










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias v.33 n.2 Lisboa dez. 2010
Crescimento e teores de clorofila em mudas de bananeira em função da supressão do pseudocaule, de doses de nitrogênio e de boro
Walter Esfrain Pereira1; José Pires Ribeiro Nóbrega2; Thiago Jardelino Dias2; Roberto Wagner Cavalcante Raposo3; Raunira da Costa Araújo4 & Francisco Assis de Oliveira3
1Departamento de Ciências Fundamentais e Sociais, CCA/UFPB, Campus II, Cidade Universitária, Areia-PB, Cep: 58397-000. Fone: 83-3362-2291. E-mail: wep@cca.ufpb.br
2Programa de Pós-Graduação em Agronomia, CCA/UFPB, Campus II, Cidade Universitária, Areia-PB, Cep: 58397-000. E-mail: tjardelino@hotmail.com
3Programa de Pós-Graduação em Manejo do Solo e da Água, CCA/UFPB, Campus II, Cidade Universitária, Areia - PB, Cep: 58397-000. E-mail: rwcraposo@cca.ufpb.br; oliveira@cca.ufpb.br
4Centro de Formação de Tecnólogos, UFPB/Campus III, Bananeiras - PB. Cep: 58220-000, E-mail: raunira@cft.ufpb.br
RESUMO
O objetivo deste experimento foi avaliar a influência da eliminação da gema apical do rizoma e de doses de nitrogênio e boro, sobre a produção e o crescimento de mudas de bananeira. O experimento foi realizado no Centro de Formação de Tecnólogos da UFPB. O delineamento foi em blocos casualizados, com quatro blocos e nove plantas matrizes por parcela, sendo duas plantas úteis. Os fatores avaliados foram doses de N (0 a 240 g/planta) e de B (0 a 2,2 g/planta) combinados com a matriz experimental Composto Central de Box, originando nove combinações, arranjados fatorialmente com e sem eliminação da gema apical da planta matriz. Os dados foram submetidos à análise de variância e de regressão. A supressão do pseudocaule da planta principal, com a eliminação da gema apical do rizoma aumentou o número de perfilhos, com maior crescimento, exceto do diâmetro do rizoma, o qual diminuiu. Os teores foliares de clorofila total e de B também foram diminuídos, nos perfilhos das bananeiras amputadas. Para a produção de mudas da bananeira Pacovan, recomenda-se a supressão do pseudocaule da planta principal, com eliminação da gema apical do rizoma e aplicação do N.
Palavras-chave: Brasil, clorofila, Musa sp., propagação, perfilhos.
Growth and chlorophyll content of banana suckers in function of pseudostem suppression and doses of nitrogen and boron
ABSTRACT
The objective of this research was to evaluate the influence of the removal in the pseudostem with elimination of the meristem apical of the rhizome and doses of nitrogen and boron, about the production and growth of banana suckers. The experiment was carried out at Centro de Formação de Tecnólogos - UFPB, State of Paraíba. The experimental design was in randomized blocks, with four replications and nine mother plants for experimental unit, being two useful plants. The evaluated valued factors were doses of N (0 to 240 g plant-1) and of B (0 to 240 g plant-1) combined in agreement with the experimental matrix Central Composite, originating nine combinations, which were arranged factorially with suppression and without suppression of the pseudostem of the mother plant. The data were submitted the variance analysis and of regression. The suppression in the pseudostem of the mother plant with elimination of the apical meristem of the rhizome increased the sucker number, with larger growth, except of the diameter of the rhizome which it reduced. In the plants with suppression, it reduced the sucker number which presented smaller growth. On the other hand, it increased the content of total chlorophyll. For the production of Pacovan banana suckers, is recommended the suppression in the pseudostem of the mother plant with elimination of the meristem apical of the rhizome and the application of the N.
Key-words: Brasil, chlorophyll, Musa sp. propagation, sucker.
INTRODUÇÃO
A banana ocupa o segundo lugar em volume de frutas produzidas no Brasil, perdendo apenas para a laranja, sendo cultivada predominantemente em pequenas propriedades, onde são colhidos 60% da produção nacional, tendo grande importância na fixação do homem no campo e na geração de emprego e renda, especialmente para as camadas carentes da população (Bernardo et. al., 2004; Embrapa, 2005). A despeito dessa grande produção, a produtividade é baixa e as exportações têm oscilado, nos últimos anos, em torno de apenas 1% do total produzido internamente. Barros & Pizzol (2001) sugerem que este baixo índice pode estar relacionado ao elevado consumo interno, além de outros fatores.
No Brasil, o número de cultivares que produzem frutos que satisfação os requisitos e características necessárias para exportação são reduzidas, destacando as cultivares de mesa do subgrupo Cavendish (Nanicão, Grand Naine e Williams). No entanto, diversas variedades regionais são consideradas de interesse comercial, pois atendem a preferência do consumidor (Souto et al., 1999; Moreira & Fageria, 2008). Nas últimas décadas, tem sido crescente a ampliação da área plantada com a bananeira cultivar Pacovan (Musa sp. AAB, subgrupo Prata), a qual ocupa uma expressiva fatia do mercado Nordestino, parte desta antes ocupada pela bananeira Prata (Musa sp. AAB), variedade semelhante, mas menos produtiva que aquela. Linden (1985) afirma que, na microrregião do Brejo Paraibano, a cultivar Pacovan destaca-se em termos de área plantada em relação às demais, sendo considerada uma variedade regional. Essa predominância ainda existe na atualidade.
Não obstante a grande produção nacional, o crescimento da bananicultura encontra uma série de obstáculos, que têm contribuído para os baixos índices de produtividade e qualidade do fruto. Dentre os entraves sobressaem a falta de mudas em quantidade e qualidade necessárias para implantação de bananais comerciais, controle fitossanitário ineficiente, manejos indevidos na colheita, pós-colheita e logística, que provocando injurias nos frutos, refletindo na qualidade do produto comercializado e consequentemente, baixa eficiência na comercialização (Ruggiero & Gottardi, 1997).
