








Servicios Personalizados
Revista
Articulo
 Portugués (pdf)
Portugués (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Ciências Agrárias
versión impresa ISSN 0871-018X
Rev. de Ciências Agrárias vol.36 no.1 Lisboa ene. 2013
Condicionamento das sementes e sombreamento na emergência e no crescimento de plantas de Bixa orellana L.
Seed priming and shading on emergency and initial growth of Bixa orellana L. seedlings
Camila Kissmann1; Silvana de Paula Quintão Scalon2 e Tiara Kesli Conticelli Teodósio3
1 Universidade Federal da Grande Dourados – Faculdade de Ciências Agrárias (FCA), Rodovia Dourados- Itahum, Km12, Dourados-MS. E-mail:camilakissmann@gmail.com
2 Universidade Federal da Grande Dourados, Rodovia Dourados- Itahum, Km12, CEP: 79804970, Dourados-MS. E-mail:silvanascalon@ufgd.edu.br
3 Universidade Federal da Grande Dourados – FCA, Rodovia Dourados- Itahum, Km12, Dourados-MS. E-mail:tiarakesli@yahoo.com.br
RESUMO
Foram realizados dois experimentos para avaliar os efeitos da temperatura e do período de condicionamento na germinação de sementes e o crescimento inicial de mudas, e o efeito do sombreamento sobre o crescimento inicial das mudas de urucum. No primeiro, as sementes foram pré-embebidas em soluções contendo polietilenoglicol (PEG) e nitrato de potássio consoante as seguintes modalidades: 1) PEG 6000 (-1,0 MPa); 2) PEG 6000 (-1,0 MPa) + KNO3 (-1,0 MPa); 3) KNO3 (-1,0 MPa); 4) água e 5) água deionizada, e incubadas em câmara de germinação tipo BOD em temperatura de 10 e 20°C durante 0 (controle), 6, 12 e 24 h. No segundo experimento as sementes foram pré-condicionadas em solução de PEG 6000 + KNO3 e em água deionizada, e incubadas a 10°C durante 24 h, além da testemunha e, semeadas em sacos de polietileno e acondicionadas em casas de vegetação sob sombreamento de 0, 50 ou 70%. O condicionamento osmótico a 10°C e a solução combinada de PEG 6000 + KNO3 durante 24 horas proporcionaram maior porcentagem de germinação (35,87%). O sombreamento 50% foi o mais favorável a emergência das plântulas e ao crescimento das mudas.
Palavras-chave: Bixaceae, germinação, KNO3, osmocondicionamento, polietilenoglicol.
ABSTRACT
Two experiments were carried out to evaluate the effect of period and temperature of conditioning in the germination of seeds and seedlings growth, and the effect of shading on the seedlings initial growth. In the first one, the seeds was conditioned in the following solutions: 1) PEG 6000 (- 1,0 MPa); 2) PEG 6000 (- 1,0 MPa) + KNO3 (- 1,0 MPa); 3) KNO3 (- 1,0 MPa); 4) water and 5) deionized water, and placed to germinate in BOD regulated to provide temperatures of 10 and 20°C during 0 (control), 6, 12 and 24 h. In the second experiment, the seeds were conditioned in solution of PEG 6000 + KNO3 and in deionized water and placed in BOD regulated to 10°C during 24 h, beyond the control. After this, they were sown in polyethylene bags and placed in greenhouses under 0, 50 or 70% shading. The conditioning at 10°C and in the mix solution of PEG 6000 + KNO3, during 24 h provided the bigger percentage of germination (35.87%). Artificial shade of 50% was the most favorable to emergency and growth of annatto seedlings.
Keywords: Bixaceae, germination, KNO3, osmoconditioning, poliethyleneglicol.
