











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
Permalink
I. Introduction
In the biosphere, amphibious and coastal tidal mudflat ecosystems are formed by saltmarsh and mangrove habitats. The latter consist of a shrubby or arboreal, helo-halophytic vegetation formation, characteristic of the low-lying muddy areas, such as coastal swamps or lagoon banks, within intertropical environments. Mangrove species are physiologically and morphologically adapted to the amphibian eu-haline environment, withstanding high osmotic pressures and developing aerial root systems and pneumatophores (Moreira, 1984). Unlike mangroves, marshes don’t have arboreal species and colonise mainly temperate zones in the western part of the continents, under the influence of cold sea currents. A marsh is defined as a “characteristic ecosystem of the highest part of the coastal low-lying platforms, occupying the space between the low water neaps and the high water springs. Morphologically it corresponds to the high slike and schorre; it is, therefore, an amphibious ecosystem, with muddy or silty soils, salty, colonized by halo-helophytic herbaceous vegetation. It develops in estuaries, deltas, lagoons and marine swamplands, especially in extratropical regions” (Moreira, 1984, p. 109).
In the intertropical zone of the African continent, mangrove ecosystems are found mainly on the eastern coasts influenced by warm sea currents, as in Mozambique, for instance (Beentje & Bandeira, 2007) where the coastline is influenced by the Mozambique Warm Current, which extends southwards by the Agulhas Current. Mangroves are also common along the coasts of West Africa, from Senegal to Angola (Ward et al., 2016).
Although at a similar latitude to Mozambique, Angola’s coastline is influenced by the Benguela Cold Current and, therefore, the low-energy environments at the mouths of the major watercourses and sheltered inlets are dominated by marshes, although mangroves also occur. However, mangroves on the western coasts of the continents, apart from being less frequent, have a much lower floristic richness than those on the eastern coasts (Moreira, 1977). In Angola, mangroves have shown a very significant decrease due to anthropic action (cutting trees for wood, systematic felling of all vegetation to eradicate diseases, land conversion for agriculture and pollution, among others). In the past, they extended south to Lobito where they are now extinct. Mangroves do not occur in the south of Lobito and along the coast of southwest Angola and Namibia, and the amphibian vegetation of muddy or sandy tidal platforms consists exclusively of saltmarshes.
The occurrence of saltmarshes in the west African coast north of the Cape of Boa Esperança is associated with the Benguela Cold Current, which constitutes a wide flow towards northwest forming the eastern branch of the South Atlantic circulation cell. Typically, cold water from the South Atlantic circulation flows up the west coast of Africa to the latitude of Benguela, about 16° South. The upwelling system known as the Benguela Upwelling System consists of a rise in cold, nutrient-rich water from depths of around 200-300m, which is responsible for the high productivity of Angola’s southwest ecosystems (Kirkman & Nsingi, 2019). However, its influence is irregular as to water characteristics, temperature, northern latitude reach, current intensity, etc. These oscillations are related to two phenomena: on the one hand, frequently warm and nutrient-poor waters from the South Indian Ocean Current, called the Agulhas Current, mix with the Benguela Current, making the thermal (and other) characteristics of the latter chaotic (Veitch & Peven, 2017); on the other hand, about once every 10 years (Mann & Lazier, 2006), during the process known as the Benguela Niño Current (Imbol Koungue et al., 2019), the Angolan Warm Current extends abnormally southwards from the usual 15° S up to 25° S, extending also offshore up to 150km and with a depth of 50m (Richter et al., 2010).