A bananeira é uma planta exigente em nutrientes e água, não só por produzir grande massa vegetativa, mas também por apresentar elevadas quantidades de elementos absorvidos pela planta e exportados pelos frutos (Silva et al., 1999). A bananeira também é exigente em água, sendo que sua deficiência promove redução da clorofila das folhas e leva-as à morte prematura, retardando o crescimento (Turner, 1994; Cayón Salinas, 2001) e, consequentemente, a produção (Coelho et al., 2006).
O nitrogênio exerce grande importância no início do desenvolvimento da bananeira até a emissão da inflorescência, havendo diferenciação entre as parte vegetativas que constituem a bananeira, quanto à concentração e extração ou acúmulo de nutrientes, tendo entre o rizoma, pseudocaule e as folhas, as maiores concentrações de nitrogênio está presente no pseudocaule (Diniz et al., 1999).
Na propagação de mudas de bananeira, a deficiência do boro promove dificuldade de brotação de mudas e deformações morfológicas nas folhas novas. Essas deformações são limbos estreitos com ondulações nos bordos; às vezes, permanece somente a nervura central. Promove ainda outros sintomas tais como necrose nas margens das folhas sem clorose prévia, principalmente na ponta a qual se encarquilha, plantas que não produzem clorose internerval e estrias perpendiculares às nervuras secundárias na face inferior, além do aparecimento de muitos filhotes com os sintomas ainda mais acentuados. Em casos extremos de carência, ocorre a paralisação do crescimento, e, por outro lado, o excesso de boro pode acarretar, na folha, clorose marginal seguida de necrose (Lahav, 1995). Nestes casos as plantas não produzem frutos.
O objetivo desta pesquisa foi avaliar a influência da eliminação da gema apical do rizoma (10 cm acima do nível superficial do solo e aos 151 dias após o plantio), com aplicação de doses de nitrogênio e boro sobre a produção, crescimento e o teor de clorofila de perfilhos (mudas) da bananeira Pacovan.
MATERIAL E MÉTODOS
Este experimento foi conduzido no Centro de Formação de Tecnólogos (CFT) da Universidade Federal da Paraíba (UFPB), Estado da Paraíba (PB) - Brasil, e decorreu no período de abril de 2005 a fevereiro de 2006. A área experimental possui altitude de 552 metros, apresenta clima quente e úmido que corresponde ao clima submediterrâneo (Brasil, 1972). O índice pluviométrico correspondente ao período de execução do experimento em campo (junho/2005 a fevereiro/2006) foi de 268 mm, tendo a temperatura e umidade relativa do ar variado entre 20-260 C e 76-93%, respectivamente.
As mudas de bananeira, cultivar Pacovan (Musa sp. AAB, subgrupo Prata), foram originárias de cultura de tecidos de ápices caulinares e adquiridas, com cerca de 6 a 7 cm de altura. Procedeu-se aclimatização das mudas em viveiros, realizando a repicagem das mudas em recipientes de polietileno preto com 20 cm de largura e 30 cm comprimento, sanfonados e perfurados em seu terço inferior, contendo substrato peneirado e solarizado, composto por terra argilo-arenosa de superfície e composto orgânico vegetal, na proporção de 3:1, respectivamente (Katan et al., 1976; Ricci et al., 1999).
O tempo de aclimatização das mudas foi de 81 dias, sendo um período de 61 dias sob telado, com 50% de luminosidade, e outro de 20 dias em viveiro a céu aberto, para completarem o seu endurecimento, até o plantio no campo. Durante o desenvolvimento das mudas, foi realizada adubação em cobertura, via solo, aos 42 dias após repicagem das mudas nos recipientes, aplicando-se por planta, em círculo e próximo as bordas dos recipientes, uma mistura composta por 2,5 g de uréia, 4,0 g de superfosfato simples e 1,2 g de cloreto de potássio. Foram realizadas ainda, quatro adubações foliares, com intervalos de 15 dias, iniciando aos 35º dia após a repicagem das mudas nos recipientes, com uréia (2,5 g/L) mais sulfato de magnésio (2,5 g/L); a segunda repedindo a aplicação da primeira; a terceira utilizando uréia (3,0 g/L) mais sulfato de zinco (2,0 g/L); e, a quarta aplicando sulfato de cobre (2,5 g/L) mais cal hidratada (5,0 g/L).
As irrigações foram realizadas periodicamente, sendo que, nos primeiros 15 dias de aclimatização as regas tinham maior frequência, porém aplicava-se menor lâmina de água, utilizando pulverizador costal manual, para simular uma nebulização sobre as folhas e melhorar a aeração do substrato (Lima et al., 2003), em intervalos de aplicação de aproximadamente uma hora, os quais foram sendo elastecidos com o avanço do processo de aclimatação. Posteriormente, passou-se a utilizar o sistema de irrigação por microaspersão, a intervalos menos freqüentes.
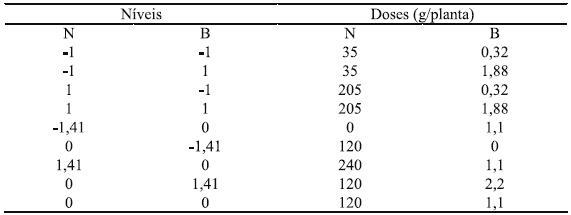
Os tratamentos foram distribuídos em delineamento em blocos casualizados, com quatro blocos e nove plantas matrizes por parcela (composta por três linhas com três plantas cada), sendo duas plantas úteis (na linha central) por unidade experimental. Os fatores avaliados foram doses de nitrogênio (N) e de boro (B) combinados de acordo com a matriz experimental Composto Central de Box, originando nove combinações (Quadro 1), as quais foram combinadas fatorialmente com dois tipos de supressão no pseudocaule: supressão a 10 cm acima da superfície do solo e sem supressão, totalizando 18 tratamentos.
