










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias v.34 n.1 Lisboa jan./jun. 2011
Seleção de genótipos de Brachiaria Ruziziensis quanto ao alumínio em solução nutritiva. II: Avaliação da tolerância ao alumínio
Paulo Sérgio Balbino Miguel1, Wadson Sebastião Duarte da Rocha2, Fausto Souza Sobrinho2, Carlos Eugênio Martins2, Fernando Teixeira Gomes3, André Vicente de Oliveira4 e Caio Antunes de Carvalho5
1Estudante de Mestrado da Universidade Federal de Viçosa – Departamento de Microbiologia Agrícola – bolsista da Fapemig; Avenida Peter Henry Rolfs, Campus Universitário, s/n, Centro – CEP 36570-000 – Viçosa/MG – Brasil. Paulo.b@ufv.br – (31) 98974905.
2 Pesquisadores – D. Sc. – Embrapa Gado de Leite – Rua Eugênio do Nascimento, 610 – Dom Bosco – CEP 36038-300 – Juiz de Fora/MG – Brasil. caeuma@cnpgl.embrapa.br
3Professor – D. Sc. – Centro de Ensino Superior de Juiz de Fora (CES-JF) – Rua Luz Interior, 345, Estrela Sul, 36030776, Juiz de Fora/MG; Brasil. ftgomes@bol.com.br.
4Aluno de graduação – Centro de Ensino Superior de Juiz de Fora - Bolsistas de Iniciação Científica do CNPq/CNPGL.
5Estudante de Mestrado da Universidade Federal de Viçosa – Departamento de Biologia Animal – bolsista do CNPq; Avenida Peter Henry Rolfs, Campus Universitário, s/n, Centro – CEP 36570-000 – Viçosa/MG – Brasil. caio.biologia@yahoo.com.br.
RESUMO
O alumínio, em solos ácidos, é um dos principais responsáveis pela baixa produtividade de culturas. O objetivo deste trabalho foi avaliar a variabilidade genética da Brachiaria ruziziensis para a tolerância ao alumínio. Foram utilizados 10 genótipos da espécie referida, crescidas em vasos com 0, 30 e 60 mg/L de alumínio em solução nutritiva. Foi utilizado o delineamento inteiramente casualizado em esquema fatorial, com seis repetições e parcelas de uma planta por vaso. Foram avaliados: a produção de massa verde da parte aérea e raízes (MVPA e MVR), a produção de matéria seca da parte aérea e raízes (MSPA e MSR), o incremento no crescimento da parte aérea e das raízes (IPA e IR), expresso pela diferença entre o crescimento final e o inicial, e o incremento no número de perfilhos (INP). O aumento nas concentrações de alumínio provocou decréscimos para as médias de todas as características avaliadas, excetuando o IR. A ausência de diferenças do genótipo 8 para seis das sete características analisadas, com exceção da MVPA, demonstra ser ela a mais promissora para estudos posteriores em melhoramento, visando maior produtividade em solos ácidos. Os resultados evidenciaram a existência de variabilidade genética entre os genótipos avaliados para a tolerância ao alumínio tóxico, sendo possível identificar materiais mais produtivos na presença desse metal.
Palavras-chave: Acidez, alumínio tóxico, braquiária, produção de forragem, solução nutritiva.
Brachiaria Ruziziensis genotypes selection. II: Evaluation of the aluminum tolerance
ABSTRACT
In acid soils, the aluminum is responsible for the low cultures productivity. The objective of this work was to evaluate the Brachiaria ruziziensis genetic variability for the aluminum tolerance.10 genotypes of that species were used, grown in vases with 0, 30 and 60 mg/L of aluminum in nutritious solution. A completely randomized design under 10 x 3 factorial, with six replicates. The following variables were evaluated: the green mass production of the aerial part and roots (MVPA and MVR), the dry matter production of the aerial part and roots (MSPA and MSR), the increment in the growth of the aerial part and of the roots (IPA and IR), expressed by the difference among the final growth and the initial, and the increment in the numbers of tillers (INP). The increase in the aluminum concentrations decreases for the averages of all the characteristics evaluated, excepting IR. The absence of differences of the genotype 8 for six characteristics, except for MVPA, demonstrates to be most promising for subsequent genetic breeding studies for larger productivity of the cultures in acid soils. The results evidenced the existence of genetic variability among the genotypes for the tolerance to the aluminum, being possible to identify more productive materials in the presence of that metal.