Due to the double influence of the Angolan Hot Current and the Benguela Cold Current on the Angolan coast, we find that all mangroves are located north of Lobito, and, in this entire sector, marshlands are floristically reduced to few succulent species (in most cases only Sarcocornia natalensis subsp. affinis (Moss) S.Steffen, Mucina & G.Kadereit), which are characteristic of marshlands of the inland sector of the mangrove forest, in contact with continental ecosystems. These very impoverished versions of marshes had also been identified for the coast of Mozambique in the inland mangrove sector (Moreira, 1977). In these versions, which Moreira (1977) calls “salty swamps”, are present species as Sesuvium portulacastrum (L.) L. and Sporobolus virginicus (L.) Kunth, among others, which also occur in Angolan marshlands. No mangroves occur south of Lobito, a reality that extends through all the Namibian territory, as in both the southwest of Angola and in the Namibian coast, the tide-influenced muddy or sandy-muddy platforms, where in low energy environments, are colonised by marsh vegetation. Globally, in terms of floristic richness, these marshlands are frankly poorer than the Western Europe ones (and poorer than all the Holarctic Kingdom saltmarshes) of both Mediterranean and temperate oceanic climates (Cardoso, 2014; Costa et al., 2009). This area is characterized by a desert to semi-desert climate, where fresh water supplies are extremely scarce, unlike in the European marshes, especially those in the Eurosiberian Region. In Angola the saltmarshes have their northernmost position in the Cuanza River estuary (near Luanda), where a very impoverished version of these communities occupies a narrow strip in the most inland sector of the mangrove forest, which occures extensively along this river.
The study we are presenting is the first phytosociological analysis of the saltmarshes and brackish marshes of the southwest of Angola and, as such, the communities identified are unprecedented for science. We also make a description of the environmental characteristics in which each community is included, as well as their floristic separation mainly through indicator species.
II. Material and methods
The vegetation relevés were collected according to the sigmatist and dynamic-catenal phytosociology approach (Braun-Blanquet, 1979; Géhu & Rivas-Martínez, 1981; Rivas-Martínez, 1976, 2005; Theurillat et al., 2020). A floristic-statistical group analysis (Müller-Dombois & Ellenberg, 1974) was used for community definitions and synthetic table arrangements. The bioclimatic typology was based on the work of Rivas-Martínez et al. (2011). For the identification of collected specimens, the most relevant bibliography for the area was used, namely Flora Zambesiaca, Flora of Tropical Africa, Flora of Southern Africa, Flora of Tropical East Africa, Conspectus Florae Angolensis (Brummitt, 1992; Exell & Fernandes, 1961; Exell et al., 1970; Exell & Mendonça, 1937, 1951, 1954; Klaassen & Craven, 2003; Ornold & De Wet, 1993; Retief & Herman, 1997; Vollesen, 2000) and their comparison with duly identified herbarium specimens (LISC herbarium, University of Lisbon). The nomenclature used follows the Plants of the World Online database (Plants of the World Online [POWO], 2019).
The Braun-Blanquet scale values were converted to numerical values based on the Müller-Dombois and Ellenberg methodology (1974), and a Principal Component Analysis (PCA) was applied to the matrix containing all the relevés using the Canoco 5.5 software (Ter Braak & Šmilauer, 2012).
III. Characterization of the region
This work was carried out in the coastal saltmarshes and brackish areas of the Namibe Province. This territory comprises the coastal strip of the Namib Desert and is crossed by the Bero and the Pindo rivers with water almost all year round. The Bero estuary is next to the city of Namibe (formerly known as Moçâmedes) and the Pindo estuary near Tombua (formerly known as Porto Alexandre). Occurring in a desert area (mostly a sandy desert), the Angolan saltmarshes in the studied region belong to the silt-loam textural class or even the sandy-loam class (Cardoso et al., 2019).
This area is characterized by a tropical hyperdesert, upper thermotropical, lower hyperarid, attenuated hyper-oceanic bioclimate (fig. 1) (Cardoso, 2014; Rivas-Martínez et al., 2011).
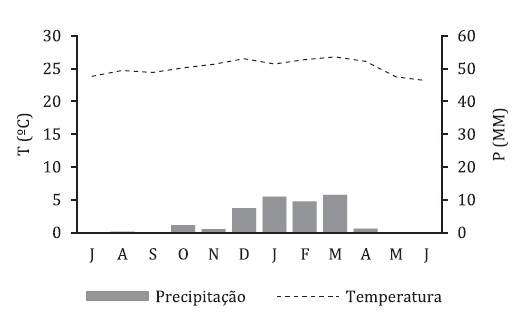
Source: Southern African Science Centre for Climate Change and Adaptative Land Management [SASSCAL] WeatherNet (2020)
Fig. 1 Thermopluviometric diagram for Namibe city (Namibe Weather Station - ID 857 331, Lat. 15 159; Long. 12 178; Alt. 11m; Hyperdesertic tropical, superior thermotropical, inferior hyperarid, attenuated hyperoceanic), 2014-2020.
