Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
Permalink
I. Introdução
Em Portugal, e de um modo geral na Península Ibérica, os teixedos (quase) não foram objeto de estudo até à sua individualização como Habitat (prioritário), no âmbito dos trabalhos da Diretiva Habitats, nos anos 90. A ficha do Habitat 9580*, para Portugal continental, demonstra a então falta de conhecimento concreto destes bosquetes (Instituto da Conservação da Natureza e das Florestas [ICNB], 2005). Por um lado, com o teixo em clara regressão na Península (Lence et al., 2011), as áreas com teixedos são raras e normalmente de pequena extensão, ocorrendo sobretudo em territórios recônditos e de difícil acesso. Por outro, tratando-se de bosquetes (co)dominados por um táxon relicto, de lento crescimento, capaz de sobreviver em ambientes sombrios, torna a sua classificação mais complexa, já que é uma vegetação descontextualizada às nossas latitudes (Honrado et al., 2001). De resto, o mesmo se passa com outros tipos de vegetação dominados por elementos relictos, e.g. Ilex aquifolium (Rodríguez Guitián et al., 2011) ou Prunus lusitanica (Calleja Alarcón, 2006).
1. Distribuição do Teixo e dos Teixedos (passada e presente)
Do conhecimento da distribuição de Taxus baccata na Península assinalam-se os territórios atlânticos, assim como os principais sistemas montanhosos ibéricos, como aqueles onde há maiores evidências da sua ocorrência (atual e passada). Os registos advêm não só de levantamentos florísticos (Anthos, n.d.; Carvalho et al., 1999; Flora-On, 2012-2019; SIVIM, n.d.), mas também da fitotoponímia (Carrillo López et al., 2010) ou de dados paleobotânicos. As análises polínicas apontam para o seu declínio na Península desde a última glaciação, com um apogeu entre 6000 e 3000 BP (Peñalba, 1994, cit. in Lence et al., 2011). No NW ibérico assinalam-se pólenes de teixo, e.g., na Serra da Estrela (van der Knaap & van Leeuwen, 1995), Lugo (Carrillo López et al., 2010), Sanábria (Sobrino et al., 2004), mas curiosamente não na Serra do Gerês (Ramil-Rego et al., 1998). A sua ocorrência pode ser negligenciada ou menosprezada, caso haja problemas na identificação do pólen ou quando não tiver deixado rastro polínico detetável (Carrillo López et al., 2010). Já na Serra do Marão detetaram-se fragmentos de carvão (Figueiral & Carcaillet, 2005).
Através da fitotoponímia é evidente o desaparecimento de teixo em territórios na Galiza (região com mais fitotopónimos) e em Portugal (Carrillo López et al., 2010), onde, consoante o critério chegaria ao Alto Alentejo (Draper & Marques, 2007) ou Baixo Alentejo (Carrillo López et al., 2010). Na Estremadura só haveria na Serra da Arrábida, ainda que os dados históricos de herbário o identifiquem aqui como cultivado (Anthos, n.d.). A dificuldade tanto na recolha como na interpretação (confusões com zootopónimos ou antrotopónimos mais recentes; Draper e Marques, 2007), ou com espécimes cultivados dado os seus múltiplos usos ancestrais) leva a que se deva ter cautelas acrescidas na sua interpretação.
Modelos ecológicos (Draper & Marques, 2007; Pina Manique e Albuquerque, 1982) apontam também para uma distribuição potencial do teixo em Portugal continental mais alargada do que hoje se conhece: serras do Gerês, Estrela, e Caramulo onde resta um único espécime (Ribeiro & Paiva, 2005). Para Pina Manique e Albuquerque (1982) o teixo (e o bidoeiro) é considerado silva climática das serras de todo N e C, e ainda nas de S. Mamede e do Caldeirão. Já com a modelação do nicho ecológico de Draper e Marques (2007) fica restrito a algumas das serras do NW (Peneda-Gerês, Cabreira, Alvão, Marão, Montemuro), alcançando a Serra da Estrela e incluindo o Caramulo, mas, curiosamente, não a Serra de Montesinho, onde a extinção recente não se confirma (Aguiar, 2000; Cortés et al., 2000; Draper & Marques, 2007), pois o único exemplar de herbário, recolhido em 1877 por M. Ferreira, e em que se basearam os botânicos portugueses (Aguiar, 2000), foi afinal no “Teixedello [de Requejo de Sanábria], pr. de Montesinho” (Aguiar, 2000, p. 78), i.e. ca. 4km a N da fronteira, já em Espanha.
Apesar de poder haver problemas de interpretação de alguns dos dados, não parece haver dúvidas da presença histórica de teixo para além das serras onde hoje existe (Catarino, 2007). Contudo, a sua presença não indica obrigatoriamente a ocorrência de teixedos, já que estes bosquetes teriam certamente uma distribuição mais restrita que os indivíduos isolados (e cultivados). E.g., em Itália, sob clima mais continental, o teixo deixa de formar “povoamentos densos”, que só ocorrem nas áreas mais oceânicas (Abbate et al., 2003).