Introdução
O urucum (Bixa orellana L.) é considerado uma das principais fontes de corantes naturais utilizados mundialmente. Tal propriedade tintorial deve-se a um carotenóide, conhecido como bixina, encontrado no arilo dessas sementes (Corlett et al., 2007). O emprego de corantes naturais em substituição aos artificiais, por diversos setores da indústria, tem gerado um crescente interesse pelos produtos provenientes do urucum. Além da aplicabilidade na indústria alimentícia (embutidos, laticínios, bebidas, entre outros) este corante é utilizado por vários outros segmentos não alimentícios, como por exemplo, nas indústrias cosméticas, farmacêuticas e têxteis (Nakano, 1998). Além dessas aplicações, o urucuzeiro pode ainda ser explorado como planta ornamental e para a recuperação de áreas degradadas, devido ao rápido crescimento (Lorenzi, 2000). Entretanto, a propagação sexuada desta espécie é dificultada pela baixa porcentagem de germinação das sementes, devido à dormência imposta pela impermeabilidade do tegumento (Amaral et al., 1995; Custódio et al., 2002; Lopes et al., 2008).
Muitas técnicas têm sido propostas para a realização de tratamentos de pré-semeadura, com o objetivo de reduzir o tempo necessário entre a semeadura e a emergência das plântulas, bem como o aumento da resistência das sementes aos diferentes tipos de estresse ambiental (Sune et al., 2002). Dentre estas técnicas destaca-se o osmocondicionamento ou priming, inicialmente proposta por Heydecker et al. (1975).
Para culturas agrícolas os efeitos do osmocondicionamento consistem na melhoria do processo germinativo e no vigor das sementes (Nunes et al., 2004; Marcos Filho, 2005), enquanto que para diversas hortaliças o osmocondicionamento melhorou a velocidade e uniformidade de germinação de suas sementes (Sune et al., 2002). Porém, para espécies florestais, devido à grande diversidade da flora brasileira e menor interesse econômico, ainda há poucas informações a respeito do condicionamento osmótico das sementes (Lars, 2000).
Dentre os estudos com espécies florestais, destacam-se os efeitos benéficos do condicionamento osmótico nos trabalhos de Borges et al. (1994) e Córdoba et al. (1995) para as espécies Miconia candolleana Trian (quaresminha) e Esenbeckia leiocarpa England (gua- rantã), respectivamente. Porém, sementes de Peltophorum dubium Spreng. Taub (canafístula) não mostraram melhoria no vigor e na viabilidade quando condicionadas em água ou em PEG (-1,0MPa) a 10 ou a 27°C (Wanli et al., 2001). Também para Pterogyne nitens Tul. (amendoim do campo) não foi observado efeito significativo do osmocondicionamento das sementes (Biruel et al., 2007). Dessa forma, muitos aspectos da utilização racional desta técnica e de seus efeitos ainda necessitam ser elucidados, como por exemplo, a determinação do agente osmótico, o tempo de exposição das sementes e o potencial hídrico da solução osmótica para o osmocondicionamento de cada espécie.
O processo de emergência e o estádio de plântula representam um período particularmente sensível no desenvolvimento do vegetal, sendo que, dentre os diversos componentes do ambiente, a luz é primordial para o crescimento das plantas. Variações na presença/ausência ou nos níveis de luz influenciam o desenvolvimento do vegetal, uma vez que afetam a abertura estomática e a síntese de clorofila a, e consequentemente, a produção de matéria orgânica (Kozlowski et al., 1991; Atroch et al., 2001). A capacidade de uma determinada espécie em adaptar-se a condições de sol ou sombra pode ser avaliada pelo crescimento inicial das plantas em diferentes condições de radiação luminosa (Valladares et al., 2000).
Estudos sobre a germinação do urucum são neces- sários para otimizar a propagação sexuada e garantir a disponibilidade de mudas, seja com finalidade econômica ou conservacionista. O objetivo deste trabalho foi avaliar o efeito da temperatura e do período de condicionamento na germinação de sementes e emergência de plântulas de urucum, e o efeito de diferentes níveis de sombreamento no crescimento inicial das mudas de urucum.
Material e Métodos
As sementes utilizadas foram obtidas de frutos em estádio inicial de deiscência, colhidos de 10 matrizes no Horto de Plantas Medicinais da Universidade Federal da Grande Dourados (UFGD) em junho de 2007. As sementes foram removidas manualmente dos frutos e armazenadas em sacos de papel Kraft® em câmara fria e seca (17°C e 69%UR) até o início dos dois experimentos.
