










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias v.32 n.2 Lisboa dez. 2009
Avaliação da nutrição de plantações jovens de eucalipto por análise foliar e métodos não destrutivos
Nutrition evaluation in young Eucalyptus plantation by foliar analysis and non-destructive methods
Ana Carla Madeira1, Manuel Madeira, António Fabião, Paulo Marques, Marta Carneiro
RESUMO
Avaliou-se o crescimento e estado nutritivo de jovens Eucalyptus por análise foliar tradicional (teores de N, P e pigmentos fotossintéticos) e com o medidor de clorofila SPAD-502. Efectuou-se uma gestão diferenciada dos resíduos de abate da plantação anterior, com incorporação dos resíduos no solo (I), remoção dos resíduos (R), e distribuição dos resíduos à superfície (S); aplicou-se um fertilizante azotado (IF, RF e SF) e introduziuse uma leguminosa do género Lupinus (IL e RL). Não houve diferenças significativas no crescimento entre I, R e S. A aplicação de fertilizante intensificou o crescimento em SF e IF. O teor foliar de N foi no início positivamente afectado pelo Lupinus (IL e RL). Após a primeira aplicação de fertilizante, este teor foi significativamente superior em IF, RF e SF, esbatendo-se a diferença entre tratamentos após as aplicações seguintes; semelhante padrão foi observado para os pigmentos e o SPAD. Os valores de SPAD correlacionaram-se (P<0,05) com N foliar. O sistema de gestão mais produtivo incluiu a incorporação dos resíduos no solo com aplicação repetida de fertilizante azotado.
Palavras-chave: Crescimento de Eucalyptus, leguminosas, nutrição, solo, SPAD
ABSTRACT
The growth and nutritional state of young Eucalyptus was evaluated by foliar analysis (contents of N, P and photosynthetic pigments) and the SPAD-502 chlorophyll meter. An experimental system was used with different harvest residues management, such as incorporation of residues into the soil (I), removal of residues (R), and distribution of residues on the soil surface (S), with N fertiliser application (IF, RF and SF), and with the leguminous Lupinus seeding (IL and RL). No significant differences in growth were found between the I, R and S. Application of fertiliser increased growth in the SF and IF. Initially, foliar N content was positively affected by the leguminous (RL and IL). After the first fertiliser application, a higher N content was observed in the IF, RF and SF, the differences being reduced in the following applications; a similar trend was observed for extracted pigments and SPAD values. Foliar N contents were correlated (P<0.05) with SPAD values. The management system more efficient regarding tree productivity included incorporation of residues into soil with repeated applications of N fertiliser.
Key-words: Eucalyptus growth, leguminous, nutrition status, soil, SPAD
INTRODUÇÃO
A gestão apropriada dos resíduos de abate e dos nutrientes é considerada crucial para aumentar a produtividade e garantir a sustentabilidade de plantações florestais (Powers et al., 1990; Smethurst & Nambiar, 1990; Proe & Dutch, 1994). Neste contexto, assume particular relevância a gestão das plantações de eucaliptos (Eucalyptus globulus) exploradas intensivamente em regime de talhadia (Adams & Attiwill, 1986; Jones et al., 1999; Shammas et al., 2003), dada a elevada quantidade de nutrientes que se acumula nos respectivos resíduos de abate e nas camadas orgânicas (Madeira et al., 1995; Spangenberg et al., 1996; Jones et al., 1999). A remoção ou manutenção (à superfície ou nas camadas superficiais do solo) dos resíduos de abate é polémica devido não só à relação entre a gestão dos resíduos e a qualidade do solo, mas também à necessidade urgente de novas formas de energia com vista à substituição dos combustíveis fósseis (Cowie et al., 2006; Stupak et al., 2007).
Para além da gestão dos resíduos de abate das plantações florestais, outros sistemas devem ser testados com o fito de melhorar a fertilidade e qualidade do solo e o crescimento das árvores (Fisher & Binkley, 2000). Em Portugal, não têm sido observadas diferenças significativas entre as várias opções de gestão dos resíduos de abate no respeitante ao crescimento e estado de nutrição das árvores e às características do solo em plantações de eucalipto, tanto no caso de replantação como no de segunda rotação (Magalhães, 2000; Soares et al., 2002). O efeito da aplicação de fertilizantes (na ausência de resíduos de abate) tem-se mostrado positivo (Pereira et al., 1996), mas as diferenças não foram significativas. Porém, a interacção entre os sistemas de gestão dos resíduos e a aplicação de fertilizante não é conhecida. Dados os desafios de competitividade que o sector florestal enfrenta, é importante compreender, nas condições Mediterrânicas, o efeito conjugado de diferentes sistemas de gestão de resíduos de abate e o uso de fertilizantes azotados ou de plantas fixadoras de azoto no incremento da produtividade das plantações de eucaliptos.
A utilização de técnicas não destrutivas para análise foliar (medidor portátil de clorofila SPAD-502) tem permitido estimar o teor relativo de clorofila com precisão. Contudo, a maioria dos estudos referidos na literatura apenas relaciona a clorofila total com os valores de SPAD, não se conhecendo a correlação com os carotenóides. Tem-se observado forte correlação entre os valores de SPAD e o teor de azoto foliar, por exemplo, em laranjeiras (Pestana et al., 2004), espécies folhosas (Chang & Robison, 2003), plantações jovens de E. globulus e E. nitens (Pinkard et al., 2006) e pés-mãe de E. globulus (Ribeiro et al., 2009). Por esta razão, o SPAD pode ser uma metodologia alternativa e promissora relativamente à tradicional e morosa análise foliar.