Quadro 1 – Níveis e doses de nitrogênio (N) e de boro (B) combinados de acordo com matriz experimental Composto Central de Box.
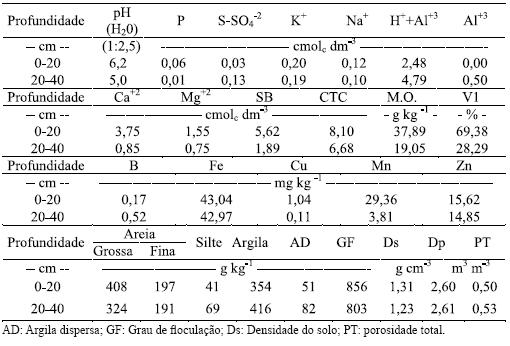
A caracterização da fertilidade do solo e das características físicas foram determinadas antes da implantação do experimento, procedendo à coleta de amostras compostas às profundidades de 0 a 20 cm e de 20 a 40 cm e submetidos a análise para fertilidade completa e física, cujos resultados estão apresentados no Quadro 2.
Quadro 2 – Características químicas e físicas do solo da área experimental, nas profundidades de 0 a 20 cm e de 20 a 40 cm.
O preparo da área experimental foi constituído de aração, gradagem e abertura das covas nas dimensões de 40 x 40 x 40 cm (Alves & Lima, 2000), obedecendo ao espaçamento de viveiro de 1,5 x 1,5 m (Silva et al., 1999), em configuração quadrangular (Soto-Ballestero, 1992). Cada bloco e suas respectivas unidades experimentais foram constituídos por três linhas de plantio.
As covas foram preparadas 30 dias antes do transplantio das mudas em campo com substrato composto de um material retirado dos locais das respectivas covas, adicionando 80 g/cova de P2O5, na forma de superfosfato simples e 40 g/cova de K2O, na forma de cloreto de potássio, além de um calcário dolomítico (715 g/cova com PRNT 65%) e esterco bovino curtido (5,0 dm3/cova), que após homogeneização foram acondicionados no interior das respectivas covas (Alvez, 1986).
As mudas (plantas matrizes) de bananeira após o período de aclimatação (81 dias) foram plantadas em solo úmido, com torrão aderido às raízes, quando apresentavam cerca de 30 a 35 cm de altura, e manejadas por sistema de irrigação localizado (microaspersão), aplicando-se o suprimento de água de acordo com a necessidade da cultura, os índices de precipitação e a disponibilidade de água acumulada nos reservatório da área experimental.
As adubações estudadas foram aplicadas em cobertura, incorporando superficialmente em faixa circular com cerca de 10 cm de largura na primeira aplicação e a 20 cm de largura, nas demais, da base do pseudocaule da planta matriz e, ou da brotação lateral (Borges & Oliveira, 2000). As doses foram fracionadas aplicando-se na área experimental a primeira parcela (25% do N e 50% do B) aos 40 dias após o plantio das mudas; a segunda (25% do N e 50% do B), aos 78 dias; e, a terceira e a quarta (25% do N), aos 117 e 148 dias.
A quebra de dominância apical, para favorecer a brotação lateral e produção de mudas, foram realizadas aos 151 dias após o plantio, seis dias após a primeira coleta de perfilhos e três dias após a ultima aplicação do N em cobertura, sendo que, a supressão pseudocaule das plantas matrizes correspondentes aos tratamentos com supressão (em bisel e a 10 cm acima do nível superficial do solo) (Silva et al., 1999). Imediatamente após essa supressão, foi feita a eliminação da gema apical de crescimento do rizoma, com auxílio da ferramenta denominada lurdinha (Moreira, 1999), desinfetada com água clorada.
Com auxílio de cavador, desinfetado com água clorada, e de régua graduada em mm, foram separados da planta matriz (coletados), os perfilhos ou rebentos com altura igual ou superior a 15,0 cm, determinada entre a distância da superfície do solo ao ponto de interseção das duas últimas folhas (Santos, 2002). Com intervalos de 28 dias, foram realizadas quatro coletas: aos 145, 173, 201 e 229 dias após o plantio.
Após a separação da planta matriz, através de corte bem rente, os rizomas foram lavados com água, embalados em sacolas plásticas etiquetadas e os perfilhos foram encaminhados para as análises no Laboratório de Controle de Qualidade de Alimentos do CFT/UFPB.
Os perfilhos coletados aos 145 dias após o plantio (primeira coleta) foram avaliados apenas o seu número, pois, naquela oportunidade ainda não havia sido feito o último parcelamento de nitrogênio em cobertura e nem a supressão com eliminação da gema apical do rizoma.
No entanto, quando todas as atividades relativas aos tratamentos foram concluídas, os perfilhos coletados aos 173, 201 e 229 dias após o plantio, procedeu-se avaliação das seguintes características: o número médio de perfilhos produzidos por planta matriz; altura média do pseudocaule dos perfilhos; diâmetro médio do rizoma dos perfilhos; peso médio da matéria seca foliar dos perfilhos adotando procedimento proposto pela Embrapa (1999); e teores foliares de clorofila total pelo método de Arnon (1949), dos perfilhos coletados aos 201 e 229 dias após o plantio.
Os dados obtidos foram submetidos à análise de variância e de regressão. Os efeitos do N e do B foram avaliados mediante regressão polinomial, enquanto os efeitos das supressões foram avaliados pelo teste F. Em face da heterogeneidade das variâncias, transformou-se os dados referentes à variável número de perfilhos em .