Experimento 1
Foi realizado de outubro a dezembro de 2007. As sementes foram distribuídas em caixas gerbox forradas com duas folhas de papel filtro umedecidas com 15 mL das respectivas soluções-teste: 1) PEG 6000 (-1,0 MPa); 2) PEG 6000 (-1,0 MPa) + KNO3 (-1,0 MPa); 3) KNO3 (-1,0 MPa); 4) Água e 5) Água deionizada (teste- munha). O cálculo para a concentração do PEG foi ob- tido de acordo Villela et al. (1991) e a concentração de KNO3 de acordo com a equação de Vant Hoff, citada por Hillel (1971); para a mistura de KNO3 e PEG 6000 foi desconsiderada a interação entre os dois produtos.
Os gerbox foram envolvidos com filme plástico para evitar a evaporação e colocados em câmaras de germinação (BOD) nas temperaturas de 10 e 20°C durante 0 (controle), 6, 12 e 24 h. Após cada tempo de embebição, as sementes foram retiradas da BOD e permaneceram em condição de ambiente de laboratório até atingirem o peso inicial de antes do condicionamento. Em seguida foram semeadas em caixas gerbox sobre duas folhas de papel filtro e mantidas em BOD em temperatura de 20-30°C em regime foto- periódico de 8 h escuro e 16 h de luz.
Os parâmetros avaliados foram: porcentagem de germinação (%G) e índice de velocidade de germinação (IVG) (conforme a fórmula proposta por Maguire (1962): IVG = N1/D1 + N2/D2 + Nn/Dn onde N1, N2 e Nn= número de sementes germinadas na primeira, segunda, até a última contagem; e D1, D2... Dn= número de dias transcorridos da semeadura até a primeira, segunda e última avaliação).
O delineamento estatístico utilizado foi o inteiramente casualizado, em esquema fatorial 5 x 2 x 4 (tratamentos de pré-condicionamento, temperaturas de incubação e tempos de incubação) em quatro repetições de 20 sementes.
Experimento 2
O segundo experimento estendeu-se de fevereiro a julho de 2008. As sementes foram embebidas em soluções de PEG + KNO3 (experimento 1) e em água deionizada e incubadas em câmaras BOD a 10°C durante 24 horas, seguindo-se o mesmo procedimento do primeiro experimento. Em seguida, as sementes condicionadas e não condicionadas (testemunha) foram semeadas em sacos de polietileno com capacidade de 1,5 cm3 preenchidos com terra + areia + cama de frango semidecomposta na proporção de 1:1:1 (v:v) e acondicionadas em estufa sob 50 e 70% de sombreamento e em condição de pleno sol (0% de sombreamento). As reduções dos dois níveis de radiação fotossinteticamente ativa (RFA) foram obtidas por meio do uso de telas pretas de polietileno, conhecidas por Sombrite®.
Neste experimento foram avaliados a porcentagem de emergência (%E) e o índice de velocidade de emergência (IVE) (conforme Maguire, 1962). O experimento foi realizado em delineamento inteiramente casualizado em esquema fatorial 3 x 3 (tratamentos de pré-condicionamento e níveis de sombreamento), em quatro repetições de 20 sementes.
Três repetições de 15 plantas foram selecionadas, aos 50 dias após a semeadura, para avaliações quinzenais quanto à altura, diâmetro do colo e teor de clorofila. Em cada avaliação, três plantas de cada repetição foram tomadas para a determinação do comprimento da maior raiz, área foliar e massa seca de raiz (MSR) e de parte aérea (MSPA) das mudas.
O teor de clorofila foi avaliado com auxílio de clorofilômetro SPAD sendo feita a média de três folhas por planta. A área foliar foi determinada por meio de um medidor de área foliar LICOR modelo 3000.
O peso seco foi obtido pela pesagem do material seco em estufa com circulação de ar a 55oC até peso constante. O delineamento experimental utilizado foi o inteiramente casualizado em esquema fatorial 3 x 3 x 5 (tratamentos de condicionamento, níveis de sombreamento e idades da muda), em 3 repetições de 10 plantas.
Procedimento estatístico
Os valores de germinação, obtidos em porcentagem, foram transformados em arco seno (Snedecor, 1962).
Os dados de ambos os experimentos foram submetidos à análise de variância e havendo significância, as médias foram comparadas pelo teste de Tukey e análise de regressão, a 5% de probabilidade.