IV. Results and discussion
1. New associations
The occurrence of 18 species, belonging to eight families (Appendix 1) and six new associations were reported for the saltmarshes and brackish marshes along the coast of the southwest of Angola, which we now describe:
1. Sarcocornietum affinis J.F. Cardoso, J.C. Costa, Neto, Maria C. Duarte & Monteiro-Henriques ass. nov.
(table I, holotypus relevé no. 7)
Halophilous chamaephytic association of Sarcocornia natalensis subsp. affinis, often monospecific, although sometimes Sporobolus virginicus and Sesuvium portulacastrum, may also occur in its floristic composition.
The bioclimate of the areas where it occurs varies from tropical xeric to tropical hyperdesert. We place this community in the Arthrocnemetea franzii class (coastal saltwater marshes flooded by sea tides or occasionally interior saline waters: succulent shrub, forbs and grass communities, growing in infra-thermotropical desertic and xeric bioclimate of tropical west coast of Africa), order Sarcocornio-Salicornietalia meyerianae (halophytic vegetation of small succulent plants); Sarcocornion decumbentis alliance (flooded communities; Boucher & Jarman, 1977; Hanekom et al., 2009; Mucina et al., 2003; Rivas-Martínez et al., 2017).
Sarcocornia natalensis subsp. affinis was first reported for Angola by Cardoso (2014) as part of the fieldwork conducted during this study. This species was listed in Plantas de Angola (Figueiredo & Smith, 2008) as Arthrocnemum macrostachyum (currently Arthrocaulon macrostachyum). However, the genus Arthrocnemum has one undivided cavity in the axis after the detachment of the flowers while the specimens we observed, in the Namibe saltmarshes, have three cavities which correspond to the genus Sarcocornia (S. natalensis subsp. affinis). It is a prostrate, rhizomatous chamaephyte, woody at the base, succulent, with segments of 5-15mm long and 2-3mm in diameter, corresponding to the description made by Le Roux and Wahl (2005).

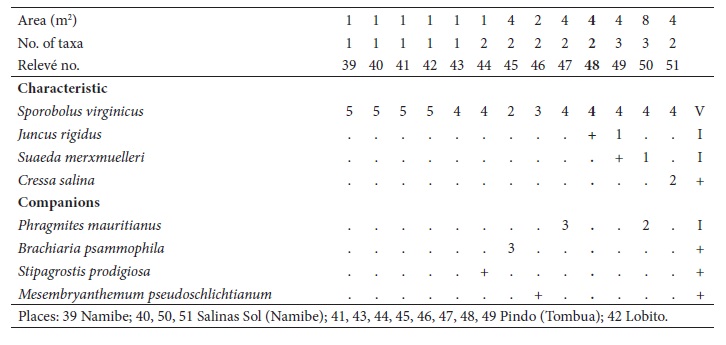
2. Suaedetum merxmuelleri J.F. Cardoso, J.C. Costa, Neto, Maria C. Duarte & Monteiro-Henriques ass. nov.
(table II, holotypus relevé no. 52)
This plant community, with a distribution in tropical desertic and hyperdesertic, upper thermotropical, ultrahyperarid to hyperarid, attenuated hyperoceanic (semi-hyperoceanic) bioclimate, is characterized, almost exclusively, by Suaeda merxmuelleri, an endemic nanophanerophyte (bush) of the southwest of Angola, sometimes accompanied by Sporobolus virginicus, Sesuvium portulacastrum and Juncus rigidus. It is found in the highest areas of the saltmarsh, with a brackish water table, and it is only occasionally flooded, sometimes by brackish waters after heavy rains. Being a desert with tropical characteristics, the soil is subjected to strong salinization for most of the year. Salinity oscillations can be high although low values can be recorded very sporadically during the scarce rainy periods. It is a frequent association in nitrified sites (with anthropic disturbance) along roads and paths. Very common in the studied area, also occurring further in the north, in Lobito marshes. We have placed this community in Arthrocnemetea franzii, Sarcocornio-Juncetalia kraussii (halophilous nanophanerophytic vegetation), Spergulario mediae-Puccinellion angustae (dense communities rich in Amaranthaceae and halotolerant grasses, adapted to brackish water flooding) (Boucher & Jarman, 1977; Hanekom et al., 2009; Mucina et al., 2003; Rivas-Martínez et al., 2017).