RESULTADOS E DISCUSSÃO
Constatou-se efeito significativo da supressão da gema apical do rizoma nas plantas de bananeira durante o período em estudo, sobre o número de perfilhos produzidos por planta matriz, aos 173, 201 e 229 dias após o plantio. Não foi verificada influência significativa da combinação N e B nem da interação entre N e B e supressão.
Na Figura 1, observa-se que a partir de 173 dias após o plantio, as plantas sujeitas à supressão da gema apical produziram significativamente mais perfilhos do que as plantas não sujeitas àquela operação, e que, os perfilhos das plantas de bananeira Pacovan com supressão apresentaram pseudocaule com maior altura do que os perfilhos das plantas sem supressão. O maior número de perfilhos e a maior altura foram observados aos 173 dias após o plantio, em plantas suprimidas.
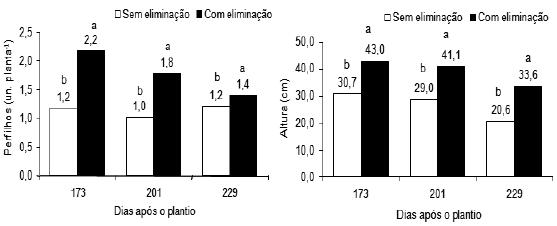
Figura 1 – Número de perfilhos e altura de plantas da bananeira Pacovan, com e sem supressão, em três períodos pós-plantio; Médias com letras diferentes diferem significativamente, em cada período pós-plantio (F, P < 0,01; F, P < 0,05; aos 229 dias após o plantio, respectivamente).
Para número de perfilhos produzidos por planta sujeita à supressão do pseudocaule, aos 201 dias após o plantio, pode verificar-se que foi significativa a regressão linear (Figura 2), indicando que à medida que aumentaram as doses de N, maiores foram os valores para produção de perfilhos por planta, cujo incremento foi de 0,002 perfilhos por cada 1 g/planta de N adicionada ao solo.
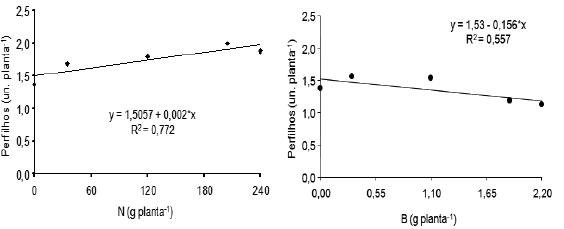
Figura 2 – Efeito de doses de nitrogênio e de boro sobre o número de perfilhos de plantas da bananeira Pacovan, com supressão, aos 201 e aos 229 dias após o plantio, respectivamente. *: Significativo a 5% pelo teste F.
Nas plantas também sem gema apical, aos 229 dias após o plantio, o B exerceu influência linear decrescente sobre o número de perfilhos produzidos por planta, diminuindo o número de perfilhos com o aumento das doses de B aplicadas (Figura 2).
O maior número de perfilhos produzido pelas plantas suprimidas com eliminação em relação às sem aquela operação ratifica a hipótese do controle hormonal de desenvolvimento de gemas laterais pelo meristema apical, sendo a auxina o principal hormônio envolvido nesse processo, representado possivelmente, pelo ácido indolacético, sintetizado na região apical e transportado para os meristemas laterais (Awad & Castro, 1983). A dominância apical se manifesta por inibição do crescimento das gemas axilares, ou laterais, pela presença da gema apical, sendo os meristemas apicais e folhas jovens os principais sítios de síntese de auxinas na parte aérea da planta, ocorrendo uma ação indireta na dominância de gemas e brotos (Philips, 1969).
Os resultados verificados para N são compatíveis com relatos de vários autores, tais como Silva et al. (1999) e Borges & Oliveira (2000), os quais afirmam que o N é o segundo elemento mais importante para a bananeira e favorece a emissão e o crescimento de filhotes ou rebentos. A despeito disso, Fontes et al. (2003) obtiveram resultados contraditórios, concluindo que houve diminuição na emissão de filhotes com o incremento das doses de N.
Vários outros autores conseguiram, através da propagação rápida in vivo, obter resultados melhores ou mais discretos que os desta pesquisa, no entanto, é conveniente ressaltar que, além das especificidades da metodologia usada particularmente, adubações e condições edafoclimáticas locais, entre outras, existe o importante fator, variedade, o qual, segundo constatações de pesquisas (Silva, 2000; Silva et al., 2000), apresenta diferenças de performance na emissão de rebentos ou perfilhos.
Estudos realizados na Universidade Estadual de São Paulo (UNESP), verificaram que a propagação da bananeira através de morfogênese indireta in vivo, resultou em número médio de 5,0; 4,0; 5,2; 5,0; 4,1; e 4,1 rebentos por rizoma, após 150 dias, respectivamente, para as variedades Nanicão, Maçã, Prata-Anã, Fhia-1 e Fhia-18, ratificando, assim, a existência de variedades mais eficientes que outras, em relação à produção de mudas (Silva, 2000; Silva et al., 2000). Constatou-se também que a primeira brotação, além de ser menos trabalhosa, resultou em maior produção de rebentos (Silva, 2000).
Aos 229 dias após o plantio, a altura do pseudocaule dos perfilhos das plantas não suprimidas amputadas comportou-se de maneira linear positiva em função das doses de B testadas, apresentando aumento estimado de 1,78 cm para cada incremento de 1 g/planta de B adicionada ao solo (Figura 3). Aos 201 dias após o plantio verificou-se efeito linear do B (Figura 3) sobre a altura do pseudocaule de perfilhos de plantas amputadas, indicando que à medida que aumentaram as doses de B, diminuíram os valores de altura do pseudocaule dos perfilhos, cujo decréscimo foi de 4,93 cm para cada incremento de 1 g/planta de B, resultado semelhante ao obtido para número de perfilhos produzidos por planta.