Neste contexto, instalou-se um sistema experimental numa área representativa de plantações de eucaliptos, no centro de Portugal, com os objectivos de (a) avaliar a influência da gestão dos resíduos de abate (remoção, distribuição à superfície ou incorporação no solo) e o uso de fertilizante azotado e associação com leguminosa no crescimento e nutrição das jovens árvores, de forma a identificar o sistema de gestão mais apropriado para optimizar a produtividade das plantações; (b) comparar o desempenho de análises foliares destrutivas e não destrutivas para determinar o estado nutricional das árvores.
MATERIAL E MÉTODOS
Sistema e desenho experimental
O sistema experimental foi instalado na Quinta do Furadouro (Óbidos; lat. 39º 20 N, long. 09º 14 W, alt. 50 m), numa área de replantação de E. globulus, após o abate (Outono de 2001) da plantação anterior com 34 anos de idade. A área útil das parcelas era de 225 m2, correspondendo a 25 árvores que foram objecto de estudo.
A área de estudo assenta em formações do Cretácico (arenitos) (Grés de Torres Vedras; Zbyzewski & Almeida, 1960), e o relevo apresenta declives entre 5 e 12%. Os solos são maioritariamente Cambissolos dístricos e Regossolos dístricos (WRB, 2006), apresentando textura franco arenosa a franca, valores de pH (H2O) < 5,3, teores baixos de C orgânico (10,9 – 24,3 g kg-1) e muito baixos de P extraível (< 5 mg kg-1) (Jones et al., 1999). A temperatura média anual é de 15,2ºC, enquanto que a média mensal varia de 10,4 a 19,8 ºC, em Janeiro e Agosto, respectivamente. A precipitação média anual é de 607 mm, mas apenas 10% ocorre no período compreendido entre Maio e Setembro (Reis & Gonçalves, 1981; estação climatológica das Caldas da Rainha, 39º 24 N, 09º 08 W, alt. 70 m).
Os tratamentos aplicados (repetidos em quatro blocos) incluíram: incorporação dos resíduos de abate no solo por intermédio de gradagem a uma profundidade média de 20 cm (I); I com aplicação de fertilizantes no final do Inverno (Março) nos três anos seguintes ao da plantação (IF); I com sementeira de leguminosas (Lupinus luteus) (IL); remoção manual da totalidade dos resíduos de abate (R); R com aplicação de fertilizantes (RF); R com sementeira de leguminosas (RL); manutenção e distribuição uniforme dos resíduos de abate na superfície do solo (S); S com aplicação de fertilizantes (SF). A leguminosa foi semeada por uma escarificação ligeira, aplicando-se 110 kg de semente e 220 kg de superfosfato a 18% por hectare. A rebentação das touças foi controlada pela aplicação de glifosato (Roundup).
Após a aplicação dos tratamentos, a plantação (ao covacho) realizou-se, entre as linhas de touças da plantação anterior e a um compasso de 3 x 3 m, no final de Março de 2002. À plantação aplicou-se 150 g de adubo composto ternário (12% N, 24% P2O5 e 12% K2O), metade de cada lado da planta e a cerca de 20-25 cm desta. Em Março de 2003, 2004 e 2005 aplicou-se manualmente 200 kg ha-1 de um fertilizante azotado (ENTEC 26, 26% N, do qual 7,5% de N-NO3 e 18,5% de N-NH4+ e 32,5% de anidrido sulfúrico, SO3 solúvel em água) nas parcelas dos tratamentos IF, RF e SF.
Medições e amostragens
Antes da instalação do sistema experimental quantificou-se a massa dos resíduos de abate que existiam nas parcelas delimitadas, a que se seguiu a caracterização dos respectivos solos por intermédio da abertura e descrição de seis perfis. Os resíduos de abate (total de 38 t ha-1) incluíam folhas (11 t ha-1), ramos (18 t ha-1), raminhos e casca (10 t ha-1) (Madeira et al., 2009). Os resíduos continham cerca de 165 kg ha-1de N e 15 kg ha-1 de P, ocorrendo principalmente nas folhas (66 e 56% de N e P, respectivamente), e cerca de 443 kg ha-1 de Ca distribuídos por folhas e ramos. Na camada superficial do solo (0-10 cm), os teores de C orgânico e N eram baixos (13,6 e 0,53 g kg-1, respectivamente), a razão C/N alta (25,9), e a disponibilidade das bases de troca razoável (2,96, 0,66, 0,27 e 0,08 cmolc kg-1, respectivamente, para Ca, Mg, K e Na).
A mortalidade das jovens árvores foi determinada em Outubro de 2002 e Setembro de 2003. A altura e o diâmetro à altura do peito (DAP) foram medidos para avaliar a evolução do crescimento das árvores. A altura das árvores foi medida em Outubro de 2002, Setembro de 2003, Agosto de 2004, Setembro de 2005, Agosto de 2006 e Novembro de 2007 (7, 18, 29, 42, 53 e 68 meses após a plantação, respectivamente) e o DAP foi medido nas quatro últimas amostragens. O volume das árvores (m3 ha-1) foi calculado segundo a equação
V = 0,00003739 D1,8151 H1,1455,
onde D é o diâmetro à altura do peito e H a altura da árvore (Tomé et al., 2007).
A componente aérea da vegetação sob coberto (expressa em biomassa, g m-2) foi estimada no final das Primaveras de 2002 a 2006 e para tal usou-se uma estrutura de madeira (0.5×0.5 m2) quatro vezes em cada parcela do respectivo tratamento.
Para a medição da área foliar (Portable Area Meter Model Li-3000A, LI-COR), teor foliar de azoto (método de Kjeldahl, sistema de destilação Kjeltec Auto 1030 Analyser), de fósforo (CEM Microwave Digestion System Model MDS-81 D e colorimetria pelo método do ácido ascórbico), e teor relativo de clorofila (SPAD-502, Minolta Corporation, Ramsey, NJ) foram feitas sete amostragens (cinco para o SPAD), tendo em duas árvores por tratamento e por bloco sido colhidas oito folhas completamente expandidas em quatro ramos do terço superior da copa, segundo as quatro direcções dos pontos cardeais.