Resultados e Discussão
Experimento 1
De acordo com a análise estatística, houve interação significativa (P<0,05) entre as temperaturas e os tempos de embebição, e entre os tratamentos e os tempos de embebição, mas não foi observada interação significativa (P>0,05) para as temperaturas e os tratamentos de embebição.
A temperatura de embebição de 10°C resultou em maior porcentagem de germinação das sementes em comparação a de 20°C (Figura 1A). A temperatura de 10°C também foi melhor que o condicionamento a 20°C para sementes de Peltophorum dubium (Perez & Negreiros, 2001) e Pterogyne nitens (Tonin et al., 2005).
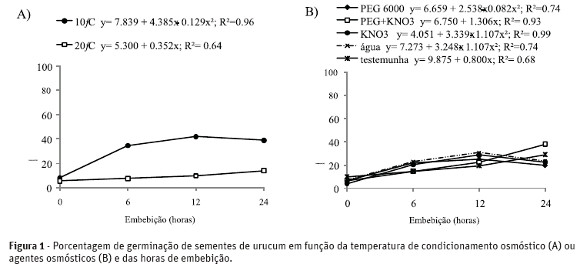
Quando o condicionamento das sementes é realizado em baixas temperaturas, a embebição é lenta e há tempo suficiente para que as membranas celulares, que ao se desidratarem passam de estado fluído para estado de gel, voltem ao estado cristalino líquido, sem ocorrer danos celulares e lixiviação de solutos (Castro & Hilhorst, 2004).
O aumento da germinação das sementes foi proporcional ao aumento das horas de embebição até um período de 12 horas para os condicionamentos em soluções isoladas de PEG 6000 e KNO3 e condicionamento em água, e até 24 h para sementes condicionadas em solução mista de PEG 600 + KNO3 e aquelas não condicionadas (testemunhas). O valor máximo de germinação (35,9%) foi observado para sementes condicionadas em solução mista de PEG 6000 + KNO3, por 24 h (Figura 1B). Todavia, apesar do tratamento de osmocondicionamento, a germinação foi inferior a 50%. A baixa germinação das sementes de urucum é atribuída, em parte, à dormência imposta pelo tegumento conforme relatado nos estudos de Amaral et al. (1995), Custódio et al. (2002) e Lima et al.(2007). Embora tratamentos de escarificação mecânica aumentam a germinação das sementes de urucum (Custódio et al., 2002; Lima et al., 2007), a porcentagem de germinação obtida ainda não é satisfatória para a produção de mudas desta espécie por reprodução sexuada.
Nas condições em que o presente experimento foi realizado, o condicionamento das sementes em água ou em soluções osmóticas não foi suficiente para melhorar o desempenho germinativo destas, possivelmente devido à impermeabilidade do tegumento das sementes do urucum. Sune et al. (2002) observaram que o condicionamento osmótico com aquecimento da solução de polietilenoglicol (com temperatura inicial de 70°C, posteriormente resfriada a 20°C) melhorou o desempenho germinativo de sementes da leguminosa Adesmia latifolia e foi eficiente em superar a dormência tegumentar destas sementes, dispensando a utilização de métodos de superação de dormência normalmente aplicados em leguminosas.
Logo, estudos combinados de superação de dormência e condicionamento das sementes devem ser realizados na tentativa de melhorar a reprodução sexuada do urucum. Além disso, deve-se considerar que a obtenção de resultados satisfatórios para a técnica de condicionamento osmótico depende, dentre outros fatores, da definição de um ajuste osmótico adequado. O efeito do condicionamento depende de fatores externos, tais como a temperatura de embebição, a concentração da solução e a duração do tratamento, e de fatores internos (inerentes da própria semente) (Bradford, 1986) e, a melhor combinação dos fatores envolvidos no tratamento e sua otimização deve ser feita por tentativas para cada lote de sementes (Santos et al., 2008).
Em relação ao índice de velocidade de germinação não houve interação significativa entre os tratamentos, temperaturas e tempos de incubação (P>0,05). A análise dos fatores isolados mostra os maiores valores de IVG para o tratamento de PEG + KNO3 (0,0312), a temperatura de 10°C (0,0125) e o tempo de incubação de 24 horas (0,0248) (dados não mostrados).