Keywords: Acidity, forageproduction, nutritious, signal grass, solution, toxicity aluminum.
INTRODUÇÃO
Os solos brasileiros, em sua maioria, apresentam baixa fertilidade, acidez elevada e altas concentrações de alumínio tóxico, fatores que limitam o crescimento das plantas. A presença do Al reduz o crescimento e o desenvolvimento das raízes e diminui a absorção de nutrientes, o que é negativo para o desenvolvimento de vegetais sensíveis a este elemento (Abreu Jr. et al., 2003). Isto afeta a produção agrícola que para obter altos rendimentos necessita de substratos que possibilitem o desenvolvimento das raízes sem obstáculos químicos e/ou físicos (Echart e Cavalli-Molina, 2001).
Algumas espécies possuem a capacidade de tolerar altas concentrações deste metal, devido a sua complexação com ácidos orgânicos exsudados pelo sistema radicular e ao genótipo vegetal que pode promover a capacidade de adequar a condições físico-químicas adversas, minimizando os problemas causados pela baixa produtividade em solos ácidos (Freitas et al., 2006; Nolla et al., 2007).
O comportamento das espécies e variedades sob efeito do alumínio é diferente. Por isso, a seleção de plantas que suportam diferentes concentrações de alumínio é considerada indispensável em programas de melhoramento genético, que visam à identificação de genótipos mais produtivos, com maior adaptabilidade em condições de estresse (Freitas et al., 2006). Reforçando essa tendência, Kim et al. (2001) apontam o desenvolvimento de cultivares tolerantes ao alumínio em solos ácidos como uma das soluções econômicas a fim de aumentar a produção mundial de alimentos. Dentre as espécies tolerantes está a Brachiaria sp., que é muito utilizada para a alimentação animal nas regiões tropicais.
A Brachiaria ruziziensis Germ. & C.M.Evrard é uma planta perene, que atinge de 1 a 1,5 m de altura, com rizomas curtos e perfilhamento intenso. Além disso, apresenta grande quantidade de pequenos tricomas, alta produção de sementes, além de baixa a média exigência nutricional (Pupo, 1979). Com bons índices de produção, boa palatabilidade e aceitação pelo animal, além de suportar bem o pastejo (Alcântara, 1988). O seu cultivo tem despertado o interesse dos agricultores, por ser uma planta que pode produzir grande volume de matéria seca, com grande importância para o cultivo em plantio direto. Apesar disso, sofre com os problemas de toxicidade em solos ácidos e de baixa fertilidade (Correa, 2002), o que restringe o seu cultivo pelos produtores, tornando importante a seleção de progênies tolerantes a estes tipos de solo. Assim, o objetivo deste experimento foi avaliar a variabilidade genética da Brachiaria ruziziensis para a tolerância ao alumínio.
MATERIAL E MÉTODOS
O experimento foi conduzido em casa de vegetação na Embrapa Gado de Leite, situada no município de Juiz de Fora, Minas Gerais, Brasil. Utilizaram-se dez genótipos de Brachiaria ruziziensis (P1, P2, P3, P4, P5, P6, P7, P8, P9, P10) que fazem parte do programa de melhoramento genético desta espécie, conduzido na Embrapa Gado de Leite. O experimento foi realizado em um delineamento inteiramente casualizado, utilizando-se de um esquema fatorial 10 x 3, com seis repetições e parcelas compostas por uma planta/vaso.