Os teores de clorofila e de carotenóides totais foram obtidos por extracção em acetona a 100% e ascorbato de sódio, a partir de discos (0,7 cm diâmetro) obtidos de oito folhas por tratamento. Os teores (µg cm-2) foram então calculados de acordo com as equações de Lichtenthaler (1987). Nas áreas correspondentes aos discos usados para extracção dos pigmentos foram feitas leituras com o SPAD para posterior correlação e calibração.
Análise estatística
O efeito dos tratamentos no crescimento e estado de nutrição das árvores foram testa-dos por análise de variância (ANOVA) e a comparação das médias dos tratamentos pelo Tukey multiple range test. Os procedimentos estatísticos foram efectuados através do Statistica 6.0 software package (StatSoft, 1996). Para discriminar a importância relativa dos factores que foram considerados (gestão de resíduos, leguminosas e aplicação de fertilizante azotado), foi aplicada uma análise de variância de três factores; para este procedimento estatístico usou-se o Statgraphics plus 5.1 (Manugistics Inc.).
RESULTADOS E DISCUSSÃO
Vegetação sob coberto
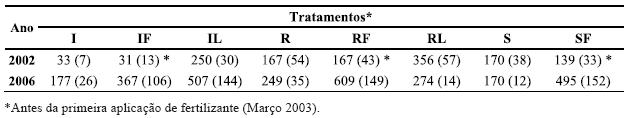
A biomassa da componente aérea da vegetação sob coberto mostrou grande variabilidade entre tratamentos durante todo o período de amostragem. No fim da primeira Primavera após a plantação (2002, Quadros 1 e 2), e antes da primeira fertilização, a biomassa foi significativamente superior (P<0,001) nos tratamentos com leguminosa (RL e IL) do que nos outros; a biomassa da vegetação sob coberto revelou também ser superior (P<0,05) nos tratamentos com resíduos relativamente aos sem resíduos (Quadros 1 e 2). Nas amostragens efectuadas em 2003, 2004 e 2005, as diferenças entre os tratamentos não foram significativas, quer para o efeito das leguminosas, quer para os dos resíduos e da fertilização. Porém, a última amostragem, realizada em 2006 (Quadro 1), mostrou que apenas a fertilização aumentou significativamente a biomassa da vegetação sob coberto (P<0,005; Quadro 2), não tendo a gestão dos resíduos ou a instalação de Lupinus tido qualquer efeito significativo (P>0,05).
Quadro 1 – Biomassa (g m-2) da vegetação sob coberto determinada no final da Primavera de 2002 e 2006.
Quadro 2 – Resultados da ANOVA multifactorial relativamente aos efeitos dos resíduos de abate (sem e com resíduos), da instalação de Lupinus (com e sem leguminosa) e da aplicação de fertilizante azotado (com e sem fertilização) na biomassa da vegetação sob coberto (BVC) e na altura, diâmetro à altura do peito e volume das árvores.
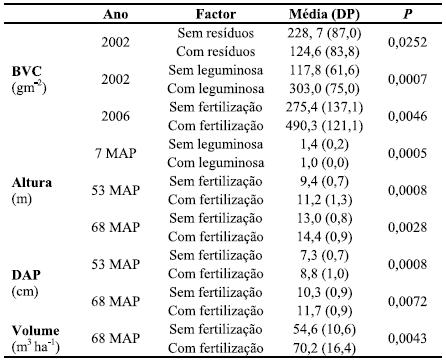
O efeito positivo das leguminosas na acumulação de biomassa, na fase inicial da rotação, sugere claramente que estas espécies poderão ter uma efeito bastante positivo nos inputs de N no sistema, o que poderá ter repercussões no estado de nutrição das árvores e também na acumulação de carbono orgânico no solo.
A acção positiva da fertilização azotada na biomassa da vegetação sob coberto, um ano após a última aplicação, sugere também um efeito positivo na ciclagem de nutrientes e na acumulação de carbono orgânico no solo. Com efeito, a massa da vegetação sob coberto, bem como a quantidade de nutrientes nela acumulados, são maiores do que os correspondentes à folhada anual das plantações de eucalipto na área de estudo (Madeira et al., 1995). Assim, a vegetação sob coberto poderá contribuir indirectamente para dilatar o efeito temporal da fertilização azotada, contribuindo para o aumento da fertilidade do solo e do respectivo stock de C. Porém, é imprescindível obter informação pertinente sobre o efeito da fertilização na biomassa da parte subterrânea da vegetação sob coberto para melhor avaliar tais tendências.
Crescimento das árvores
A mortalidade das árvores, avaliada 7 e 18 meses após a plantação (MAP), foi baixa (7 a 12%) e sem diferenças significativas entre tratamentos.
A incorporação de resíduos de abate (e também das camadas orgânicas) no solo (I) parece melhorar o crescimento das jovens árvores, relativamente à sua remoção (R) ou distribuição na superfície (S). Contudo, as diferenças entre tratamentos diminuíram com o tempo. No fim do estudo (68 MAP, Figura 1), apesar das árvores do tratamento I serem mais altas (13,6 m) e mais grossas (10,7 cm) do que as dos tratamentos S (12,2 m e 9,1 cm, respectivamente) e R (12,1 m e 9,0 cm, respectivamente), não se observaram diferenças significativas (Figura 1). Com efeito, no início do estudo, 7 MAP, a altura das árvores do tratamento I era cerca de 1,3 vezes maior do que nas de S e R, comparativamente a 1,16 e 1,12 a 68 MAP. Esta tendência, também observada para o DAP, sugere que o efeito positivo do tratamento I no crescimento das árvores se esbateu em fase mais avançada da rotação. Assim, não obstante o menor volume ter sido medido nos tratamentos S e R (41,3 e 45,2 m3 ha-1) (68 MAP; Figura 1), este não foi significativamente inferior ao observado no tratamento I.