Experimento 2
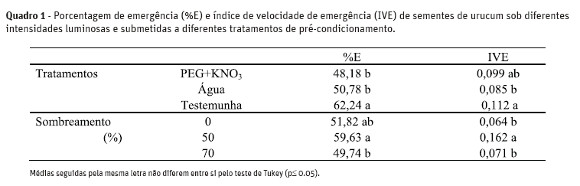
No segundo experimento, o maior valor de emergência foi observado para as sementes que não foram submetidas ao condicionamento (62,2%). Sementes não condicionadas também apresentaram maior valor de IVE, o qual não diferiu significativamente do condicionamento realizado com PEG+KNO3. Mudas mantidas sob 50% de sombreamento apresentaram os maiores valores de %E e IVE (Quadro 1).
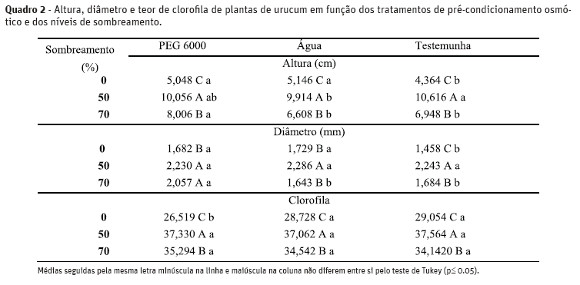
O crescimento das mudas também foi favorecido quando as plântulas foram mantidas sob 50% de sombreamento, exibindo maiores valores de altura, diâmetro e teor de clorofila, enquanto que mudas crescidas em pleno sol apresentaram os menores valores para essas três características (Quadro 2 e Figura 2A, 2B e 2C).

O resultado observado para a altura das mudas corrobora com outros observados na literatura para outras espécies florestais como Croton urucurana Baill (Alvarenga et al., 2003), Bombacopsis glabra (Pasq.) A.
Rob (Scalon et al., 2003), Senna macranthera (Collad.) Irwin et Barn. (Almeida et al., 2005) e Theobroma grandiflorum (Willd. ex Spreng.) Schum. (Silva et al., 2007).
Pesquisas desenvolvidas sobre o crescimento de mudas arbóreas em função da luminosidade têm apontado respostas diferenciadas entre as espécies, principalmente considerando seu estágio sucessional. A capacidade de crescer rapidamente quando moderadamente sombreadas é um mecanismo importante de adaptação da espécie, constituindo uma estratégia de fuga à baixa e à alta intensidade luminosa. A adaptação às baixas luminosidades é uma característica genética, a qual faz com que as folhas tenham estrutura anatômica e propriedades fisiológicas que as capacitem ao uso efetivo da radiação solar disponível (Larcher, 2000).
Foi observada interação significativa entre os tratamentos de embebição e os níveis de sombreamento, entretanto, não houve um padrão de crescimento das mudas. Sob 70% de sombreamento, os maiores valores de altura e diâmetro foram observados para mudas provenientes de sementes tratadas com solução de PEG + KNO3. Sob 50% de sombreamento, a altura das mudas provenientes de sementes condicionadas em solução de PEG + KNO3 não diferiu daquelas provenientes de sementes não condicionadas, enquanto que o diâmetro não variou entre os tratamentos de embebição. Já sob pleno sol, mudas provenientes de sementes não condicionadas apresentaram valores significativamente inferiores aos das mudas provenientes de sementes condicionadas para as variáveis altura e diâmetro (Quadro 2).
Plantas sombreadas apresentaram teores mais elevados de clorofila em relação àquelas cultivadas em pleno sol (Figura 2C). O aumento do conteúdo da clorofila total com a redução da intensidade luminosa tem sido relatado na literatura para outras espécies, como, por exemplo, para Bombacopsis glabra (Scalon et al., 2003), Croton urucurana (Alvarenga et al., 2003), Maclura tinctoria, Senna macranthera e Hymenaea courbaril L. var. stilbocarpa (Almeida et al. 2004).
Não houve interação entre a idade das mudas e os tratamentos pré-germinativos sobre a altura, o diâmetro do colo, teor de clorofila e crescimento da raiz.