As sementes dos genótipos foram colocadas para germinar em caixas plásticas contendo substrato comercial à base de casca de Pinus. Cerca de 60 dias depois, as mudas, após lavagem do sistema radicular, foram transplantadas para vasos contendo dois litros de solução nutritiva de Clark (1975) modificada em relação à concentração de fósforo (0,069 mol/L) com o objetivo de evitar a sua precipitação pelo alumínio. Nesse momento foi realizada a mensuração dos comprimentos iniciais da parte aérea (CIPA) e das raízes (CIR) de cada planta. Os vasos, mantidos em aeração constante, foram revestidos internamente com filme plástico de polietileno de cor preta e externamente por tinta preta betuminosa e alumínica, a fim de evitar a passagem de luz para o não crescimento de algas (Camargo e Camargo, 1985).
Na primeira semana, para adaptação dos genótipos ao ambiente aquático e uniformização dos materiais, as plantas foram submetidas à solução com metade da concentração recomendada e na segunda foi utilizada solução completa, mas sem Al (AlCl3.6H2O). Após este período, semanalmente foi realizada a troca da solução nutritiva, com a inclusão do alumínio nas diferentes doses, 0 (controle); 30; 60 mg/L. A cada troca semanal de solução o pH foi ajustado para 4,5 ± 0,1, utilizando NaOH ou HCl 0,5 mol/L, com pHgâmetro digital, não havendo necessidade de leitura diária do pH. Isto porque Martins et al. (2010) verificaram não ser necessária a correção do valor de pH da solução nutritiva para esta espécie, resultado este de muita importância e praticidade uma vez que os ajustes diários, além de aumentarem o custo os experimentos, principalmente quando o número de unidades experimentais é maior, causa estresse às plantas.
Após 21 dias de exposição ao alumínio as plantas foram retiradas da solução nutritiva, sendo seccionado o sistema radicular da parte aérea, realizando a medição do comprimento final da raiz (CFSR) e parte aérea (CFPA). Com o auxílio de uma balança analítica foi obtida a produção de massa verde da parte aérea (MVPA) e das raízes (MVR). Posteriormente, o material foi colocado em sacos de papel para secagem em estufa de circulação de ar durante 96 horas a 55°C para determinação da matéria seca de cada uma das partes da planta. Pela associação da MVPA e MVR e suas respectivas porcentagens de matéria seca, foram obtidas as estimativas da produção de matéria seca da parte aérea (MSPA) e das raízes (MSR). Depois de conhecidos os comprimentos inicial e final da parte aérea e raízes foi determinado o incremento, diferença entre os comprimentos inicial e final de raízes e parte aérea, identificados como IPA e IR. No início da fase experimental e na colheita, foi determinado o número de perfilhos (NP) de cada planta, sendo determinado por diferença o incremento do número de perfilhos (INP). Os dados foram submetidos à análise de variância, e as médias, quando necessário, foram comparadas pelo teste de Scott e Knot (1974), a 5% de probabilidade.
RESULTADOS E DISCUSSÃO
Foi verificada a existência de variabilidade genética entre os genótipos de B. ruziziensis para a tolerância ao alumínio (Quadro 1). O desenvolvimento das braquiárias também foi afetado pela presença de alumínio na solução (Quadro 1).
Quadro 1 - Valores médios de incremento no crescimento de parte aérea (IPA) e de raízes (IR), incremento no número de perfilhos (INP), massa verde da parte aérea (MVPA) e raízes (MVR), matéria seca da parte aérea (MSPA) e raízes (MSR), de 10 genótipos de B. ruziziensis, em três concentrações de alumínio.
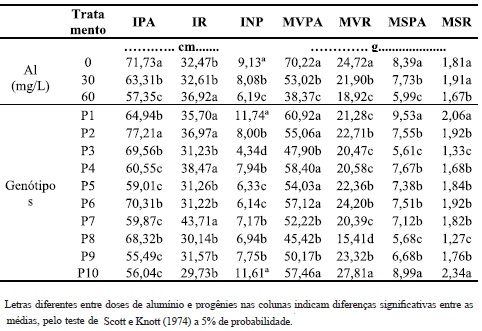
O aumento na concentração de alumínio em solução nutritiva provocou decréscimos nos valores médios de IPA, INP, MVPA e MVR e MSPA e MSR. No entanto, o incremento de crescimento do sistema radicular (IR) foi maior nas plantas cultivadas nas soluções com as doses mais altas de alumínio (Quadro 1). Portanto, quanto maior a concentração de alumínio na solução mais prejudicado será o desenvolvimento das plantas de B. ruziziensis. A dose de 30 mg/L foi crítica para os genótipos utilizados, pois foi verificado que o melhor desempenho das plantas ocorreu quando não havia alumínio na solução. Isto ocorreu em cinco das sete características avaliadas (71 %, Quadro 1). Este comportamento não foi verificado para duas características relacionadas ao sistema radicular. Estes resultados revelam a inconstância em relação ao comportamento destas progênies para as características avaliadas. O resultado diferenciado das características relacionadas com as raízes de genótipos de B. ruziziensis também foi verificado no trabalho de Martins et al. (2010).