2. Interpretação dos Teixedos (e Habitat 9580*)
As primeiras abordagens aos teixedos em Espanha passaram pela enumeração ou análise dos inventários que continham Taxus baccata (Cortés et al., 2000; Lence et al., 2011; Serra, 2009). Em Portugal, essa enumeração foi feita recentemente (Portela-Pereira et al., 2014, 2016) no âmbito do projeto LIFE TAXUS 2013-2016 (Associação Nacional de Conservação da Natureza [QUERCUS], 2016). Já no resto da Europa descreveram-se mais de uma dúzia de sintáxones dominados por Taxus baccata (Paule et al., 1993; Rodwell, 1991, cit. in Rodríguez Guitián et al., 2011) e, posteriormente, alguns mais foram classificados na Sardenha (Bacchetta & Farris, 2007; Farris et al., 2012) e também no Irão (Esmailzadeh et al., 2007). Por sua vez, na Península Ibérica são conhecidas apenas duas associações em que o teixo pode ser dominante (Rodríguez Guitián et al., 2011): Saniculo europaeae-Taxetum baccataBólos 1967 (Bòlos, 1967, cit. in Rodríguez Guitián et al., 2011) e Ononido aragonensis-Tilietum platyphylli (Pitarch, 2002, in Crespo et al., 2008). A primeira refere-se a teixedos de montanhas costeiras e pré-pirenaicas na Catalunha (Rivas-Martínez et al., 2001, 2011), a segunda, corresponde a bosques de Tilia em que o teixo pode ser codominante, no SE do Sistema Ibérico (Crespo et al., 2008). Há ainda alguns sintáxones de nível inferior que foram delimitados tendo em conta a presença de teixo (Rodríguez Guitián et al., 2011).
O único estudo na Península Ibérica que analisou inventários de teixedos foi o de Rodríguez Guitián et al. (2011) no W da Cordilheira Cantábrica. Duas interpretações foram apresentadas: i) atender exclusivamente ao aspeto atual dos teixedos, ou ii) considerar a eliminação recente de elementos arbóreos de altura superior ao teixo. Dada a complexa interpretação fitossociológica dos teixedos, a opção dos autores foi a 2.ª, interpretando-os como faciações dos bosques zonais em que estão “inseridos”, como os faiais ou bosques mistos com Acer pseudoplatanus e Corylus avellana (Rodríguez Guitián et al., 2011). No entanto, com a análise preliminar de novos inventários de teixedos em Portugal (Portela-Pereira et al., 2016), que completam a sua distribuição no NW ibérico, considera-se possível outra interpretação. Até porque, considerar inúmeras faciações de teixo como Habitat da Rede Natura torna-se, evidentemente, demasiado complexo, já que este ocorre isoladamente em diversos sintáxones (Lence et al., 2011), sem que se esteja na presença de um verdadeiro teixedo e do respetivo Habitat 9580*, i.e., bosquetes (co)dominados por Taxus baccata.
Sendo assim, o objetivo deste trabalho é classificar os remanescentes dos bosques de Taxus baccata L. no NW da Península Ibérica, comparando os inventários do W da Cordilheira Cantábrica, já publicados, e os inéditos e recém-inventariados das serras do Gerês e da Estrela, em Portugal.
II. Materiais e métodos
Este trabalho centrou-se num total de 33 inventários fitossociológicos de teixedos, dos quais onze provêm da Serra do Gerês e três da Serra da Estrela, Portugal (Honrado, 2003; Portela-Pereira et al., 2016) e os restantes do W da Cordilheira Cantábrica em Espanha (Ortiz, 1986, cit. in Rodríguez Guitián et al., 2011; Rodríguez Guitián et al., 2011; Romero Rodríguez, 1993).
Ao nível taxonómico segue-se a Flora iberica (Castroviejo, 1986-2015) e a Nova Flora de Portugal (Franco, 1984; Franco & Rocha Afonso, 1998), tendo sido seguida a Flora-On (2012-2019) para alguns casos de atualizações nomenclaturais mais recentes. No caso de táxones não abordados por estas obras seguiu-se a interpretação dos autores dos inventários (Rodríguez Guitián et al., 2011; Romero Rodríguez, 1993). Devido a diferentes interpretações entre autores, foi realizada uma generalização taxonómica, não entrando na análise estatística alguns táxones infraespecíficos, considerando-se em sentido lato: Arrhenatherum elatius, Betula pubescens, Digitalis purpurea, Dryopteris affinis, Galium saxatile, Lonicera periclymenum e Luzula sylvatica. Táxones não completamente determinados até ao nível de espécie foram retirados da análise numérica DiffVal, uma vez que poderiam promover artificialmente a criação de grupos, com a exceção de Rubus spp., onde se incluíram outros táxones que não Rubus ulmifolius e Rubus idaeus. Finalmente, retiraram-se da análise os táxones exóticos (no caso, Prunus laurocerasus), mantendo-se os táxones sobre os quais recaem algumas dúvidas sobre serem nativos de Portugal, nomeadamente: Acer pseudoplatanus (S. Estrela), Castanea sativa, e Betula pubescens subsp. carpatica (S. Estrela) que ficou incluído em Betula pubescens.