Analisando-se os fatores isolados observou-se que independentemente da idade da muda o condicionamento das sementes com PEG 6000 proporcionou os maiores valores de altura (7,70 cm) e diâmetro (1,99 mm) das mudas, enquanto que para o teor de clorofila e comprimento de raiz não houve variação estatística entre os tratamentos estudados, com média de 33,36 µg cm-² e 15,39 cm, respectivamente (dados não mostrados).
Os maiores valores de comprimento de raiz ocorreram em plantas cultivadas sob 50% de sombreamento e pleno sol em relação àquelas cultivadas sob 70% de sombreamento, independentemente do tratamento de condicionamento (Figura 2D). Resultados semelhantes foram obtidos por Martinazzo et al. (2007) para Eugenia uniflora Linn. O aumento da biomassa radicial proporciona um melhor desempenho das mudas no campo, provendo sustentação e maior área de absorção de água e nutrientes (Martinazzo et al., 2007).
A expansão da folha em condição de baixa luminosidade é uma resposta frequentemente relatada e indica uma maneira da planta compensar ou aproveitar melhor à baixa luminosidade (Alvarenga et al., 2003, Silva et al., 2007). As plantas sob baixa luminosidade tendem a expandir o limbo foliar, aumentando a área de captação de luz (Larcher, 2000).
As maiores médias de massa seca de raiz, aos 110 dias de idade das mudas foram observadas para plantas crescidas sob 50% de sombreamento, o que não ocorreu em detrimento do crescimento de parte aérea, pois os maiores valores médios de altura e massa seca de parte aérea também foram observados para esta condição de sombreamento (Figura 4A e 4C). A resposta de acúmulo de massa seca é variável, devido cada espécie possuir características genéticas diferentes e, consequentemente adaptações fisiológicas diferentes. Plantas de Theobroma grandiflorum, por exemplo, apresentaram maior produção de massa seca quando cultivadas sob 50% de sombreamento (Silva et al., 2007), enquanto plantas de Eugenia uniflora apresentaram maior produção de matéria seca quando acondicionadas sob sol pleno (Martinazzo et al., 2007).
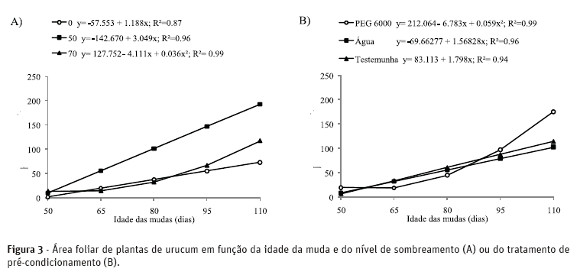

O condicionamento das sementes com PEG resultou em plântulas com maior área foliar, massa seca de raiz e de parte aérea (Figura 3B, 4B e 4D). De modo geral, o condicionamento proporcionou um melhor desenvolvimento das plantas e um maior acúmulo de biomassa, o que pode ser explicado porque durante o condicionamento osmótico ocorrem incrementos no teor de proteínas solúveis e de enzimas específicas, o que proporciona maior concentração de solutos, resultando em crescimento mais rápido e, por conseqüência, maior acúmulo de biomassa (Smith & Cobb, 1991).
Conclusões
As condições mais eficientes para o condicionamento de sementes foi o uso de solução mista de PEG + KNO3 sob 10°C após 24 horas de embebição.
O sombreamento de 50% foi o mais favorável a emergência das plântulas e crescimento da muda e, aos 110 dias, os maiores valores de biomassa foram observados para plântulas originárias de sementes condicionadas.