A maioria dos trabalhos realizados envolvendo acidez tanto em campo quanto em solução nutritiva demonstram que o sistema radicular é a característica mais afetada pelo alumínio (Howeler e Cavadid, 1976). Camargo et al. (1987) trabalhando com a seleção de cultivares de milho em solução nutritiva, Sanchez-Chacón et al. (2000) em aveia, Mistro et al. (2001) em trigo, encontraram redução no crescimento do sistema radicular na presença de alumínio. Todos esses trabalhos estão de acordo com os resultados observados no Quadro 2, evidenciando que houve redução na produtividade de massa verde e seca de raízes com o incremento do alumínio em solução. Porém, o mesmo não foi verificado para o IR, que mostrou desempenho contrário. Este resultado, aparentemente contraditório, é comum na literatura, como os relatados por Martins et al. (2010) em B. ruziziensis e Mattiello et al. (2008) em café arábica. Por sua vez, Veloso et al. (1995), em pimenteira, não encontraram diferenças significativas para o comprimento das raízes na presença de Al, mas na sua ausência o comprimento foi menor, corroborando parcialmente com os resultados deste experimento, no qual o maior crescimento foi na dose mais elevada de Al.
Quadro 2 - Incremento no crescimento de parte aérea (IPA) e das raízes (IR) e incremento no número de perfilhos (INP) de dez genótipos (Gn) de B. ruziziensis, em três concentrações de alumínio.
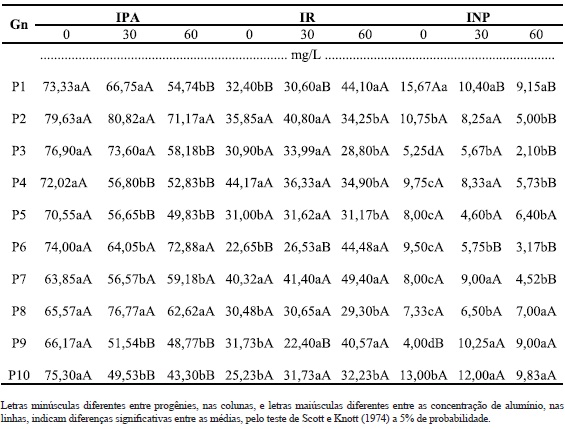
O maior crescimento do sistema radicular pode ser uma resposta imediata da planta ao estresse causado pelo alumínio. Nesse caso, a planta busca, por meio do maior crescimento das raízes, explorar maior volume para obter os nutrientes necessários ao seu desenvolvimento. Esse esforço se faz com perdas mais acentuadas na parte aérea das plantas, o que é comprovado pela maior redução percentual da parte aérea em relação ao sistema radicular. Considerando-se as doses 0 e 60 mg/L de alumínio, observou-se uma redução média de 28,6% na MSPA e de apenas 7,73% na MSR (Quadro 1). Como o período em que as plantas ficaram submetidas ao estresse foi curto (21 dias), não é possível comprovar essa teoria e saber quais as conseqüências futuras para as plantas. Entretanto, admite-se que este crescimento desproporcional entre raízes e parte aérea não é sustentável. Portanto, novos experimentos deverão ser conduzidos, com períodos de avaliação maiores.