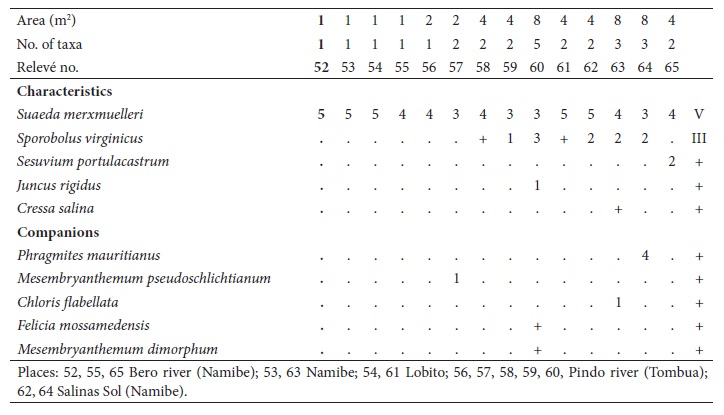
3. Cypero laevigati-Cressetum salinae J.F. Cardoso, J.C. Costa, Neto, Maria C. Duarte & Monteiro-Henriques ass. nov.
(table III, holotypus relevé no. 3)
This community is dominated by halophilous annual (therophytes) and stoloniferous (hemicryptophytes) species, with fugacious hydromorphy (only when heavy rainfall occurs), inserted in a tropical hyperdesert, upper thermotropical, lower ultrahyperarid, attenuated hyperoceanic (semi-hyperoceanic) bioclimate. It is characterized by Cressa salina, Sporobolus virginicus and Cyperus laevigatus. This community is infrequent, having been observed only in moderately nitrified saline habitats with short flood periods, near the city of Namibe. We consider it to be a geovicarious of the Cressetum salinae association, present in Cabo Verde, where it also occurs in temporarily flooded sandy soils (Rivas-Martínez et al., 2017). This association has been positioned in the Arthrocnemetea franzii class, Sarcocornio-Juncetalia kraussii order, Spergulario mediae-Puccinellion angustae alliance.
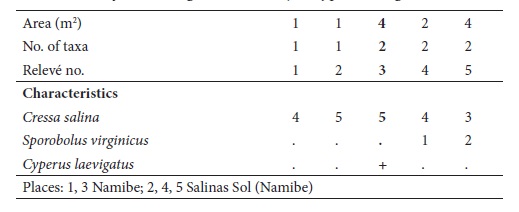
4. Junco rigidi-Sporoboletum virginici J.F. Cardoso, J.C. Costa, Neto, Maria C. Duarte & Monteiro-Henriques ass. nov.
(table IV, holotypus relevé no. 48)
This community was observed in occasionally flooded depressions, in solonchak arenic soils with very occasional hydromorphy (characterized by a strong concentration of soluble salts due to strong evaporation, in desert and semi-desert climates). It is composed almost exclusively by Sporobolus virginicus, sometimes accompanied by Juncus rigidus, and, in the higher areas, by Suaeda merxmuelleri. In areas where the flooding period is longer, the lower salinity allows the occurrence of reed (Phragmites mauritianus). Besides the study area, we also observed this community further north, at Lobito. This community occurs in tropical hyperdesert, upper thermotropical, ultrahyperarid to hyperarid, hyperoceanic attenuated (semi-hyperoceanic) bioclimate, although it can occur in tropical desert and xeric bioclimate too. It has been placed in the Arthrocnemetea franzii class, Sarcocornio-Juncetalia kraussii order and Spergulario mediae-Puccinellion angustae alliance.
5. Sesuvietum crithmoidis J.F. Cardoso, J.C. Costa, Neto, Maria C. Duarte & Monteiro-Henriques ass. nov.
(table V, holotypus relevé no. 23)
Halonitrophilous, psammophilous and monospecific community of the perennial Sesuvium crithmoides (= S. mesembryanthemoides, = S. crystallinum) sometimes accompanied by Zygophyllum simplex (table V). It occurs on intertidal sands, flooded by oceanic waters usually at maximum tidal level, where the sea deposits organic (responsible for the nitrification) and inorganic debris. The relevés were collected in areas characterized by tropical hyperdesertic and desertic, thermotropical, ultrahyperarid to arid, hyperoceanic attenuated (semi-hyperoceanic) bioclimate. According to our observations, this community occurs along the maritime dunes at least up to Luanda (tropical xeric bioclimate).