Referências Bibliográficas
Almeida, L.P.; Alvarenga, A.A.; Castro, E.M.; Zanela, S.M. & Vieira, C.V. (2004) - Crescimento inicial de plantas de Cryptocaria aschersoniana Mez. subme- tidas a níveis de radiação solar. Ciência Rural, 34, 1: 83-88. [ Links ]
Almeida, S.M.Z.; Soares, A.M.; Castro, E.M.; Vieira, C.V. & Gajego, E.B. (2005) - Alterações morfoló- gicas e alocação de biomassa em plantas jovens de espécies florestais sob diferentes condições de sombreamento. Ciência Rural, 35, 1: 62-68. [ Links ]
Alvarenga, A.A.; Castro, E.M.; Lima, E.C. & Magalhães, M.M. (2003) - Effects of different light levels on the initial growth and photosynthesis of Croton urucurana Baill. in southeastern Brazil. Revista Árvore, 27, 1: 53-57. [ Links ]
Amaral, L.I.V.; Pereira, M.F.A. & Cortelazzo, A.L. (1995) - Quebra da dormência em sementes de Bixa orellana. Revista Brasileira de Fisiologia Vegetal, 7, 2: 151-157. [ Links ]
Atroch, E.M.A.C.; Soares, A.M.; Alvarenga, A.A. & Castro, E.M. (2001) - Crescimento, teor de clorofilas, distribuição de biomassa e características anatômicas de plantas jovens de Bauhinia forficata Link. submetidas a diferentes condições de sombreamento. Ciência e Agrotecnologia, 25, 4: 853- 862. [ Links ]
Biruel, R.P.; Borba Filho, A.B.; Araújo, E.C.E.; Fracassaro, F.O. & Perez, S.C.J.G.A. (2007) - Efeitos do condicionamento seguido ou não de secagem em sementes de Pterogyne nitens Tul. sob estresse. Ciência Florestal, 17, 2: 119-128. [ Links ]
Bradford, K.J. (1986) - Manipulation of seed water relations via osmotic priming germination under stress conditions. HortScience, 21, 5: 1105-1112. [ Links ]
Borges, E.E.L.; Silva, L.F. & Borges, R.C.G. (1994) - Avaliação do osmocondicionamento na germinação de sementes de quaresminha (Miconia candolleana Trian.). Revista Brasileira de Sementes, 16, 1: 90-94. [ Links ]
Castro, R.D. & Hilhorst, H.W.M. (2004) - Embebição e reativação do metabolismo. In: Ferreira, A. G. & Borghetti, F. Germinação: do básico do aplicado. Artmed, Porto Alegre, pp.149-162. [ Links ]
Córdoba, G.A.T.; Borges, E.E.L.; Borges, R.C.G. & Neves, J.C.L. (1995) - Osmocondicionamento, secagem e armazenamento de sementes de Esenbe- ckia leiocarpa England (guarantã). Revista Brasileira de Sementes, 17, 2: 220-226. [ Links ]
Corlett, F.M.F.; Barros, A.C.S.A. & Villela, F.A. (2007) - Qualidade fisiológica de sementes de urucum armazenadas em diferentes ambientes e embalagens. Revista Brasileira de Sementes, 29, 2: 148-158. [ Links ]
Custódio, C.C.; Machado-Neto, N.B.; Caseiro, R.F.; Ikeda, M. & Bomfim, D.C. (2002) - Germinação de sementes de urucum (Bixa orellana L.). Revista Brasileira de Sementes, 24, 1: 197-202. [ Links ]
Heydecker, W.; Higgins, J. & Turner, I.J. (1975) - Invigoration of seeds? Seed Science and Technology, 3: 881-888. [ Links ]
Hillel, D. (1971) - Soil and Water: physical principles and process. Academic Press, New York, 288pp. [ Links ]
Kozlowski, T.T.; Kramer, P.J. & Paltardy, S.G. (1991) - The physiological ecology of Woody plants. Academic Press, San Diego, 657pp. [ Links ]
Larcher, W. (2000) - Ecofisiologia vegetal. RiMa, São Carlos, 531pp. [ Links ]
Lars, S. (2000) - Guide to handling of tropical and subtropical forest seeds. Borch Tryc, Denmark, 512pp . [ Links ]
Lima, R.V.; Lopes, J.C. & Coelho, R.I. (2007) - Germinação de sementes de urucu em diferentes temperaturas e substratos. Ciência e Agrotecnologia, 31, 4: 1219-1224. [ Links ]