Figura 1 – A. altura das árvores medida 7 meses após a plantação (MAP); B. altura e diâmetro à altura do peito (DAP) medidos 53 MAP; C. altura e DAP medidos 68 MAP (no fim do período experimental); D. volume do tronco (m3 ha-1) das árvores medidos 68 MAP. As barras representam o desvio padrão.
Os resultados do presente estudo contradizem os obtidos por Soares et al. (2002) num ensaio lisimétrico em condições controladas e na ausência de cobertura vegetal do solo, onde as árvores de I mostraram um crescimento inicial mais lento do que em S e semelhante a R. No presente estudo, a perturbação causada pelo operação de gradagem na incorporação dos resíduos pode ter diminuído a massa volúmica e a compactação do solo, como já observado anteriormente por Madeira et al. (1989), favorecendo condições para uma mais rápida e profunda expansão do sistema radicular e, portanto, um melhor uso da água e dos nutrientes das camadas subsuperficiais. Em conformidade com Fabião et al. (1990), o sistema radicular do E. globulus tem uma expansão rápida, atingindo camadas profundas de solo, durante a primeira estação de crescimento, o que parece explicar as diferenças de crescimento observadas no tratamento I. A menor biomassa de vegetação espontânea nas parcelas deste tratamento (Quadro 1) pode também ter implicado me-nor restrição no crescimento das árvores relativamente a R e S.
No presente estudo, a manutenção dos resíduos na superfície do solo (S), apesar da elevada quantidade de nutrientes neles acumulada, não aumentou o crescimento das árvores quando comparado com a sua remoção (R), o que corrobora os resultados de Jones et al. (1999) para o crescimento de plantações jovens de eucaliptos em solos com características semelhantes (nas imediações da área do presente estudo) e na Galiza (Espanha). Porém, os nossos resultados são contrários aos relatados por Nzilla et al. (2002) no Congo, Merino et al. (2003) na Galiza (Espanha) e Xu et al. (2004) na China, onde a manutenção dos resíduos melhorou o crescimento e as condições nutricionais das árvores. Esta discordância pode ser atribuída às condições mediterrânicas, em que a produtividade está principalmente limitada pela disponibilidade de água (Pereira et al., 1989; Pereira et al., 1996), bem como a maiores disponibilidades e reserva de nutrientes no solo do presente estudo.
No início da rotação, a sementeira de Lupinus levou a um menor crescimento em altura das árvores comparativamente com os tratamentos sem leguminosa (P<0,001; Quadro 2); os resíduos de abate (presença ou ausência) não afectaram significativamente a altura (P>0,05). A leguminosa L. luteus afectou mais negativamente as árvores em solos com a incorporação de resíduos de abate, sendo a sua altura em IL (1,0 m) significativamente mais baixa do que em I (1,7 m) a 7 MAP (Figura 1). Esta diminuição de crescimento corrobora resultados de Malik et al. (2001) e Mendham et al. (2004), os quais sugerem es-tar relacionado com a competição pela água entre as árvores e a vegetação espontânea, ou leguminosas, mormente se o sistema radicular das jovens árvores não tenha colonizado as camadas mais profundas de solo. Contudo, o efeito negativo das leguminosas na fase inicial da rotação modificou-se fortemente a meio do período da mesma. De facto, 68 MAP (Figura 1) a altura das árvores em IL era muito próxima da observada em I (13,2 e 13,6 m, respectivamente). Pelo contrário, a altura das árvores em RL era maior do que em R (13,8 e 12,1 m, respectivamente; Figura 1), ao invés do padrão observado 7 MAP (Figura 1). Este comportamento parece estar relacionado com o balanço entre a remoção de N contido nos resíduos de abate e a adição de N pela leguminosa.
Tal como para a biomassa da vegetação sob coberto, a fertilização azotada teve um efeito positivo e significativo nos tratamentos onde foi efectuada relativamente aos que não tiveram aplicação, quer para a altura, quer para o DAP (P<0,005 e P<0,01, respectivamente; Quadro 2). A aplicação do fertilizante em Março de 2003 e 2004 aumentou a altura das árvores nos tratamentos IF (8,0 m), SF (7,1 m) e RF (6,0 m) relativamente aos outros, sendo as diferenças significativas entre SF e S, e entre IF e IL (29 MAP; Quadro 3). A altura das árvores foi 1,3, 1,5 e 1,2 vezes maior nestes tratamentos, respectivamente, do que nos respectivos não fertilizados I, S e R. Similarmente, o DAP foi 1,3 (IF e RF) e 1,5 (SF) vezes maior, respectivamente. No termo do estudo (68 MAP; Figura 1), as árvores foram 1,1 (IF e RF) e 1,2 (SF) vezes mais altas e mais grossas do que nos tratamentos homólogos não fertilizados, sendo o tratamento IF significativamente diferente dos tratamentos R e S. O efeito da remoção dos resíduos de abate parece ter sido compensado pelo fertilizante, dado que o crescimento das árvores em RF foi semelhante ao observado nos tratamentos I e S. O volume foi maior nos tratamentos fertilizados (P<0,005; Quadro 2) relativamente aos não fertilizados e não foi significativamente afectado pela manutenção dos resíduos de abate ou da leguminosa (P>0,05).
Quadro 3 – Evolução dos teores de N foliar (mg g-1), clorofila total (CLT) e carotenóides totais (CART) expressos em µg cm-2, e valores de SPAD para todos os tratamentos durante o período experimental; os valores representam a média e o desvio padrão (entre parêntesis).