O maior crescimento radicular verificado na dose 60 mg/L de Al (Quadro 1), não constitui em essencialidade deste elemento para a espécie. Este catião, provavelmente, acumulou-se na planta, já que ele tem sido detectado na parede celular, nos núcleos e nucléolos das células das raízes e em concentrações altas no citoplasma. A complexação do alumínio na raiz causa danos ao metabolismo do fósforo, retendo-o no órgão de absorção e reduzindo a disponibilidade do mesmo para assimilação, transporte e translocação de nutrientes para caules e folhas (Diniz e Calbo., 1990; Echart e Cavalli-Molina., 2001; Abichequer et al., 2003; Mattiello et al., 2008). Tal fato pode explicar os decréscimos nos incrementos de crescimento da parte aérea das plantas de B. ruziziensis, além de ser um fator importante nos mecanismos de tolerância. Simões et al. (1973), ao trabalharem com pinheiro-do-Paraná (Araucaria angustifolia), demonstraram existir uma tendência nítida para o decréscimo da parte aérea com o aumento nas doses de Al em solução, o que também foi verificado neste experimento (Quadro 1). Em cana-de-açúcar o alumínio afeta o desenvolvimento das raízes, bem como inibe o desenvolvimento dos demais órgãos vegetais, por prejudicar a absorção e o transporte de água pela planta, comprometendo a absorção, transporte e utilização de nutrientes (Drummond et al., 2001). Assim, existe uma relação da toxicidade do Al no comprimento das raízes com reações expressas na parte aérea (Crestani et al., 2009).
Mattiello et al. (2008) verificaram em dois cultivares de café maiores teores de alumínio nas raízes quando comparados às folhas e caules, não ressaltando diferenças no crescimento radicular entre os materiais genéticos avaliados. Apesar do alumínio não ter afetado de forma negativa, como era esperado, de acordo com a maioria dos trabalhos publicados, foi verificado que elas apresentavam-se grossas, quebradiças e pouco volumosas nas concentrações mais altas de alumínio (30 e 60 mg/L). É importante enfatizar que estas plantas, quando cultivadas em hidroponia, não apresentam obstáculos físicos para o seu desenvolvimento, as quais poderiam apresentar dificuldades de crescimento no solo, pela sua maior fragilidade.
O aumento nas doses de Al em solução nutritiva foi seguido pelo decréscimo (p < 5%) no número de perfilhos. As características IPA, INP, MVPA, MVR e MSPA apresentaram comportamento semelhante, uma vez que com a elevação nos níveis de Al observou-se o declínio nas suas médias (Quadro 1), verificando que o Al influencia no desenvolvimento dos genótipos de B. ruziziensis.
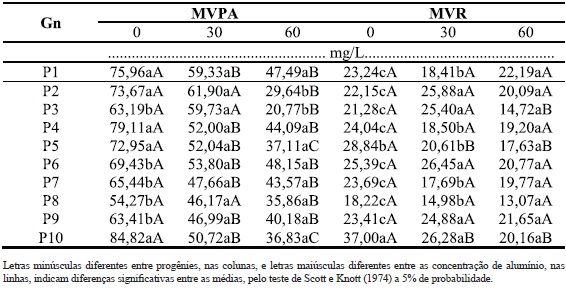
Os genótipos 1 e 10 foram os que apresentaram melhor desenvolvimento para o maior número de características, sendo cinco em cada sete (71%). No caso do genótipo 1, destacam-se o IR, INP, MVPA, MSPA, MSR. No caso do genótipo 10, destacam-se o INP, MVPA, MVR, MSPA e MSR. Além disto, apresentaram uma das maiores médias de matéria seca das raízes e parte aérea quando cultivada em 60 mg/L de Al (Quadro 4). Freitas et al.(2006) sugerem que algumas plantas, quando cultivadas em ambiente com alumínio, apresentam acréscimo na assimilação de nutrientes, uma vez que estes genótipos são beneficiados por pequenas doses de Al na translocação de fósforo, o que aumenta a absorção do mesmo. Portanto, a maior produção de matéria seca das raízes e parte aérea em alguns genótipos de B. ruziziensis podem estar associadas à assimilação de algum nutriente quando na presença de Al. Apesar disso, não se trata dos genótipos mais tolerantes ao Al, mesmo com características favoráveis para: INP, MVPA, MVR, MSPA e MSR em 60 mg/L. Quando comparadas com o genótipo 8, verifica-se que esta última apresentou menor variação entre as doses para o maior número de características tais como: IPA, IR, INP, MVR, MSPA e MSR e (Quadros 2, 3 e 4), ou seja, seis em sete características. Apesar de se tratar de um material estável, uma vez que apresenta pouca variação entre a presença e ausência de alumínio, suas médias nem sempre são altas, o que não lhe confere qualidades para algumas características tais como MVR, MSPA e MSR (Quadros 2, 3 e 4).