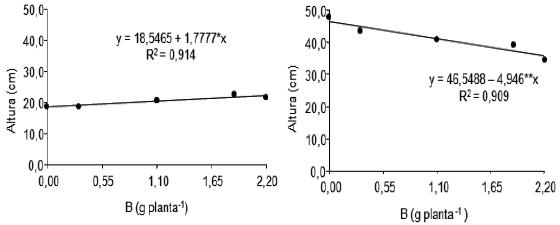
Figura 3 – Efeito de doses de boro sobre a altura do pseudocaule de perfilhos de plantas da bananeira Pacovan, sem supressão, aos 229 dias e com supressão, aos 201 dias após o plantio.
*,**: Significativo a 5 e a 1%, respectivamente pelo teste F.
Verificou-se influência significativa da combinação N e B sobre o diâmetro do rizoma dos perfilhos de bananeira Pacovan, apenas aos 201 dias após o plantio. O efeito da supressão foi significativo sobre o diâmetro do rizoma dos perfilhos, aos 173, 201 e 229 dias após o plantio (Figura 4).
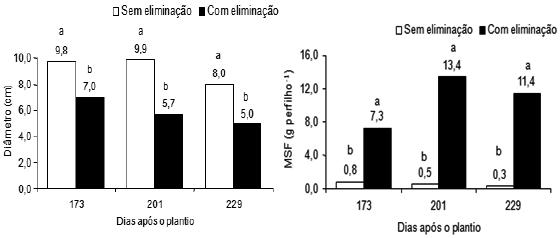
Figura 4 – Diâmetro do rizoma e peso da matéria seca foliar (MSF) dos perfilhos de plantas da bananeira Pacovan, com e sem supressão, em três períodos pós-plantio. Médias com letras diferentes diferem significativamente, em cada período pós-plantio (F, P < 0,01).
As plantas que não foram amputadas apresentaram perfilhos com maior diâmetro de rizoma do que as plantas que sofreram aquela operação, com diferenças significativas de 2,8; 4,2; e 3,0 cm aos 173, 201 e 229 dias após o plantio, respectivamente. Os maiores diâmetros de rizoma dos perfilhos, 9,9 cm e 9,8 cm, foram observados, respectivamente, aos 201 e 173 dias após o plantio, em plantas sem supressão (Figura 4).
As doses de N e de B não exerceram influência significativa no diâmetro do rizoma de perfilhos das plantas não amputadas, em nenhum período pós-plantio estudado. No entanto, nas plantas amputadas, a aplicação de N e B teve efeito linear significativo aos 201 dias após o plantio.
Observa-se na Figura 5, que aos 201 dias após o plantio, à medida que aumentaram as doses de N e B, diminuiu o diâmetro de rizoma dos perfilhos das plantas com supressão, à razão de 4,0 mm e de 0,54 cm, respectivamente, para cada incremento de 1 g/planta de N e B adicionada ao solo.
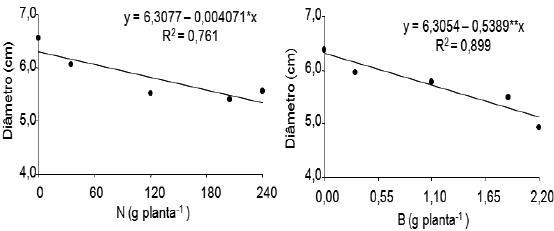
Figura 5 – Efeito de doses de nitrogênio e do boro sobre o diâmetro do rizoma de perfilhos de plantas da bananeira Pacovan, com supressão, aos 201 dias após o plantio.
*,**: Significativo a 5 e a 1%, respectivamente pelo teste F.
Foi constatada influência significativa da supressão sobre o peso da matéria seca foliar (MSF) dos perfilhos de plantas de bananeira, nos três períodos pós-plantio estudados. Foi verificada apenas nas plantas amputadas, a aplicação das doses de N e de B teve influência significativa sobre o peso da MSF dos perfilhos aos 173 e 201 dias após o plantio, respectivamente (Figura 6).
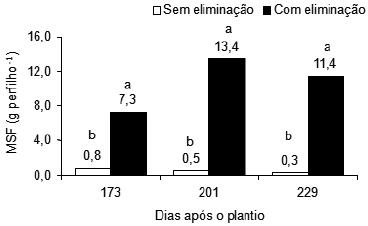
Figura 6 – Peso da matéria seca foliar (MSF) dos perfilhos de plantas da bananeira Pacovan, com e sem supressão, em três períodos pós-plantio. Médias com letras diferentes diferem significativamente, em cada período pós-plantio (F, P < 0,01).
Verificou-se uma resposta positiva da supressão sobre o peso da MSF dos perfilhos. A partir de 173 dias após o plantio, os perfilhos das plantas com supressão apresentaram significativamente maiores pesos da matéria seca de folhas do que os perfilhos das plantas sem supressão (Figura 6). O maior peso da MSF dos perfilhos ocorreu aos 201 dias após o plantio, nas plantas com supressão. A supressão, de modo geral, aumentou o peso da MSF dos perfilhos de plantas da bananeira Pacovan.
Nas plantas com supressão, houve aumento linear significativo do peso da matéria seca foliar dos perfilhos à medida que aumentaram as doses de N, aos 173 dias após o plantio (Figura 7). O aumento observado foi de 15 mg para cada incremento de 1 g/planta de N adicionada ao solo. Esse aumento de matéria seca, em perfilhos de bananeira Pacovan, favorecido pelo N, está em consonância com relatos de Borges et al. (1997), Silva et al. (1999) e Borges & Oliveira (2000), quando citam que o N é muito importante para o crescimento vegetativo da bananeira e aumenta bastante a quantidade de matéria seca.
