
Matos, D. M. S., & Pivello, V. R. (2009). O impacto das plantas invasoras nos recursos naturais de ambientes terrestres: alguns casos brasileiros [The impact of invasive plants on natural resource environmental resources - some brazilian cases]. Ciência e Cultura, 61(1), 27-30.
Menezes, E. S., Mucida, D. P., França, L. C. J., Aguilar, M. V. M., & Pereira, I. M. (2019). Análise temporal de focos de calor na reserva da biosfera da Serra do Espinhaço [Temporal Analysis of heat foci at Espinhaço Range biosphere reserve]. Nativa, 7(3), 256-261. DOI: 10.31413/nativa.v7i3.6877.
Milan, E., & Moro. R. S. (2016). O conceito biogeográfico de ecótono [The biogeographic concept of ecotone]. Revista Terr@Plural, 10(1), 75-88.
Ministério Do Meio Ambiente. (MMA). (2002). Mapas de Cobertura Vegetal dos Biomas Brasileiros [Maps of Vegetable Cover of the Brazilian Biomes]. Retrieved from http://mapas.mma.gov.br/mapas/aplic/probio/datadownload.htm
Mucida, D. P., Gontijo, B., Morais, M., & Fagundes, M. (2019). A degradação ambiental em narrativas de naturalistas do século XIX para a reserva da Biosfera da Serra do Espinhaço [Environmental degradation in narratives of naturalists of the 19th century for the Espinhaço Range Biosphere Reserve]. Caderno de Geografia, 29(57), 465-495. DOI: 10.5752/P.2318-2962.2019v29n57p465-495.
Mügge, F. L. B., Paula-Souza, J., Melo, J. C., & Brandão, M. G. L. (2016). Native plants species with economic value from Minas Gerais and Goiás: a discussion on the currentness of the data recovered by the French naturalist Auguste de Saint-Hilaire. Horticultura Brasileira, 34, 455-462. DOI: 10.1590/s0102-053620160402. [ Links ]
Negreiros, D., Fernandes, G. W., Berbara, R. L. L., Rodarte, L. H. O., & Barbosa, N. P. U. (2011). Caracterização físico-química de solos quartzíticos degradados e áreas adjacentes de campo rupestre na Serra do Cipó, MG, Brasil [Physical-chemical characterization of degraded quartzite soils and adjacent rock field in Serra do Cipó, MG, Brazil]. Neotropical Biology and Conservation, 6(3), 156-161. DOI: 10.4013/nbc.2011.63.02.
Noce, C. M., Pedrosa-Soares, A., Silva, L., & Alkmim, F. (2007). O embasamento arqueano e paleoproterozoico do Orógeno Araçuaí [The Archaean and Paleoproterozoic basement of the Araçuaí Orogen]. Revista Geonomos, 15(1), 17-23. DOI: 10.18285/geonomos.v15i1.104.
Oliveira, W. L., Medeiros, M. B., Moser, P., Pinheiro, R., & Olsen, L. B. (2011). Regeneração e estrutura populacional de jatobá-da-mata (Hymenaea courbaril L.), em dois fragmentos com diferentes graus de perturbação antrópica [Regeneration and population structure of jatobá-da-mata (Hymenaea courbaril L.) in two fragments with distinct degrees of anthropic disturbance]. Acta Botanica Brasilica, 25(4), 876-884. DOI: 10.1590/S0102-33062011000400014.
Oliveira-Filho, A. T., & Fluminhan-Filho, M. (1999). Ecologia da vegetação do Parque Florestal Quedas do Rio bonito. [Vegetation ecology of the Parque Florestal Quedas do Rio Bonito, Brazil]. CERNE, 5(2), 51-64.
Pinheiro, E. S., & Durigan, G. (2009). Dinâmica espaço-temporal (1962-2006) das fitofisionomias em unidade de conservação do Cerrado no sudeste do Brasil [Spatial-temporal dynamics (1962-2006) of the phytophysiognomies in the Cerrado conservation unit in southeastern Brazil]. Brazilian Journal of Botany, 32(3), 441-454. DOI: 10.1590/S0100-84042009000300005.
Rapini, A., Ribeiro, P. L., Lambert, S., & Pirani, J. R. (2008). A flora dos campos rupestres da Cadeia do Espinhaço [The flora of the rupestrian fields of Cadeia do Espinhaço]. Megadiversidade, 4(1-2), 16-24.
Reis, C. R., Bueno, M. L., Rocha, L. F., Santos, L. G., & Gorgens. E. B. (2019). Climate Change Influencing the Potential Distribution of a Brazilian Savanna Indicator Species. FLORAM, 26(3), e20180085. DOI: 10.1590/2179-8087.008518. [ Links ]
Ribeiro, K. T., Freitas, L. (2010). Impactos potenciais das alterações no Código Florestal sobre a vegetação de campos rupestres e campos de altitude [Potencial impacts of changes in the Forest Code on the vegetation of rock fields and altitude fields]. Biota Neotropica, 10(4), 239-246.
Ribeiro, F., & Walter, B. M. T. (2008). As principais fitofisionomias do bioma Cerrado [The main phytophysiognomies of the Cerrado biome]. In S. M. Sano, S. P. Almeida, & J. F. Ribeiro (Eds.), Cerrado: ecologia e flora [Cerrado: ecology and flora] (pp.151-212). Brasília: EMBRAPA. [ Links ]
Saadi, A. (1995). A Geomorfologia da Serra do Espinhaço em Minas Gerais e de suas Margens [The Geomorphology of the Espinhaço Range in Minas Gerais and its Margin]. Revista Geonomos, 3(1), 41-63. DOI: 10.18285/geonomos.v3i1.215.
Saint-Hilaire, A. (2014 [1828]). Plantas Usuais dos Brasileiros [Usual Plants of Brazilians]. Belo Horizonte: Fino Traço. [ Links ]
Saint-Hilaire, A. (2011 [1837]. Quadro geográfico da vegetação primitiva na província de Minas Gerais [Geographical chart of the primitive vegetation in the province of Minas Gerais]. Belo Horizonte: Fino Traço.
Saint-Hilaire, A. (2004 [1830]). Viagem pelo distrito dos diamantes e litoral do Brasil [Travel through the Diamond and Coastal District of Brazil]. Belo Horizonte: Itatiaia.
Saint-Hilaire, A. (2000 [1833]). Viagem pelas províncias do Rio de Janeiro e Minas Gerais [Travel through the provinces of Rio de Janeiro and Minas Gerais]. Belo Horizonte: Itatiaia, São Paulo: Edusp.
Walter, B. M. T. (2006). Fitofisionomias do bioma Cerrado: síntese terminológica e relações florísticas [Phytophysiognomies of the Cerrado biome: terminological synthesis and floristic relations]. (Tese de Doutorado). Retrieved from http://repositorio.unb.br/handle/10482/3086 [ Links ]
Agradecimentos
Este estudo foi parcialmente financiado pela Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) - Código Financeiro 001. Os autores agradecem aos revisores anônimos pela leitura cuidadosa de nosso manuscrito e por significativos comentários e sugestões.
Recebido: maio 2019. Aceite: novembro 2019.