Quadro 3 - Massa verde da parte aérea (MVPA) e raízes (MVR), de dez genótipos (Gn) de B. ruziziensis, em três concentrações de alumínio.
Quadro 4 - Matéria seca da parte aérea (MSPA) e raízes (MSR), de dez genótipos (Gn) de B. ruziziensis, em três concentrações de alumínio.
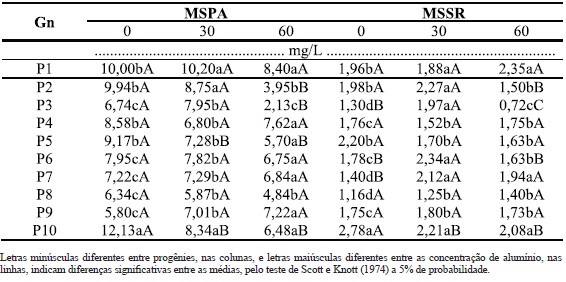
Ao analisar os desdobramentos das interações, os genótipos 2, 4, 5, 7, 8 e 10 foram as que obtiveram melhor comportamento nas doses analisadas, para a maioria das características. Sendo que, P7 e P8 obtiveram algumas semelhanças, diferindo nas características INP na dose 60 mg/L, MVPA na dose 30 mg/L e MSSR na dose 0 mg/L, com diferenças favoráveis para o P8, na emissão de perfilhos, característica de relevância na busca por plantas mais tolerantes, uma vez que um dos sintomas de toxicidade ao alumínio é o reduzido perfilhamento (Camargo e Camargo, 1985; Camargo et al., 2000). A ausência de diferenças para a progênie 8 para seis das sete características analisadas, excetuando a MVPA, demonstrou que ela possui o maior potencial para adaptação a solos ácidos, com tolerância para as doses de Al testadas. O P2, por sua vez, tolerou o Al na dose 30 mg/L para todas as características mensuradas e também a dose 60 mg/L para IPA, IR e MVR.
Os genótipos P5 e P10 também demonstraram comportamento semelhante quando comparados IPA, IR, INP, MVPA, MVR e MSPA, exceto para MSR, no qual P5, apesar de produzir menos matéria seca, obteve maior tolerância a presença do Al em solução, pela menor variação (amplitude) nas quantidades de matéria seca das raízes. Embora as demais características não tenham apresentado diferenças significativas em relação ao aumento das dosagens, P10 perfilhou melhor nas duas doses com maior produção de matéria seca da parte aérea em 30 mg/L. Algumas plantas, como é o caso de P3, apresentam tolerância somente até 30 mg/L, para todas os caracteres mensurados.
A concentração de 60 mg/L de alumínio foi a que proporcionou melhor discriminação entre plantas tolerantes e sensíveis (Quadro 1), por apresentar maiores diferenças significativas entre os parâmetros testados, corroborando com os resultados de Braccini et al. (2000). Para esses autores a concentração de 45 mg/L de Al, foi adequada para detectar diferenças entre linhagens de cafeeiro quanto à tolerância ao Al. Sanchez-Chacon et al. (2000), trabalhando com aveia, também encontraram uma dose (20 g kJ-1) que melhor demonstrou as diferenças entre os genótipos.
CONCLUSÕES
O estudo efectuado demonstrou que o genótipo 8 de Brachiaria ruziziensis foi o que apresentou as melhores condições para adaptação a solos ácidos, sendo este o mais promissor para a utilização em estudos complementares em condições de campo.