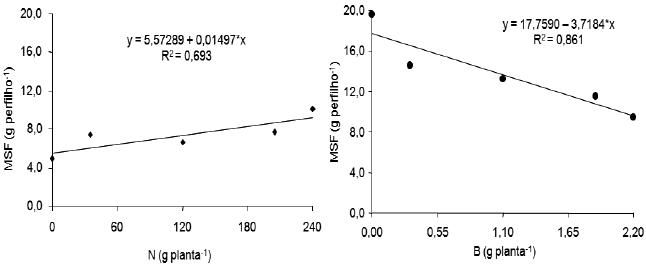
Figura 7 – Efeito de doses de nitrogênio e do boro sobre a matéria seca foliar (MSF) dos perfilhos de plantas da bananeira Pacovan, com supressão, aos 173 e aos 201 dias, respectivamente, após o plantio.
*: Significativo a 5% pelo teste F.
Também nas plantas com supressão, aos 201 dias após o plantio, o peso da matéria seca foliar dos perfilhos seguiu uma tendência linear negativa em função das doses de B (Figura 7), sendo o decréscimo de 3,72 g para cada 1 g/planta de B adicionada ao solo.
Quanto ao teor de clorofila dos perfilhos aos 229 dias após o plantio, foram verificados efeitos significativos da combinação N e B, supressão e interação entre N e B e supressão sobre a clorofila total foliar de perfilhos de plantas da bananeira Pacovan (Figura 8).
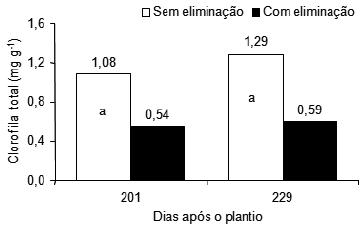
Figura 8 – Teor de clorofila total foliar de perfilhos de plantas da bananeira Pacovan, com e sem supressão, em dois períodos pós-plantio e efeito de doses de boro sobre o teor de clorofila total foliar de perfilhos de plantas da bananeira Pacovan, com supressão, aos 229 dias após o plantio. Médias com letras diferentes diferem significativamente, em cada período pós-plantio (F, P < 0,01);
*: Significativo a 5% pelo teste F.
Aos 201 dias após o plantio, não houve efeito significativo de doses de N nem de B sobre a clorofila total foliar de perfilhos de plantas de bananeira Pacovan com e sem supressão. Resultado similar foi observado aos 229 dias após o plantio, para os perfilhos das plantas sem supressão. Nos perfilhos das plantas de bananeira com supressão houve efeito linear significativo das doses de B sobre o teor de clorofila total foliar.
O fato de ocorrer, neste trabalho, significativamente maior teor foliar de clorofila total justamente naqueles perfilhos de plantas sem supressão (Figura 8), os quais apresentavam menores teores foliares de N que os perfilhos das plantas com supressão, contraria resultados que afirmam que o N apresenta importante função como integrante da molécula de clorofila, (Lichtenthaler, 1987; Büll, 1993), onde cada átomo de Mg está ligado a quatro átomos de N (Monteiro et al., 1995), plantas deficientes em N têm baixa capacidade e eficiência fotossintética, e que doses elevadas prolongam a fase vegetativa e elevam o período de fotossíntese (Hendry & Price, 1993; Marschner, 1995). Isso pode ser atribuído, em parte, provavelmente ao fato de os perfilhos originados das plantas com supressão apresentarem maior expansão folia (Ronchi et al., 2001).
Na Figura 8, observa-se que o B exerceu, aos 229 dias após o plantio, influência linear positiva e significativa sobre o teor de clorofila total foliar dos perfilhos eliminados em plantas de bananeira Pacovan, indicando que a clorofila total aumentou linearmente com o aumento das doses de B aplicadas, cujo incremento foi de 0,07 mg/g para cada 1 g/planta de B adicionada ao solo.
Este resultado expressa a importância do B para a fotossíntese (Pilbeam & Kirkby, 1983) e para a síntese de bases nitrogenadas como a uracila (Albert, 1968), a qual é componente essencial do RNA e, se ausente, afetará a proteossíntese. A propósito, a deficiência deste micronutriente nas bananeiras causa diversos sintomas, tais como: necrose nas folhas, listras amarelo-esbranquiçadas espalhadas na superfície da folha, goma no pseudocaule (Borges & Oliveira, 2000; Cordeiro & Borges, 2000), paralisação do crescimento das partes terminais e secagem das folhas novas (Raij, 1991).
Os valores médios de clorofila total foliar, aos 229 dias após o plantio, Figura 9, variaram de 0,51 a 0,74 mg/g nos perfilhos das plantas amputadas e, de 0,69 a 2,17 mg/g, nos perfilhos das plantas sem supressão. O maior teor de clorofila total foi observado no tratamento 7 (240 g/planta de N + 1,1 g/planta de B), seguido do tratamento 9 (120 g/planta de N + 1,1 g/planta de B), em perfilhos de plantas sem supressão.
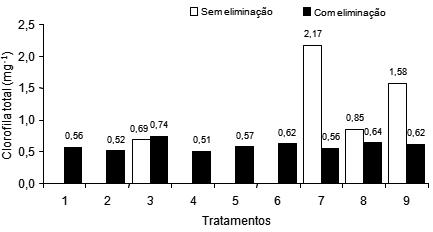
Figura 9 – Teor de clorofila total foliar de perfilhos de plantas da bananeira Pacovan, em função dos tratamentos, com e sem supressão, aos 229 dias após o plantio.
Nos estádios de desenvolvimento das plantas de bananeira Pacovan aos 201 e 229 dias após o plantio, os teores médios de clorofila total foliar foram significativamente maiores nos perfilhos das plantas sem supressão do que nos perfilhos das plantas com supressão. Este resultado expressa o efeito da supressão sobre a clorofila total foliar dos perfilhos das plantas de bananeira Pacovan. O maior teor médio de clorofila total foliar, 1,29 mg/g, foi observado nos perfilhos das plantas sem supressão, aos 229 dias após o plantio, independentemente dos tratamentos entre doses de nitrogênio e de boro. Este resultado pode ser atribuído à redução da parte aérea das plantas matrizes em decorrência do efeito da supressão.