A sintaxonomia está de acordo com o trabalho-síntese de Costa et al. (2012), com as atualizações propostas por Mucina et al. (2016).
Para a análise dos inventários recorreu-se às ferramentas de optimização aplicadas à classificação de vegetação e tabulação fitossociológica, usando os índices DiffVal1 e TotDiffVal1 baseadas no conceito de “espécie diferencial” (Monteiro-Henriques, 2016; Monteiro-Henriques & Bellu, 2014) implementados em ambiente R (R Core Team, n.d.), comparando os seus resultados com uma classificação aglomerativa através método de Ward, utilizando a dissemelhança de Bray-Curtis. Para a comparação dos resultados obtidos pelas ferramentas de optimização e pela classificação aglomerativa são criadas imagens da matriz ordenada (atribuindo uma cor sempre que o táxon ocorre num determinado grupo), usando a função tabulate. Esta função devolve também uma imagem da matriz ordenada num formato condensado (onde cada bloco de táxones diferenciais é evidenciado) e a distribuição dos inventários num mapa. A função tabulate ordena os táxones, primeiro pelo número de grupos em que estes ocorrem e, segundo, pela sua frequência relativa ajustada (Monteiro-Henriques, 2016).
Nesta análise utilizou-se uma matriz binária (i.e. presença-ausência), tendo sido retirados os táxones monopresentes (i.e., táxon que só ocorre num único inventário), obtendo-se assim uma matriz de 134 táxones x 33 inventários.
III. Resultados
1. Composição Florística
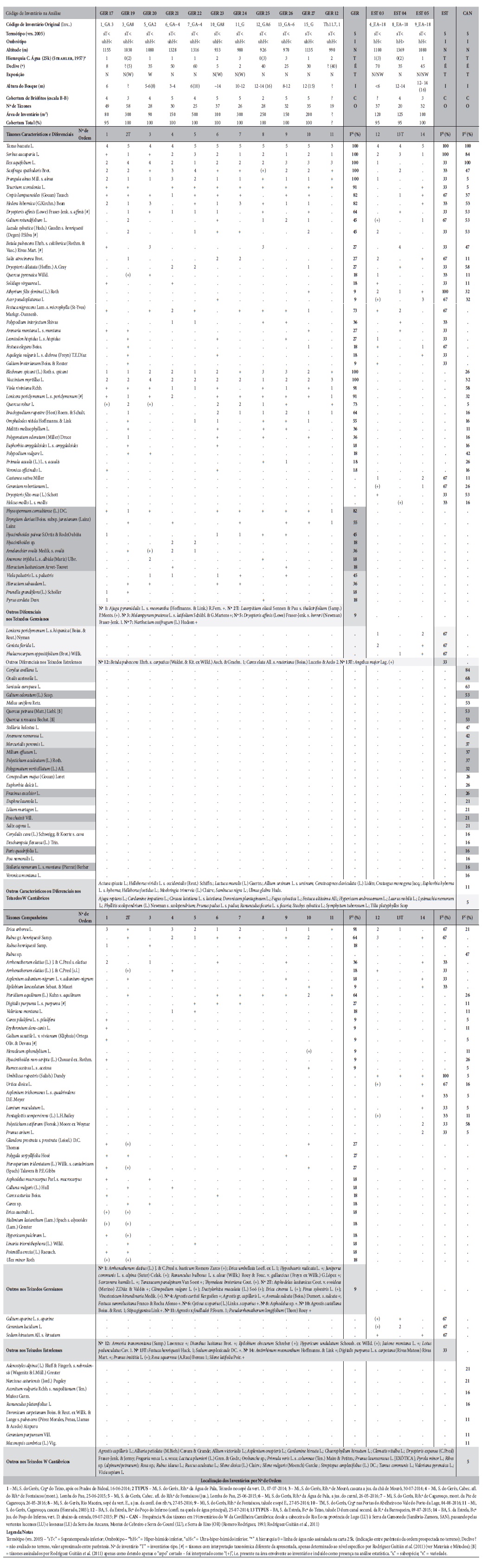
Nesta secção apresentam-se os principais resultados. Na solução aglomerativa, onde o método de classificação utilizado não produz resultados por otimização, a obtenção de três grupos (A1, A2 e A3) baseia-se na separação entre os inventários lusitanos e os inventários W cantábricos, dos quais se individualiza um pequeno terceiro grupo com a separação de três dos inventários do Teixedelo de Requejo de Sanábria (A3; fig. 1). Analisando este grupo, observa-se que não possui táxones diferenciais face aos restantes inventários do W cantábrico (fig. 1, colunas A3). Devido ao método de Ward + Bray-Curtis, que faz a agregação de inventários aos pares, promove-se a criação de um grupo de inventários similares entre si, mas sem táxones diferenciais que o caracterizem. Comparando as figuras 1 e 2, com a optimização do índice TotDiffVal1 (fig. 2) obtém-se três grupos distintos de teixedos (B1, B2 e B3), mas desta feita com diferenciação biogeográfica e táxones diferenciais em todos os grupos (com base na matriz). Estes são bem visíveis nos primeiros três blocos de colunas na matriz que ladeia o mapa, assim como são também os táxones comuns aos três grupos (fig. 2), dos quais se destacam, para além de Taxus baccata, como muito frequentes nos teixedos NW ibéricos, Sorbus aucuparia e Ilex aquifolium (Anexo I). Com a optimização do TotDiffVal1 consegue-se assim distinguir teixedos estrelenses, geresianos e W cantábricos. Os táxones diferenciais entre os teixedos W cantábricos e os lusitanos são inúmeros, já que diversos táxones têm o seu limite nesta cordilheira (e.g. Galium odoratum, Quercus petraea e seu híbrido; Anexo I), e outros só ocorrem de forma finícola em território português, não surgindo nos teixedos lusos inventariados (e.g. Oxalis acetosella ou Anemone nemorosa). Por outro lado, nos teixedos W cantábricos é frequente Corylus avellana, que ocorre em Portugal a uma altitude menor que a dos teixedos, onde surge associado a bosques mesotróficos de solos profundos e frescos dos sopés das vertentes (mesotemperados).