A falta de resposta significativa, em termos nutricionais, à gestão dos resíduos de abate é também suportada pelo facto de que o crescimento das árvores aumentou (31, 18 e 40% nos tratamentos I, R e S, respectivamente) pela repetida fertilização azotada tanto na ausência como na presença de resíduos, sugerindo que as taxas de mineralização de N foram insuficientes para que um crescimento optimizado ocorresse. O crescimento destas árvores em resposta à fertilização corrobora os resultados obtidos em eucaliptais jovens, onde a adição de fertilizante em solo arenoso sem resíduos (de abate ou orgânicos) aumentou a produtividade em 19% (Pereira et al., 1989; Pereira et al., 1996).
A manutenção dos resíduos de abate no solo pode ter também efeitos de sinergia com a fertilização, dado que o aumento no crescimento foi superior em I (31%) e S (40%) do que em R (18%). Assim, em solos com baixa fertilidade, independentemente do sistema de gestão dos resíduos de abate, haverá a necessidade de aumentar o fornecimento de N ao sistema através de fertilização para que se atinja a taxa de crescimento optimizada.
Análise foliar e SPAD-502
Antes da primeira aplicação de fertilizante azotado (8 MAP), o teor foliar de N foi maior em IL (26,7 mg g-1) e RL (25,5 mg g-1) do que nos outros tratamentos (21,0-21,8 mg g-1) (Quadro 3), o que se atribui ao aumento de inputs de N devido às leguminosas; a similitude entre os tratamentos I, S e R corrobora os resultados relatados por Jones et al. (1999), para tratamentos semelhantes localizados próximo da área do presente estudo.
Dois meses após a primeira fertilização (14 MAP), o N foliar em IF, RF e SF foi estatisticamente superior (22,4-23,6 mg g-1) ao obtido nos outros tratamentos (14,7-16,5 mg g-1). No entanto, o teor de N diminuiu relativamente às amostragens imediatamente anterior e posterior, o que pode estar relacionado com uma infecção por Micosphaerella spp. que afectou as folhas das árvores. Não obstante o fertilizante ter suplementado apenas N, teores de P mais elevados foram também observados em IF, RF e SF (1,7, 1,5 e 1,5 mg g-1, respectivamente; Quadro 3), mas somente IF foi estatisticamente diferente de R, RL e S (1,3 mg g-1). A razão N/P foi também estatisticamente superior nos tratamentos fertilizados (15,2) do que nos não fertilizados (11,7).
Oito meses após a primeira fertilização (20 MAP), a diferença verificada entre os tratamentos fertilizados (26,9-28,0 mg g-1) e os não fertilizados (25,1-26,0 mg g-1) não foi tão pronunciada como a verificada 14 MAP. A segunda fertilização azotada (Março 2004) não afectou igualmente os tratamentos 26 MAP: o teor de N nas folhas foi de 28,3, 23,0 e 21,2 mg g-1, em IF, RF e SF, respectivamente, sendo estatisticamente diferentes apenas os valores correspondentes a IF e SF. Um teor mais elevado de P foi medido em IF (1,8 mg g-1) nesta amostragem, sugerindo um efeito conjugado positivo entre a fertilização azotada e a incorporação de resíduos no solo. A terceira aplicação de fertilizante (Março 2005) não afectou o N foliar em IF, RF e SF. Pelo contrário, os valores mais altos de N, 44 e 50 MAP, foram medidos no tratamento IL, o que sugere o efeito combinado a médio prazo da incorporação de resíduos e da leguminosa L. luteus.
O teor de pigmentos fotossintéticos e os valores de SPAD foram estatisticamente semelhantes a 8, 20 e 32 MAP (Quadro 3). Contudo, 14 MAP foram observadas diferenças significativas entre os tratamentos fertilizados e os não fertilizados, sendo esta diferença relacionada com o maior teor de N foliar nos tratamentos fertilizados. Embora não tenham sido observadas grandes diferenças nos valores de SPAD entre tratamentos a 26 MAP, a clorofila total em SF (63,1 µg cm-2) foi estatisticamente superior à extraída das árvores dos tratamentos IL, RL e R (45,4, 47,2 e 48,1 µg cm-2, respectivamente; Quadro 3). Também, o teor de carotenóides mais elevado ocorreu em SF (13,6 µg cm-2) relativamente a IL e RL (cerca de 10,0 µg cm-2). A partir de 32 MAP, não se observaram diferenças significativas entre tratamentos para os teores de pigmentos fotossintéticos e valores de SPAD.
Para além da lacuna de informação sobre a utilização do SPAD como medidor de carotenóides, ela também é escassa relativamente à clorofila a e b, excepto para plantas hortícolas (Madeira et al., 2000, 2005) e uma espécie florestal - a bétula (Richardson et al., 2002). Assim, neste estudo analisaram-se as correlações entre os referidos pigmentos e os valores de SPAD, sendo os coeficientes de determinação (R2) destas correlações muito elevados a 14 e 20 MAP (Quadro 4). A forte correlação com os carotenóides totais sugere que o SPAD-502 pode ser também muito útil na determinação do teor relativo deste pigmento.
Quadro 4 – Equações de regressão para a relação entre os valores de SPAD (x) e os pigmentos fotossintéticos (y, µg cm-2) a 14, 20 e 26 MAP.