The taxon Sesuvium crithmoides, previously considered endemic in Angola, was recently (Sukhorukov et al., 2018) observed on the sandy coast of the Democratic Republic of Congo.
We have placed this association in the Arthrocnemetea franzii class, Sesuvietalia persoonii order (communities of succulent stolonate, hydro-halophytic and aero-halophytic species from the western coasts of Africa, from Morocco to South Africa, in infra-thermotropical, ultrahyperarid to arid bioclimate) and Sesuvion persoonii alliance (see Costa, 2018; Rivas-Martínez et al., 2017).
6. Sarcocornio affinis-Sesuvietum portulacastri J.F. Cardoso, J.C. Costa, Neto, Maria C. Duarte & Monteiro-Henriques ass. nov.
(table VI, holotypus relevé no. 37)
Halophilous community dominated by Sesuvium portulacastrum, a perennial, succulent, pioneer species, of pantropical distribution, accompanied by Sarcocornia natalensis subsp. affinis, typical of saltmarshes and sandy depressions flooded and submerged by brackish waters for more or less long periods. It occurs in tropical hyperdesertic and desertic, thermotropical, ultrahyperarid to hyperarid, hyperoceanic attenuated (semi-hyperoceanic) bioclimate. According to our observations it occurs in Lobito marshes and southward until the river Cunene estuary, almost always in monospecific formations. It is a geovicarious of the Cape Verde association Sesuvietum portulacastri, also dominated by Sesuvium portulacastrum (Rivas-Martínez et al., 2017), however, in SW Angola this species is accompanied by Sarcocornia natalensis subsp. affinis and Sporobolus virginicus, which do not occur in Cape Verde. We also include this community in the Sesuvietalia persoonii order and Sesuvion persoonii alliance.

2. Multivariate analysis (ordination) of the relevés
The relevés were collected in a table and the Braun-Blanquet scale values were converted into numerical values to be used in a Principal Component Analysis (PCA). The purpose of this analysis was primarily to determine the dispersion patterns of the relevés as a result of their floristic composition. The closest points correspond to similar relevés in terms of floristic composition and the farthest points correspond to more dissimilar relevés. In this way we can test the manual organisation of the relevés that has been carried out with the previously presented tables and the respective plant associations. The observed structure, although being the result of the floristic composition of the relevés, is also the result of the implicit effect of the environmental factors, which were discussed in the description of each of the plant communities.
Figures 2 and 3, resulting from the PCA, show a good segregation of the plant communities described (fig. 2) and clearly characterized by the bioindicator species presented (fig. 3). Suaeda merxmuelleri is the diagnostic species of the association Suaedetum merxmuelleri; Sporobolus virginicus is the diagnostic species of the association Junco rigidi-Sporoboletum virginici; Sarcocornia affinis is the diagnostic species of the association Sarcocornietum affinis; Sesuvium portulacastrum is the diagnostic species of the association Sarcocornio affinis-Sesuvietum portulacastri; Cressa salina is the diagnostic species of the association Cypero laevigati-Cressetum salinae; and Sesuvium crithmoides is the diagnostic species of the association Sesuvietum crithmoidis.
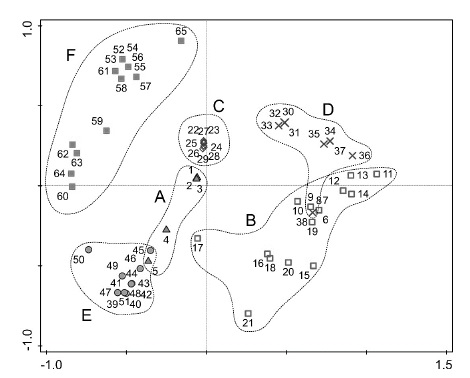
Fig. 2 Cypero laevigati-Cressetum salinae (samples 1-5); B. Sarcocornietum affinis (samples 6-21); C. Sesuvietum crithmoidis (samples 22-29); D. Sarcocornio affinis-Sesuvietum portulacastri (samples 30-38); E. Junco rigidi-Sporoboletum virginici (samples 39-51); F. Suaedetum merxmuelleri (samples 52-65)Principal Component Analysis (PCA) - plot showing the ordination of vegetation samples.