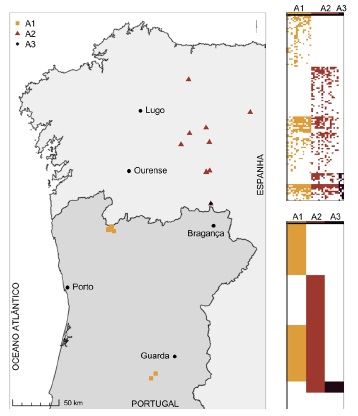
Fig. 1 Tabulação, usando a função tabulate, para o resultado da classificação aglomerativa (método de Ward, Bray-Curtis, matriz binária sem táxones monopresentes) para três grupos (TotDiffVal1 para a solução 3 grupos = 0,1525837). Figura a cores disponível online.
Fig. 2 Tabulação, usando a função tabulate, do resultado da otimização do índice TotDiffVal1 para três grupos (matriz binária, sem táxones monopresentes); valor TotDiffVal1 máximo encontrado = 0,1758473. Figura a cores disponível online.
Dos teixedos do Gerês são diferenciais, e.g. Eryngium juresianum, Hyacinthoides paivae, Amelanchier ovalis ou Anemone albida (Physospermum cornubiense é conhecido na S. da Estrela em bidoal supratemperado (Rivas-Martínez, 1981), muito próximo dos teixedos inventariados; Anexo I).
Porém, a separação dos teixedos estrelenses é bem mais complexa, não só pelo reduzido número de inventários disponíveis, dada a raridade do próprio Taxus baccata nesta serra (Portela-Pereira, 2019), como também pelo estado de conservação dos pequenos teixedelos existentes. Os táxones diferenciais evidenciados nos inventários, essencialmente transgressivos a partir da matriz de vegetação envolvente, são disso amostra, acusando claramente a prevalência de condições de orla, correspondendo a elementos heliófilos como Genista florida ou Phalacrocarpum oppositifolium, que também ocorrem na Serra do Gerês, mas onde são pouco frequentes na área dos teixedos, não surgindo nos inventários efetuados.
Outro táxon que poderá ser um verdadeiro diferencial, não utilizado na análise numérica devido a diferentes interpretações taxonómicas, é Lonicera periclymenum subsp. hispanica, que será um táxon ausente (ou muito pouco frequente) na Serra do Gerês, em detrimento de Lonicera periclymenum subsp. periclymenum (Flora-On, 2012-2019; Honrado, 2003).
Há ainda outros elementos relevantes (com elevado valor biogeográfico e também ecológico - elementos rupícolas) nos inventários estrelenses, que não ocorrem no Gerês, assinalados na parte final do anexo I. Ainda assim, a separação dos teixedelos estrelenses faz-se sobretudo pela falta de táxones que ocorrem no Gerês. Tendência que, de resto, se reflete desde o W cantábrico: com a mediterraneidade a aumentar para S, vários táxones nemorais atlânticos vão perdendo habitat, reflexo do seu limite meridional de distribuição. Por outro lado, o aumento da mediterraneidade provoca a subida em altitude dos teixedos, perdendo-se o habitat para outras espécies menos orófilas. Por razões climáticas, ou pelo elevadíssimo grau de hemerobia dos bosques nativos estrelenses, vários táxones são hoje raros ou ausentes no supratemperado estrelense: e.g., Vaccinium myrtillus, Blechnum spicant, Quercus robur, Omphalodes nitida, Polygonatum odoratum. Neste sentido propõe-se que a separação entre os teixedos geresianos e estrelenses se reconheça ao nível de subassociação.