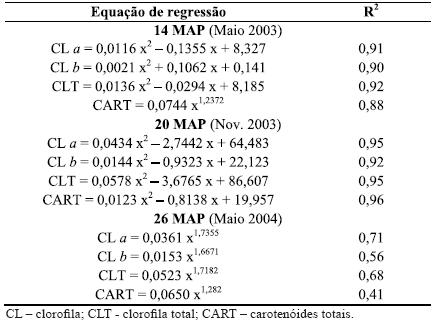
Contudo, o R2 diminuiu 26 MAP (Quadro 4) e ainda mais 32 MAP (inferior a 0,50 para todos os pigmentos, dados não incluídos). De facto, o aumento verificado nos teores dos pigmentos não foi acompanhado pelo aumento dos valores de SPAD e, por essa razão, esta técnica não foi utilizada nas últimas amostragens. Este padrão parece estar relacionado com a idade das árvores e a maturidade das folhas, relativamente às alterações que se verificam na sua estrutura interna e espessura, o que provavelmente afecta as leituras do SPAD a 940 nm (dependente da estrutura foliar). A coloração verde-escura típica das folhas de eucalipto adultas, associada ao aumento do teor de clorofila, pode ter afectado a absorção da luz a 650 nm, diminuindo a transmissão da luz neste comprimento de onda, como foi sugerido por Richardson et al. (2002). Donde, ter-se observado uma diminuição da transmissão para 940/650, valor que representa a leitura sem dimensões do SPAD, isto é, o teor relativo de clorofila.
Uma correlação (P<0,05) linear relativamente forte foi obtida num estudo preliminar (Madeira et al., 2009) entre o N foliar (mg g-1) e os valores de SPAD (R2=0,74) a 14 MAP (Maio 2003, dois meses após a primeira fertilização azotada), tendo porém diminuído (<0,15) nas outras amostragens. Maiores coeficientes de determinação foram obtidos, contudo, quando o N foliar foi expresso por unidade de área foliar (mg cm-2; Figura 2), como também relata-do por Chang & Robinson (2003).
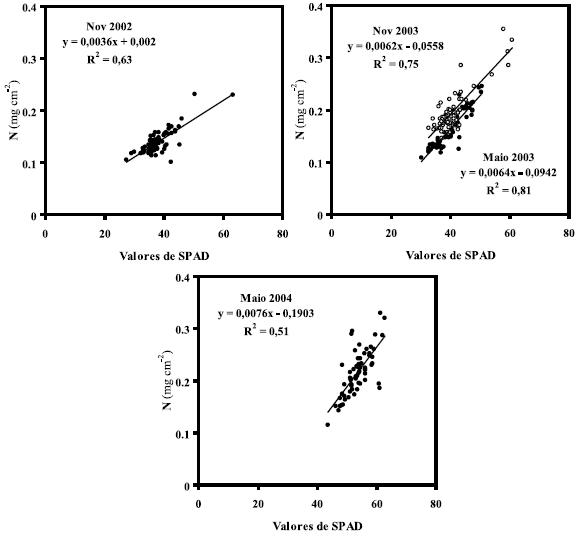
Figura 2 – Relação entre o teor de N (mg cm-2) e os valores de SPAD em folhas de E. globulus, a 8 (Nov. 2002), 14 (Maio 2003), 20 (Nov. 2003) e 26 (Maio 2004) MAP.
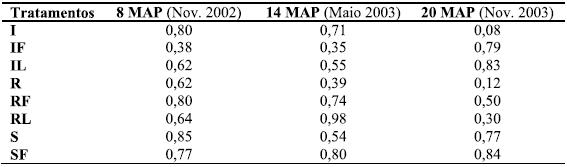
Alterações na morfologia das folhas, sazonalidade e condições nutricionais das árvores podem condicionar a correlação entre o teor de N e os valores de SPAD. Contudo, a sazonalidade não parece ter uma grande influência em R2 (Figura 2), visto o seu valor variar de 0,63 (Nov. 2002) e 0,75 (Nov. 2003) a 0,16 (Nov. 2004, dado não incluído), e de 0,81 a 0,51 em Maio de 2003 e 2004, respectivamente; de forma semelhante a idade da folha não parece ter afectado os teores de N e valores de SPAD até 20 MAP (Maio 2003). Os coeficientes determinados para cada tratamento (Quadro 5) variaram de 0,08 a 0,98, entre datas (8, 14 e 20 MAP) e tratamentos, e mostram a inexistência de uma tendência comum em termos de idade foliar ou sazonalidade, isto é, diminuem com a idade principalmente quando os resíduos são removidos (R, RL e RF), enquanto que aumentam quando os resíduos são incorporados (IL e IF) ou distribuídos à superfície (S e SF).
Quadro 5 – Coeficientes (R2) para correlações lineares entre a concentração de N (mg cm-2) e os valores de SPAD, para cada tratamento (n=8), a 8, 14 e 20 MAP.
Neste contexto, sugere-se que o medidor de clorofila pode ser usado para monitorizar o teor de azoto em E. globulus jovens (até dois anos de idade), desde que convenientemente calibrado.
CONCLUSÕES
O crescimento e o estado de nutrição de jovens árvores de E. globulus não parecem ter sido afectados pelos resíduos de abate, quer estes tenham sido mantidos ou removidos do sítio. A sua incorporação no solo promoveu o crescimento, mas as diferenças iniciais verificadas entre tratamentos esbateram-se com o tempo. A leguminosa L. luteus (IL e RL) gerou um aumento inicial do teor de N foliar, mas afectou negativamente o crescimento das árvores; contudo, no fim do estudo, o teor de N foi superior em IL, mas o crescimento foi semelhante ao de outros tratamentos. A aplicação de fertilizante azotado, pelo contrário, aumentou os teores de N e de pigmentos fotossintéticos, bem como os valores de SPAD, aproximadamente até 20 meses após a plantação, quer os resíduos de abate tenham sido mantidos ou removidos do sítio; o fertilizante promoveu o crescimento das árvores, principalmente quando os resíduos foram incorporados no solo. Atendendo à correlação entre o N foliar e os valores de SPAD, o medidor de clorofila parece ser uma técnica promissora para avaliar o teor de azoto em jovens plantações (até dois anos de idade). Nas condições Mediterrâneas em que o presente trabalho foi efectuado, o melhor sistema de gestão para o incremento da produtividade correspondeu à incorporação dos resíduos de abate associada à aplicação repetida de fertilizante azotado.