Lopes, J.C.; Lima, R.V. & Macedo, C.M.P. (2008) - Germinação e vigor de sementes de urucu. Horticultura Brasileira, 26, 1: 19-25. [ Links ]
Lorenzi, H. (2000) - Árvores Brasileiras. Plantarum, Nova Odessa, 352pp. [ Links ]
Maguire, J.D. (1962) - Speed of germinationaid in selection and evaluation for seedling emergence and vigor. Crop Science, 2, 2: 76-177. [ Links ]
Marcos Filho, J. (2005) - Fisiologia de sementes de plantas cultivadas. Fealq, Piracicaba, 495pp. [ Links ]
Martinazzo, E.G.; Anese, S.; Wandscheer, A.C.D. & Pastorini, L.H. (2007) - Efeito do sombreamento sobre o crescimento inicial e teor de clorofila foliar de Eugenia uniflora Linn (Pitanga) - Família Myrtaceae. Revista Brasileira de Biociências, 5, 2: 162-164. [ Links ]
Nakano, L.C.G. (1998) - Considerações sobre o plantio e a extração do urucum (Bixa orellana L.) e sua utilização como corante. Arquivos de Ciências da Saúde, 2, 1: 33-39. [ Links ]
Nunes, U.R.; Reis, M.S.; Del Giúdice, M.P.; Sediyama, C.S. & Sediyama, T. (2004) - Qualidade fisiológica e sanitária de sementes de soja submetidas ao condicionamento osmótico. Revista Ceres, 51, 294: 163-177. [ Links ]
Perez, S.C.J.G.A. & Negreiros, G.F. (2001) - Efeito do pré-condicionamento na viabilidade e no vigor de sementes de Canafístula (Peltophorum dubium Spreng. Taub.) em condições de estresse. Revista Brasileira de Sementes, 23, 1: 175-183. [ Links ]
Santos, M.C.A.; Aroucha, E.M.M.; Souza, M.S.; Silva, R.F. & Souza, P.A. (2008) - Condicionamento osmótico de sementes. Revista Caatinga, 21, 2: 1-6. [ Links ]
Scalon, S.P.Q.; Mussury, R.M.; Rigoni, M.R. & Scalon Filho, H. (2003) - Crescimento inicial de mudas de Bombacopsis glabra (Pasq.) A. Robyns sob condição de sombreamento. Revista Árvore, 27, 6: 753-758. [ Links ]
Silva, R.R.; Freitas, G.A.; Siebeneichler, S.C.; Mata, J.F. & Chagas J.R. (2007) - Desenvolvimento inicial de plântulas de Theobroma grandiflorum (Willd. ex Spreng.) Schum. sob influência de sombreamento. Acta Amazonica, 37, 3: 365-370. [ Links ]
Smith, P.T. & Cobb, B.G. (1991) - Accelerated germination of pepper seed by priming with salt solutions and water. HortScience, 26, 4: 417-419. [ Links ]
Snedecor, G.W. (1962) - Statistical methods. Iowa State University Press, Ames, 422pp. [ Links ]
Sune, A.D.; Franke, L.B. & Sampaio, T.G. (2002) - Efeitos do condicionamento osmótico na qualidade fisiológica de sementes de Adesmia latifolia (Spreng) Vog. Revista Brasileira de Sementes, 24, 1: 18-23. [ Links ]
Tonin, G.A.; Gatti, A.B.; Carelli, B.P. & Perez, S.C.J.G.A. (2005) - Influência da temperatura de condicionamento osmótico na viabilidade e no vigor de sementes de Pterogyne nitens Tul. Revista Brasileira de Sementes, Pelotas, 27, 2: 35-43. [ Links ]
Valladares, F.; Wright, S.J.; Lasso, E.; Kitajima, K. & Pearcy, R.W. (2000) - Plastic phenotypic response to light of 16 congeneric shrubs from a panamanian rainforest. Ecology, 81, 7: 1925-1936. [ Links ]
Villela, F.A.; Doni-Filho, L. & Sequeira, E.L. (1991). [ Links ] Tabela de potenciais osmóticos em função da concentração de polietilenoglicol 6000 e da temperatura. Pesquisa Agropecuária Brasileira, 26, 11/12: 1957-1968. [ Links ]
Wanli, Z.; Leihong, L. & Perez, S.C.J.G.A. (2001) - Pré-condicionamento e seus efeitos em sementes de canafístula (Peltophorum dubium (Spreng.) Taub.). Revista Brasileira de Sementes, 23, 1: 146-153. [ Links ]
Recebido/Received: 2011.09.14
Aceitação/Accepted: 2012.11.28