2. Características ambientais dos teixedos
Sintetizado no cabeçalho do anexo I, os teixedos lusitanos surgem de forma relicta nos distritos “Serras da Peneda e Gerês” e “Serrano Estrelense” (genericamente correspondente, respetivamente, ao subsetor Geresiano-Queixense e ao setor Estrelense de Costa et al. (1999), que pertencem à nova subprovíncia Orolusitana Atlântica, no SW da Região Eurossiberiana (Rivas-Martínez et al., 2017). Bioclimaticamente, o teixedo geresiano ocupa sobretudo o supratemperado inferior eutemperado, mas na face E do Gerês é já submediterrânico, tal como o teixedo estrelense [C.B.T. de Rivas-Martínez, versão 2005 (Monteiro-Henriques, 2010)]. A presença no supratemperado superior será atualmente residual dada a falta de condições edáficas e topográficas vs. ação antrópica, tanto na Estrela como no Gerês (pequena área no cume). No andar mesotemperado superior, o teixo ocorre sobretudo isolado, não se detetando bosquetes consistentes para inventário (Portela-Pereira et al., 2016). Em termos ômbricos, o teixedo geresiano diferencia-se do estrelense, já que se desenvolve no ultra-hiper-húmido, enquanto o segundo no hiper-húmido (Monteiro-Henriques, 2010). Assinala-se ainda a dominância da exposição ao quadrante N (e W).
Os teixedos surgem em Portugal em terrenos siliciosos, de baixa mineralização, nomeadamente granitos (e corneanas, na S. da Estrela) em “Rios Montanhosos do Norte” (Tipo M) (Instituto da Água [INAG], 2008). Estes cursos de água de pequena dimensão, com leito rochoso e pedregoso, possuem declives acentuados e ocorrem em altitudes elevadas no contexto nacional (os teixedos inventariados estão entre os 920 e os 1370m), possuindo elevado escoamento médio anual (INAG, 2008). No entanto, os teixedos não colonizam estes cursos de água em toda a sua extensão, habitando sobretudo no leito rochoso/pedregoso ou nas margens de corgas e barrocas, i.e., cursos de água com hierarquia de ordem 1 ou 2 (Strahler, 1957), e nas margens declivosas de pequenos ribeiros de ordem 3, embora já mais raramente. Assim, mais do que em fluvissolos, que já começam a formar-se em cursos de ordem 3, é nos leitos pedregosos (dominados por grandes blocos) e nos coluvissolos que os teixedos ocorrem. Em suma, os teixedos encontram-se hoje sobretudo refugiados em cursos de água essencialmente torrenciais, rochoso-coluviais, onde a dinâmica de vertentes acaba por dominar sobre a dinâmica fluvial (Portela-Pereira et al., 2016).
Na Serra do Gerês, onde os teixedos são mais abundantes (Carvalho et al., 1999), não se encontram nos cursos de água principais (e.g. Rio Homem, Rio Fafião ou Rio de Cabril - ordem 4 e 5), estando presentes sobretudo teixos isolados integrados em bosquetes de folhosas. A ocorrência de possíveis teixedos nestes rios estará limitada a trechos com canal menos instável, e.g., com menor declive ou com obstáculos rochosos, mais protegidos da dinâmica fluvial (Portela-Pereira et al., 2016). Em vales íngremes acabam por se estender pelo sopé das vertentes, usufruindo de coluviossolos profundos abastecidos por escoamento (sub)superficial (Portela-Pereira et al., 2016). Ocorrem ainda espécimes isolados de teixo em diversos habitat (rupícolas, orófilos, mesófilos ou paludo-turfófilos; Portela-Pereira et al., 2016), demostrando a grande amplitude ecológica desta espécie. Os teixedos lusitanos ocorrem, assim, associados a territórios onde dominam solos do tipo ranker (Carvalho Cardoso et al., 1971), i.e., leptossolos na World Reference Base for Soil Resources (WRB; IUSS, 2007).