AGRADECIMENTOS
O estudo foi realizado no âmbito dos projectos AGRO 19 – OFLORINT (INIAP) e POCTI/AGG/42985/2001 (FCT). À empresa Celulose Beira Industrial S.A. (CELBI), proprietária da Quinta do Furadouro, agradecese a disponibilização e a supervisão da área experimental. Ao pessoal do laboratório de solos do Departamento de Ciências do Ambiente agradece-se as análises de solos e de materiais orgânicos. Agradece-se ainda o apoio de Mário Abrantes e Luís Hilário nas amostragens.
REFERÊNCIAS BIBLIOGRÁFICAS
Adams M.A. & Attiwill, P.M. (1986) -Nutrient cycling and nitrogen mineralisation in eucalypt forests of southeastern Australia. II. Indices of nitrogen mineralisation. Plant and Soil 92: 341-362.
Chang, S.X. & Robinson, D.J. (2003) -Nondestructive and rapid estimation of hardwood foliar nitrogen status using the SPAD-502 chlorophyll meter. Forest Ecology and Management 181: 331-338.
Cowie, A. L.; Smith, P. & Johnson, D. (2006) -Does soil carbon loss in biomass production systems negate the greenhouse benefits of bioenergy? Mitigation and Adaptation Strategies for Global Change 11: 979-1002.
Fabião, A.; Madeira, M. & Steen, E. (1990) -Effect of water and nutrient supply on root distribution in an Eucalyptus globulus plantation. Water, Air and Soil Pollution 54: 635-640.
Fisher, R.F. & Binkley, D. (2000) - Ecology and Management of Forest Soils (3rd ed.). John Wiley & Sons, New York.
Jones, H.E.; Madeira, M.; Herraez, L.; Dighton, J.; Fabião, A.; González-Rio, F.; Fernandez-Marcos, M.; Gomez, C.; Tomé, M.; Feith, H.; Magalhães, M.C. & Howson, G. (1999) -The effect of organic matter management on the productivity of Eucalyptus globulus stands in Spain and Portugal: tree growth and harvest residue decomposition in relation to site and treatment. Forest Ecology and Management 122: 73-86.
Lichtenthaler, H.K. (1987) -Chlorophylls and carotenoids: pigments of photosynthetic membranes. Methods in Enzymology 148:350-382.
Madeira, A.C.; Carneiro, M.; Marques, P. & Madeira, M. (2009) - Avaliação do estado de nutrição de plantas jovens de Eucalyptus globulus por análise foliar e espectroradiométrica. Revista de Ciências Agrárias 31, 3-26. [ Links ]
Madeira, A.C.; Mendonça, A.; Ferreira, M.E. & Taborda, M.L. (2000) - Relationship between spectroradiometric and chlorophyll measurements in green beans. Communications in Soil Science and Plant Analysis 31: 631-643.
Madeira, A.C. & Varennes, A. (2005) - Use of chlorophyll meter to assess the effect of nitrogen on sweet pepper development and growth. Journal of Plant Nutrition 28: 1-12.
Madeira, M.; Araújo, M.C. & Pereira, J.S. (1995) - Effects of water and nutrient supply on amount and on nutrient concentration of litter fall and forest floor litter in Eucalyptus globulus plantations. Plant and Soil 168-169: 287-295.
Madeira, M.; Melo, G.; Alexandre, C. & Steen, E. (1989) - Effects of deep ploughing and superficial disc harrowing on physical and chemical soil properties and biomass in a new plantation of Eucalyptus globulus. Soil Tillage Research 14: 163-175.
Magalhães, M.C. (2000) - Efeitos de Técnicas de Preparação do Solo e Gestão dos Resíduos Orgânicos em Características Físico-Químicas do Solo de Plantações Florestais. Dissertação de doutoramento, Universidade Técnica de Lisboa, Instituto Superior de Agronomia, Lisboa.
Malik, R.K.; Green, T.H.; Brown G.F.; Beryl, C.A.; Sistani, K.R. & Maes, D.A. (2001) -Biomass production of short-rotation bionergy hardwood plantations affected by cover crops. Biomass Bioenergy 21: 21-33
Mendham, D.S.; Kumarawamy, S.; Balasundaran, M.; Sankaran, K.V.; Corbeels, M.; Grove, T.S.; O´Connell, A.M. & Rance, S.J. (2004) -Legume cover cropping effects on early growth and soil nitrogen supply in eucalypt plantations in southwestern India. Biology and Fertility of Soils 39: 375-382.
Merino, A.; López, A.R.; Brañas, J. & Rodriguez-Soalleiro, R. (2003) -Nutrition and growth in newly established plantations of Eucalyptus globulus in northwestern Spain. Annals of Forest Science 60: 509-517.
Nzila, J.D.; Boillet, J.P.; Laclau, J.C. & Ranger, J. (2002) -The effects of slash management on nutrient cycling and tree growth in Eucalyptus plantations in the Congo. Forest Ecology and Management 171: 209-221.
Pereira, J.S.; Linder, S.; Araújo, M.C.; Pereira, H.; Ericsson, T.; Borralho, N. & Leal, L. C. (1989) -Optimization of biomass production in Eucalyptus globulus plantations: A case study. In: Pereira, J.S. & Landsberg, J.J. (Eds.) Biomass Production by Fast-Growing Trees. Kluwer Academic Publishers, Dordrecht, pp.101-121.