CONCLUSÕES
A supressão do pseudocaule com a eliminação da gema apical do rizoma aumentou o número, a altura do pseudocaule e a massa da matéria seca foliar e diminuiu o diâmetro do rizoma dos perfilhos;
Os teores de clorofila total e de boro, nas folhas dos perfilhos da bananeira, diminuíram com a supressão da planta mãe;
O incremento das doses de nitrogênio aumentou o número de perfilhos e a massa da matéria seca foliar e diminuiu o diâmetro do rizoma dos perfilhos das plantas com supressão;
O aumento das doses de boro diminuiu o número perfilhos, a altura do pseudocaule, o diâmetro do rizoma e a massa da matéria seca foliar dos perfilhos das plantas amputadas e aumentou a altura do pseudocaule dos perfilhos das plantas não amputadas;
O incremento das doses de boro aumentou o teor foliar de clorofila total dos perfilhos das plantas suprimidas;
Para a produção de mudas da bananeira Pacovan recomenda-se a supressão no pseudocaule da planta matriz, com a eliminação da gema apical do rizoma e a aplicação do nitrogênio, sendo necessárias outras pesquisas para determinar a dose recomendada do mesmo.
REFERÊNCIAS BIBLIOGRÁFICAS
Albert, L.S. (1968) - Induction and antagonism of boron-like deficiency symptoms of tomato plants by selectded nitrogen-bases. Plant Physiology 43, 51-54.
Alves, E.J. & Lima, M.B. (2000) - Estabelecimento do bananal. In: Cordeiro, Z.J.M. (Ed.) Banana. Produção: aspectos técnicos. Brasília, DF: Embrapa Comunicação para Transferência de Tecnologia, pp. 73-82.
Alves, E.J. & Macedo, M.M.C. (1986) - Lurdinha, desbaste eficiente na bananeira. Bnb/Etene, Fortaleza, Brasil, pp. 460-462. (Seminário de tecnologia agropecuária inovadora para o nordeste).
Alvez, E.J.A. (1986) - Bananicultura brasileira e o programa de pesquisa coordenada pela Embrapa em prol do seu melhoramento. Cruz das Almas: EMBRAPA-CNPMF, (EMBRAPA-CNPMF. Documentos, 17), 47pp.
Arnon, D.I. (1949) - Copper enzymes in isolated chloroplasts. Polyphenoloxidade in Beta vulgaris. Plant physiology 24, 1: 1-15.
Awad, M. & Castro, P.R.C. (1983) - Introdução à fisiologia vegetal. Editora Nobel, São Paulo, 177pp.
Barros, E.M. & Pizzol, S.J.S. (2001) - Fruticultura: Comércio Internacional de Banana. Preços Agrícolas. São Paulo Piracicaba.
Bernardo, W.F.; Rodrigues, B.I.; Cassiere Neto, P.; Ando, A.; Tulmann Neto, A.; Ceravolo, L.C. & Montes, S.M.N.M. (2004) - Micropropagação de baixo custo em bananeira cv. Maçã em meios com diferentes fontes de carbono e avaliação da performance em campo das mudas produzidas. Revista Brasileira de Fruticultura 26, 3: 503-506.
Borges, A.L.; Alves, E.J.; Silva, S.O.; Souza, L.S.; Matos, A.P.; Fancelli, M.; Oliveira, A.M.G.; Cordeiro, Z.J.M.; Silveira, J.R.S.; Costa, D.C.; Medina, V.M.; Oliveira, S.L.; Souza, J.S.; Oliveira, R.P.; Cardoso, C.E.L.; Matsura, F.C.A.U. & Almeida, C.O. (1997) - O cultivo da banana. Embrapa-Cnpmf, Cruz das Almas, 109 pp.
Borges, A.L. & Oliveira, A.M.G. (2000) - Nutrição, Calagem e Adubação. In: Cordeiro, Z.J.M. (Ed.). Banana. Produção: aspectos técnicos. Embrapa Comunicação para Transferência de Tecnologia, Brasília, pp. 47-59.
Brasil, (1972) - Reconhecimento de solos do Estado da Paraíba. Ministério da agricultura. Levantamento exploratório. Rio de Janeiro: MA/EPE - SUDENE/DRN, (Boletim Técnico, 15), 683 pp.
Büll, L.T. (1993) - Nutrição mineral do milho. In: Bull, L.T. & Cantarella, H. Cultura do milho: fatores que afetam a produtividade. Potafos, Piracicaba, pp. 63-145.
Cayón Salinas, D.G. (2001) - Evolución de la fotosíntesis, transpiración y clorofila durante el desarrollo de la hoja de plátano (Musa AAB Simmonds). InfoMusa, Montpellier 10, 1: 12-15.
Coelho, E.F.; Ledo, C.A.S. & Silva, S.O. (2006) - Produtividade da bananeira ´Prata-Anã´ e ´Grande Naine´ no terceiro ciclo sob irrigação por microaspersão em tabuleiros costeiros da Bahia. Revista Brasileira de Fruticultura 28, 3: 435-438.
Cordeiro, Z.J.M. & Borges, A.L. (2000) - Problemas de causa abiótica e anormalidades de causa desconhecida. In: Cordeiro, Z.J.M. (Ed.) Banana. Fitossanidade. Embrapa Comunicação para Transferência de Tecnologia, Brasília, pp. 87-95.
Diniz, J.D.N.; Gonçalves, A.N.; Hernandez, F.F.F. & Torres, A.C. (1999) - Absorção de macronutrientes por explantes de bananeira in vitro. Pesquisa Agropecuária Brasileira 34, 7: 1201-1209.