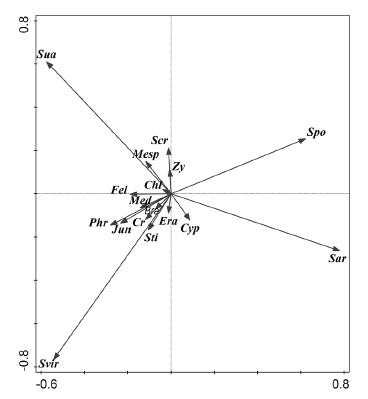
Fig. 3 Chl (Chloris flabellata); Cr (Cressa salina); Cyp (Cyperus laevigatus); Era. (Eragrostis prolifera); Fel (Felicia mossamedensis); Jun (Juncus rigidus); Bra (Brachiaria psammophila); Phr (Phragmites mauritianus); Med (Mesembryanthemum dimorphum); Mesp (Mesembryanthemum pseudoschlichtianum); Sar (Sarcocornia natalensis subsp. affinis); Scr (Sesuvium crithmoides); Spo (Sesuvium portulacastrum); Svir (Sporobolus virginicus); Sti (Stipagrostis prodigiosa); Sua (Suaeda merxmuelleri); Zy (Zygophyllum simplex).Principal Component Analysis - plot showing the ordination of species.
Syntaxonomical scheme (in bold, new syntaxa described in the context of this work):
Class: Arthrocnemetea franzii Rivas-Martínez, Lousã, J.C. Costa & Maria C. Duarte 2017
Order: Sesuvietalia persoonii Rivas-Martínez, Lousã, J.C. Costa & Maria C. Duarte 2017 corr. Rivas-Martínez, Lousã, J.C. Costa & Maria C. Duarte in J.C. Costa 2018
Alliance: Sesuvion persoonii Lousã, J.C. Costa & Maria C. Duarte 2017 corr. Rivas-Martínez, Lousã, J.C. Costa & Maria C. Duarte in J.C. Costa 2018
Sesuvietum crithmoidis J.F. Cardoso, J.C. Costa, Neto, Maria C. Duarte & Monteiro-Henriques (new)
Sarcocornio affinis-Sesuvietum portulacastri J.F. Cardoso, J.C. Costa, Neto, Maria C. Duarte & Monteiro-Henriques (new)
Order: Sarcocornio-Salicornietalia meyerianae Boucher ined.
Alliance: Sarcocornion decumbentis Mucina, Jansen & O’ Callaghaen 2003
Sarcocornietum affinis J.F. Cardoso, J.C. Costa, Neto, Maria C. Duarte & Monteiro-Henriques (new)
Order: Sarcocornio-Juncetalia kraussii Boucher ined.
Alliance: Spergulario mediae-Puccinellion angustae Mucina, Jansen & O’Callaghan 2003
Suaedetum merxmuelleri J.F. Cardoso, J.C. Costa, Neto, Maria C. Duarte & Monteiro-Henriques (new)
Cypero laevigati-Cressetum salinae J.F. Cardoso, J.C. Costa, Neto, Maria C. Duarte & Monteiro-Henriques (new)
Junco rigidi-Sporoboletum virginici J.F. Cardoso, J.C. Costa, Neto, Maria C. Duarte & Monteiro-Henriques (new)
V. Conclusions
The coastal region of the southwest of Angola (south of Lobito), despite being under a tropical macrobioclimate, presents no mangroves in its coastal swamps. Instead, saltmarshes are present, much probably owing to the influence of the cold oceanic waters of the Benguela Cold Current. These marshes are floristically very poor and often monospecific. This is a substantially different situation from the South African and Holarctic marshes characterized by a much greater floristic richness. Angolan marshlands are characterized by a dominance of fine sandy soil texture which is associated to the presence of the extensive field of dunes typical of the Namib Desert. The research carried out during 2010-2015 and 2018 allowed to analyse the floristic composition of the Namibe province’s marshlands, as well as the originality of the floristic combinations associated to the more or less differentiated environments.
In this way, a total of six new plant associations were described. Their spatial distribution is related to the ecological gradients associated to water table salinity, time of submersion by sea waters, soil texture, as well as nitrogen levels. These communities, clearly segregated by the PCA, do not present a seral logic and, therefore, as a whole, constitute an original geopermasigmetum, presented for the first time in this work.