Pereira, J.S.; Tomé, M.; Madeira, M.; Oliveira, A. & Almeida, M.H. (1996) -Eucalypt plantations in Portugal. In: Attiwill, P.M. & Adams, M.A. (Eds.) Nutrition of the Eucalypts. CSIRO, Australia, pp. 371-387.
Pestana, M.; Varennes, A.; Goss, M.J.; Abadia, J. & Faria, E.A. (2004) - Floral analysis as a tool to diagnose iron chlorosis in orange trees. Plant and Soil 259: 287-295.
Pinkard, E.A.; Patel, V. & Mohammed, C. (2006) - Chlorophyll and nitrogen determination for plantation-grown Eucalyptus nitens and E. globulus using a non-destructive meter. Forest Ecology and Management 223: 211-217.
Powers, R.F.; Alban, D.H.; Miller, D.H.; Tiarks, A.E.; Wells, C.G.; Avers, P.E.; Cline, R.G.; Loftus, Jr.; N.S. & Fitzgerald, R.O. (1990) -Sustaining productivity in North America forests: problems and prospects. In: Gessel, S.P. et al. (Eds.) Sustained Productivity of Forest Soils. Proc. of 7th North American For. Soils Conf. Vancouver, Canada, 21-28 July 1988. University of British Columbia, Faculty For. Publ., Vancouver, pp. 49-79.
Proe, M. F. & Dutch, J. (1994) - Impact of whole tree harvesting on second-rotation growth of Sitka spruce: the first ten years. Forest Ecology and Management 65: 39-54.
Reis, R.M.M. & Gonçalves, M. Z. (1981) - Caracterização Climática da Região Agrícola do Ribatejo e Oeste. O Clima de Portugal. Fasc. XXXII. Instituto Nacional de Meteorologia e Geofísica, Lisboa
Ribeiro, H. M.; Vasconcelos, E.; Ramos, A. & Coutinho, J. (2009) – Avaliação do estado nutricional azotado de pés-mãe de Eucalyptus globulus Labill. ssp. globulus com recurso a um medidor portátil de clorofila. Revista de Ciências Agrárias 31: 40-49. [ Links ]
Richardson, A.D.; Duigan, S.P. & Berlyn, G.P. (2002) - An evaluation of non-invasive methods to estimate foliar chlorophyll content. New Phytologist 153: 185-194.
Shammas, K.; OConnell, A. M.; Grove, T. S.; McMurtrie, R.; Damon, P. & Rance, S.J. (2003) - Contribution of decomposing harvest residues to nutrient cycling in a second rotation Eucalyptus globulus plantation in south-western Australia. Biology and Fertility of Soils 38: 228-235.
Smethurst, P.J. & Nambiar, E.K.S. (1990) -Effect of slash and litter management on fluxes of nitrogen and tree growth in a Pi-nus radiata plantation. Canadian Journal of Forest Research 20: 1498-1507.
Soares, H.; Madeira, M.; Fabião, A.; Azevedo, A.; Kätterer, T.; Abreu, F. & Cortez, N. (2002) -Efeitos da gestão de resíduos de abate no crescimento inicial de Eucalyptus globulus Labill.: resultados de um estudo lisimétrico. Revista de Ciências Agrárias 25: 394-410. [ Links ]
Spangenberg, A.; Grimm, U.; Silva, J.R.S. & Folster, H. (1996) -Nutrient store and export rates of Eucalyptus globulus plantations in eastern Amazonia (Jari). Forest Ecology and Management 80: 225-234.
Stupak, I.; Asikainen, A.; Jonsell, M.; Karl-tun, E.; Lunnan, A.; Mizaraité, D.; Pasanen, K.; Pärn, H.; Raulund-Rasmussen, K.; Röser, D.; Schroeder, M.; Varnagiryté, I.; Vilkriste, L., Callesen, I.; Clarke, N.; Gaitnieks, T.; Ingerslev, M.; Mandre, M.; Ozolincius, R.; Saarsalmi, A.; Armolaiti, K.; Helmisaari, H.-S.; Indriksons, A.; Kairiukstis, L.; Katzensteiner, K.; Kukkola, M.; Ots K.; Ravn, H.P. & Tamminen, P. (2007) -Sustainable utilisation of forest biomass for energy – Possibilities and problems: Policy, legislation, certification, and recommendations and guidelines in the Nordic, Baltic, and other European countries. Biomass Bioenergy 31: 666-684.
Tomé, M.; Tomé, J.; Ribeiro, F. & Faias, S. (2007) - Equação de volume total, volume percentual e de perfil do tronco para Eucalyptus globulus Labill. em Portugal. Silva Lusitana 15: 25-39 [ Links ]
WRB (2006) - World Reference Base for Soil Resources. (2nd ed.) World Soil Resources Reports No. 103. FAO, Rome.
Xu, D. P.; Yang, Z.J. & Zhang, N.N. (2004) -Effect of site management on tree growth and soil properties of a second-rotation plantation of Eucalyptus urophylla in Guangdong Province, China. In: Nambiar E.K.S.; Ranger J.; Tiarks A. & Toma T. (Eds.) Site management and productivity in tropical plantations forests. Proceedings of Workshops in Congo July 2001 and China February 2003. Bogor, Indonesia, CIFOR, pp. 45-60.
Zbyszewski, G. & Almeida, F. (1960) - Carta geológica de Portugal (1:50 000). Notícia explicativa da folhas 26D (Caldas da Rainha). Serviços Geológicos de Portugal, Lisboa.
1 Instituto Superior de Agronomia, Universidade Técnica de Lisboa, Tapada da Ajuda, 1349-017 Lisboa, Portugal. e-mail: acmadeira@isa.utl.pt
Recepção/Reception: 2009.07.28
Aceitação/Acception: 2009.08.31