No que respeita aos teixedos do W cantábrico, estes ocorrem entre os (630)1180 e os 1415m de altitude, em vertentes com forte inclinação e expostas a N (Rodríguez Guitián et al., 2011). A litologia predominante são rochas metamórficas siliciosas, sendo que apenas um inventário corresponde a calcários e dolomitos (Rodríguez Guitián et al., 2011). Os solos caracterizam-se, tal como os lusitanos, pela sua elevada pedregosidade e abundância de afloramentos rochosos, classificáveis como leptossolos e umbrissolos segundo a WRB (Rodríguez Guitián et al., 2011). No entanto, sobre as características hidrogeomorfológicas e bioclimáticas as informações recolhidas são muito escassas, apenas se salienta que no teixedo de Requejo de Sanábria os bosquetes surgem nas proximidades de “umbrias y vaguadas” (Romero Rodríguez, 1993).
IV. Discussão
1. Problemas na interpretação sintaxonómica dos teixedos
Considera-se que os problemas de interpretação sintaxonómica dos teixedos na Península Ibérica (Cortés et al., 2000; Rodríguez Guitián et al., 2011), bem como do Habitat 9580*, encontram justificação nos seguintes aspetos:
O facto de Taxus baccata ser um táxon relicto em regressão (Lence et al., 2011) promoveu o desconhecimento dos seus pequenos e raros bosquetes. Tal facto estava refletido no número reduzido de inventários e estudos fitossociológicos que até há pouco tempo se encontravam disponíveis. Em Portugal conhecia-se um único inventário com Taxus baccata codominante, da Serra do Gerês (Honrado, 2003; nº 11 do quadro I). Em Espanha, eram igualmente residuais (Lence et al., 2011; Rodríguez Guitián et al., 2011), embora se conhecessem alguns, poucos, inventários de teixedos (Bòlos, 1967; Romero Rodríguez, 1993), ou de comunidades codominadas por Taxus baccata (Pitarch García, 2002; Rivas Goday & Borja, 1961);
A interpretação dinâmica destas formações é muito diferente entre os vários autores. Nalguns territórios ibéricos (Catalunha e SE do Sistema Ibérico) definiram-se associações novas de teixedos (Rodríguez Guitián et al., 2011), considerando-se determinante o lento crescimento do teixo e a sua capacidade para sobreviver em condições de sombra total. Porém, no NW da Península, optou-se por não os diferenciar dos bosques envolventes (Rodríguez Guitián et al., 2011; Romero Rodríguez, 1993). No entanto, analisando os inventários de teixedos disponíveis para o NW Ibérico, agora estudados, os elementos arbóreos, que resultam dos contactos catenais, são raros ou subjugados à dominância do teixo e, no geral, estão ausentes dos núcleos mais densos. A grande exceção é Betula pubescens, que em três inventários apresenta uma cobertura entre 25 e 75%, e pontualmente Acer pseudoplatanus e Fraxinus excelsior (anexo I);
Sendo Taxus baccata uma das árvores de maior longevidade da flora europeia eurossiberiana, podendo atingir 1500 a 3000(5000) anos (Benham et al., 2016; Cortés et al., 2000), não parece fazer sentido que os seus bosques sejam apenas faciações de outros, que poderão até comportar-se como formações secundárias dos primeiros, tais como de Fagus sylvatica [150-300(400) anos; Houston Durrant et al., 2016] ou Betula pubescens nos teixedos lusos [(80)100-150 anos; Beck et al., 2016; Cortés et al., 2000]. Já Quercus robur é uma espécie mais longeva [>1000 anos (Eaton et al., 2016)], mas nas áreas estudadas em Portugal não se observaram teixedos em mosaico com os respetivos carvalhais, embora seja admissível que possam ocorrer pontualmente, sobretudo em áreas pedregosas e expostas a N.;
A posição catenal das comunidades de teixo é igualmente potenciadora de interpretações díspares. Cortés et al. (2000) e Díaz González e Penas (2017) referem que, na Galiza, Cordilheira e Cornija Cantábrica, os teixos surgem provavelmente mais difundidos nos faiais e bidoais do andar montano (supratemperado inferior) e sobretudo nas suas orlas. Nas serras lusitanas e noutros territórios de Espanha (e.g. Sierra de Francia, Salamanca) esta espécie, de ótimo atlântico, ocorre em corgas e barrocas parecendo beneficiar da compensação hídrica por via edáfica (Fernández-González et al., 2015). Ainda que o teixo não seja um táxon ripícola s.str., suporta a inundação e o escoamento superficial intenso devido ao seu sistema radicular, nomeadamente se tiver na maior parte do ano boas condições de drenagem (Benham et al., 2016). Encontram-se padrões semelhantes noutros táxones considerados relictos, tais como Prunus lusitanica (Calleja Alarcón, 2006), ou Ilex aquifolium, (Costa et al., 2012; Rodríguez Guitián et al., 2011), com preferência por áreas compensadas hidricamente, com elevada pedregosidade e rochas aflorantes, tais como vales relativamente pequenos, mas com elevado encaixe, o que reduz a exposição solar e promove a ocorrência de microclimas com consideráveis níveis de ensombramento e humidade atmosférica;
Por último, o conceito fitossociológico de fácies (Capelo, 2003) tem dissuadido o reconhecimento dos bosquetes de teixo como comunidades autónomas, pela inexistência de táxones com ótimo ecológico nestas comunidades, à exceção do próprio teixo.