Embrapa - Empresa Brasileira de Pesquisa Agropecuária. (1999) - Manual de análises químicas de solos, plantas e fertilizantes. 2.ª Ed. Centro Nacional de Pesquisa de Solos. Embrapa-Cnps, Rio de Janeiro, pp. 212.
Embrapa (2005) - Dia de campo na TV enfoca sigatoka-negra. Raiz e Fruto - Informativo da Embrapa Mandioca e Fruticultura Tropical, 18, 49: 4.
Fontes, P.S.F.; Carvalho, A.J.C.; Cereja, B.S.; Marinho, C.S. & Monnerat, P.H. (2003) - Avaliação do estado nutricional e do desenvolvimento da bananeira Prata-anã (Musa spp.) em função da adubação nitrogenada. Revista Brasileira de Fruticultura 25, 1: 156-159.
Hendry, G.A.F. & Price A.H. (1993) - Stress indicators: chlorophylls and carotenoids. In: Hendry, G.A.F. & Grime, J.P. (Eds.) Methods in comparative plant ecology. Chapman & Hall, London, pp.148-152
Katan, J.; Greenberger, A.; Alon, H. & Grinsten, A. (1976) - Solar heating by polyethylene mulching for the control of diseases caused by soil-borne pathogens. Phytopathology 66, 5: 683-688.
Lahav, E. (1995) - Banana nutrition. In: Gowen, S. (Ed.). Bananas and plantains. London, Chapman & Hall, pp. 258-316.
Lichtenthaler, H.K. (1987) - Clorophylls and carotenoids: pigments of photosynthetic biomembranes. Methods in Enzymology 148: 350-382.
Lima, N.P.; Biasi, L.A.; Zanette, F. & Nakashima, T. (2003) - Produção de mudas por estaquia de duas espécies de guaco. Horticultura Brasileira, Campinas 21, 1: 106-109.
Linden, M.M.G. van der. (1985) - Sobre a organização da produção da banana no município de Bananeiras (PB). Dissertação de mestrado, Universidade Federal da Paraíba, Campina Grande.
Marschner, H. (1995) - Mineral nutrition of higher plants. 2nd. ed. San Diego, USA, Academic Press, 1995.
Monteiro, F.A.; Dechen A.R. & Monteiro, Q.C. (1995) - Elementos essenciais - absorção, transporte, redistribuição e funções. Abeas, Brasilia, 58 pp. (Curso de Nutrição Mineral de Plantas, Módulo 1).
Moreira, R.S. (1999) - Banana: teoria e prática de cultivo. 2.ª Ed. Fundação Cargill, 1. São Paulo, Cd-Rom.
Moreira, A. & Fageria, N.K. (2008) - Potential of Brazilian Amazon soils for food and fiber productions. Dynamic Soil, Dynamic Plant, Tokyo 2, 2: 82-88.
Phillips, I.D.J. (1969) - Apical dominance. In: Wilkids, M.D. (Ed.) The physiology of growth and development. Mc Graw-Hill, London, 26: 341-367.
Pilbeam, D.J. & Kirkby, E.A. (1983) - The physiological role of boron in plants. Journal of Plant Nutrition 6: 56–82.
Raij, B.V. (1991) - Avaliação da Fertilidade do solo. Instituto da Potassa e do Fosfato, Piracicaba, 142 pp.
Ricci, M.S.F.; Almeida, D.L.; Ribeiro, R.L.D.; Aquino, A.M.; Pereira, J. C.; De-Polli, H.; Reis, V.M. & Eklund, C.R. (1999) - Cyperus rotundus control by solarization. Biological Agriculture and Horticulture, Husbandry 17: 151-157.
Ronchi, C.P.; Fontes, P.C.R.; Pereira, P.R.G.; Nunes, J.C.S.; Martinez, H.E.P. (2001) - Índices de nitrogênio e de crescimento do tomateiro em solo e em solução nutritiva. Revista Ceres 48, 278: 469-484. [ Links ]
Ruggiero, C. & Gottardi, M.V.C. (1997) - Avaliação do Método de Propagação Rápida da Bananeira, Vilamoura 18: 130-135. (2º Congresso Iberoamericano, 3º Congresso Ibérico de Ciências Hortícolas).
Santos, C.S. (2002) - Nitrogênio e Potássio aplicados via fertirrigação em banana Pacovan. Dissertação de mestrado, Universidade Federal da Paraíba, Areia, 76pp.
Silva, C.R.R.; Souto, R.F. & Menegucci, J.L.P. (1999) - Propagação da Bananeira. Informe Agropecuário, Belo Horizonte 20, 196: 16-20.
Silva, G.D. (2000) - Propagação da bananeira (Musa sp.) através de moforgênese indireta in vivo. Dissertação em doutoramento, Universidade Estadual de São Paulo, Jaboticabal, 74 pp.
Silva, G.D.; Perecin, D.; Goes, A. & Natale, W. (2000) - Método de propagação da bananeira (Musa spp.) através da eliminação da gema apical do rizoma obtida em primeira brotação. SBF, Fortaleza, pp. 122-123. (Congresso Brasileiro de Fruticultura, 16).
Soto Ballestero, M. (1992) - Bananas: cultivo y comercializacion. 2. ed. Litografia e Imprensa Lil, San José, Costa Rica, 649 pp.
Souto, R.F.; Rodrigues, M.G.V.; Ruggiero, C.; Menegucci, J.L.P. (1999) - Novas perspectivas em sistemas de implantação, condução e práticas de manejo da bananeira. Informe Agropecuário, Belo Horizonte 20, 196: 10-15.
Turner, D. (1994) - Bananas and plantains. In: Schaffer, B. & Andersen, P.C. (Eds.) Handbook of environmental physiology of fruit crops. Library of Congress, Massachusetts 2, pp. 37-66.
Recepção/Reception: 2008.10.07
Aceitação/Acception: 2010.04.12