2. Classificação sintaxonómica
Com base nos inventários fitossociológicos do anexo I e nos resultados da análise DiffVal1 efetuada julga-se consequente o reconhecimento dos seguintes sintáxones:
Para o teixedo geresiano propõe-se o nome Eryngio juresiani-Taxetum baccatae Portela-Pereira, Monteiro-Henriques, Casas, Forner e Neto ass. nov. (holotypus: inventário nº 2, anexo I);
Para o teixedo estrelense, apesar do contexto florestal fortemente fragmentado e alterado do andar supratemperado (Jansen & Capelo, 2013; Meireles, 2010), propõe-se a subassociação Eryngio juresiani-Taxetum baccatae loniceretosum hispanicae Portela-Pereira, Monteiro-Henriques, Forner e Neto subass. nov. (holotypus: inventário nº 13, anexo I). Deste modo, o teixedo geresiano constitui a subassociação typicum;
Para o teixedo do W cantábrico propõe-se o nome Corylo avellanae-Taxetum baccatae Portela-Pereira, Monteiro-Henriques e Neto ass. nov. (holotypus: columna 6, tabla 2 inRodríguez Guitián et al., 2011).
O enquadramento sintaxonómico dos teixedos é, como se referiu, complexo, dada a posição ecológica em que ocorrem. O elenco florístico presente nestas comunidades contém táxones característicos de diferentes classes de vegetação mesófila, nomeadamente da Quercetea robori-petraeae (sobretudo nas serras da Estrela e do Gerês), da Carpino-Fagetea sylvaticae e mesmo da Quercetea pubescentis (no W cantábrico), bem como da classe de vegetação higrófila Alno glutinosae-Populetea albae (nas três áreas estudadas). Ainda assim, parecem predominar os táxones mesófilos em relação aos higrófilos. Pitarch García (2002) e Crespo et al. (2008) consideraram a associação Ononido aragonensis-Tilietum platyphylli, pontualmente codominada por Taxus baccata, como a ocorrência mais meridional da aliança Tilio-Acerion, aliança que foi recentemente estendida ao território português (Monteiro-Henriques et al., 2010) e que representa vegetação arbórea de barrocas e de canhões muito encaixados.
O posicionamento dos bosquetes de teixo do NW da Península Ibérica poderá merecer uma aliança geovicária (para a região eurossiberiana) da aliança Lathyro veneti-Taxion baccatae (Quercetea pubescentis, ver Mucina et al., 2016), que inclui bosques relictos mistos de carvalhos caducifólios e teixo das ilhas de Córsega e Sardenha. Porém, dada a sua ecologia fortemente relacionada com os declives acentuados das margens de pequenos ribeiros propõe-se incluir provisionalmente (até um estudo mais aprofundado a nível europeu e mediterrânico) na aliança Tilio-Acerion.
O quadro I apresenta o esquema sintaxonómico com o posicionamento das novas associações, seguindo Mucina et al. (2016).

Defende-se aqui que, futuramente, deverão ser analisadas as comunidades relictas dominadas por Taxus baccata num contexto europeu alargado, uma vez que não é de descartar a possibilidade de se incluírem estas comunidades numa aliança comum.
3. Os Teixedos na paisagem orófila lusitana
A classificação dos teixedos aqui alcançada é mais um contributo para o conhecimento dos bosques orófilos lusitanos, cuja sintaxonomia não se encontra resolvida ou, nalguns casos, sequer estabelecida (Portela-Pereira et al., 2016). Os habitat colonizados pelos teixedos lusitanos, assim como os seus elencos florísticos, sugerem claramente uma relação sucessional entre estes e os bosques de Betula pubescens supratemperados (Betulion fontquerio-celtibericae; Mucina et al., 2016) considerados bosques secundários ou potencialmente climácicos e mesófilos (altimontanos) nas serras do Gerês e da Estrela (Honrado, 2003; Meireles, 2010). Disso é exemplo o facto de uma boa parte da paisagem florestal atual no Gerês supratemperado (inferior) ser constituída por bidoais, em condições mais ou menos higrófilas, que no inverno deixam ver inúmeros exemplares ou núcleos de Taxus baccata no seu interior.
Nos estudos decorrentes do projeto LIFE TAXUS discutiram-se estas e outras questões da relação/contactos entre teixedos, bidoais e outras séries de vegetação geresianas e estrelenses, e até que ponto estes poderão ser mesófilos em serras altimontanas onde o solo florestal está limitado a valeiros e covões, numa paisagem convexa dominada por granito exposto (Portela-Pereira et al., 2016). Tal discussão baseia-se na valência ecológica do teixo e no reconhecimento, por parte de vários autores, da possibilidade de um andar de vegetação com codominância de teixo nas áreas cuminais da Serra da Estrela (Catarino, 2007; Jansen, 2011; Pinto da Silva & Teles, 1999) ou na ilha da Madeira (Capelo et al., 2007), e, consequentemente, de outras serras ibéricas, como o Gerês; assim como na evidência histórica de um teixedo ter ocupado (parte) da laurissilva basal açoriana num passado recente (Dias et al., 2007). No entanto, na prática, com o aumento da mediterraneidade do nosso território e dada a elevada frequência de fogos, as serras portuguesas, muito provavelmente, nunca alcançarão um equilíbrio natural suficiente para ter um andar dominado por teixedos. Dada a distribuição atual muito restrita de Taxus baccata, os impactos potenciais das mudanças climáticas projetadas poderão ser especialmente negativos, não só devido à redução do habitat disponível (Draper & Marques, 2007), como pela reduzida dimensão dos bosquetes atuais e pela depressão de propágulos (Aguiar & Pinto, 2007).
Em suma, em termos sucessionais, os teixedos lusitanos constituem-se (sobretudo) como séries climácicas, relictas, fluvio-rivulares ou rivular-coluviais, tempori-higrófilas a mesófilas de montanha (meso)supratemperadas ou suprassubmediterrânicas, frequentemente substituída por um bidoal nas áreas mais expostas a perturbações.
V. Conclusão
A análise da matriz dos teixedos do NW ibérico demonstrou claramente três grupos de teixedos com significado biogeográfico e com táxones diferenciais. A diferença entre os teixedos W cantábricos e os lusitanos é deveras evidente, já que se destacam nos primeiros inúmeros táxones que não fazem parte da flora lusitana. A diferenciação entre os teixedos geresianos e estrelenses é um pouco mais complexa (dado o estado atual do teixedo estrelense), mas faz-se, de forma semelhante, por táxones diferenciais da vegetação geresiana que não alcançam a Serra da Estrela. Correspondem a endemismos do noroeste ibérico ou outros elementos que, naturalmente, não atingem esta serra, onde os teixedos têm um dos seus limites meridionais de distribuição global. A reduzida presença de espécies diferenciais com valor biogeográfico leva-nos a fazer essa diferenciação ao nível de subassociação.
Com este trabalho pretendeu-se facilitar a interpretação fitossociológica do Habitat 9580* da Rede Natura 2000, nomeadamente no NW peninsular, onde se classificam três teixedos na aliança Tilio-Acerion da classe Carpino-Fagetea sylvaticae (Mucina et al., 2016).
Sendo estas comunidades raras e (co)dominadas por um táxon relicto em regressão na Península Ibérica, é crucial a interpretação florística e aprofundamento das suas características ecológicas (sucessionais e catenais) que aqui se apresentaram. Até porque, nomeadamente na Serra da Estrela, o teixedo (Habitat 9580*) encontra-se extremamente ameaçado e reduzido. Tentando reverter essa situação e a ameaça constante dos incêndios florestais, o projeto LIFE TAXUS (QUERCUS, 2016), através de plantações e controlo de matos, procurou restaurar esse Habitat e fomentar novas áreas na Serra da Estrela, assim como salvaguardar os teixedos da Serra do Gerês.
Agradecimentos
Este trabalho decorre do Projecto LIFE TAXUS, LIFE12 NAT/PT/000950 Taxus - Restoring Yew Thickets [9580* Mediterranean Taxus baccata Woods].
Agradecimentos especiais a António Rebelo do Parque Nacional Peneda-Gerês e Rafael Neiva do Parque Natural da Serra da Estrela pela ajuda e partilha no campo.
No âmbito da sua participação neste trabalho, T. M. H. foi financiado pelo Fundo Social Europeu (POCH) e por Fundos Nacionais (MCTES), através de uma bolsa de pós-doutoramento da Fundação para a Ciência e a Tecnologia (SFRH/BPD/115057/2016). Neste trabalho foi financiado por Fundos Nacionais através da FCT - Fundação para a Ciência e a Tecnologia, no âmbito do projeto UIDB/04033/2020.
Contributos dos/as autores/as
Estêvão Portela-Pereira: Conceptualização; Metodologia; Validação; Análise formal; Investigação; Curadoria dos dados; Escrita - preparação do esboço original; Redação - revisão e edição. Tiago Monteiro-Henriques: Conceptualização; Metodologia; Validação; Análise formal; Investigação. Carme Casas: Validação; Redação - revisão e edição. Nuno Forner: Validação; Redação - revisão e edição; Administração do projeto; Aquisição de financiamento. Isabel Garcia-Cabral: Validação; Redação - revisão e edição; Administração do projeto; Aquisição de financiamento. João Paulo Fonseca: Validação; Redação - revisão e edição. Carlos Neto: Conceptualização; Metodologia; Validação; Redação - revisão e edição; Supervisão.