As diferenças significativas entre os genótipos analisados demonstram a presença de variabilidade genética em B. ruziziensis.
A dose de 60 mg/L de alumínio possibilitou maior discriminação entre plantas tolerantes e sensíveis ao Al disponível.
REFERÊNCIAS BIBLIOGRÁFICAS
Abreu Jr,C.H.; Muraoka, T. e Lavorante, A.F. (2003) - Relationship between acidity and chemical properties of Brazilian soils. Scientia Agricola, 60, 2/3: 337-343. [ Links ]
Abichequer, A.D.; Bohnen, H. e Anghinoni, I. (2003) - Absorção, translocação e utilização de fósforo por variedades de trigo submetidas à toxidez de alumínio. Revista Brasileira de Ciências do Solo, 27, 2/6: 373-378. [ Links ]
Alcântara, P.B. (1998) - Plantas forrageiras: gramíneas e leguminosas. São Paulo, Nobel,162 p. [ Links ]
Braccini, M.D.L.; Martinez, H.E.P. e Braccini, A.L. (2000) - Avaliação de linhagens de cafeeiros quanto à tolerância ao alumínio pelo método do papel-solução. Bragantia, 59, 2/4: 221-226. [ Links ]
Camargo, C.E.O. e Camargo, O.B.A. (1985) - Estudo comparativo da tolerância à toxicidade de Ferro e Alumínio em arroz. Bragantia,44, 1/4: 97-114. [ Links ]
Camargo, C.E.O.; Camargo, O.B.A. e Rocha-Júnior, L. S. (1987) - Trigo: Tolerância ao alumínio em solução nutritiva. Bragantia, 42, 2/4: 183-190. [ Links ]
Camargo, C.E. de O.; Ferreira Filho, A.W.P e Felicio, J.C. (2000) - Herança da tolerância ao alumínio em populações híbridas de trigo. Pesquisa Agropecuária Brasileira, 35, 3/12: 517-522. [ Links ]
Clark, R.B. (1975) - Caracterization of phosphatase in intact maize roots. Journal of Agricultural and Food Chemistry, 23, 3/4: 458-460. [ Links ]
Correa, L.A. (2002) – Características agronômicas das principais plantas forrageiras tropicais. Embrapa, 35, 1/1: 1-5. [ Links ]
Crestani, M.; Carvalho, F.I.F.; Oliveira, C.O.; Silva, J.A.G.; Souza, V.Q.; Parachu, E. A.M.; Silveira, G. ; Ribeiro, G. e Luche, H.S. (2009) - Estresse por alumínio em genótipos de aveia preta em condição hidropônica. Bragantia, 68, 3/4: 639-649. [ Links ]
Diniz, V.P.M. e Calbo, M.E.R. - Efeito da aplicação foliar de fósforo sobre a toxidez de alumínio em plantas de tomate. Revista Brasileira de Fisiologia Vegetal, 2, 2/3: 57-61. [ Links ]
Drummond, R.D.; Guimarães, C.T.; Feliz, J.; Ninamanfo-Cardenas, F.E; Carneiro, N.P.; Paiva, E. e Menossi, M. (2001) - Prospecting sugarcane genes involved in aluminum tolerance. Genetics and Molecular Biology, 24, 1/4: 221–230. [ Links ]
Echart, C.L. e Cavalli-Molina, S. (2001) - Fitotoxicidade do alumínio: efeitos, mecanismo de tolerância e seu controle genético. Revista Ciência Rural, 31, 3/4; 531-541. [ Links ]
Freitas, F.A.; Kopp, M.M.; Sousa, R.O.; Zimmer, P.D.; Carvalho, F.I.F. e Oliveira, A.C. (2006) - Absorção de P, Mg, Ca e K e tolerância de genótipos de arroz submetidos a estresse por alumínio em sistemas hidropônicos.Revista Ciência Rural, 36, 1/6: 72-79. [ Links ]
Howeler, R.H. e Cavadid, L.F. (1976) - Screening of rice cultivars for tolerance to Al-toxicity in nutrients solutions as compared with a field screening method. Agronomy Journal, 68, 4/6: 551-555. [ Links ]
Kim, B.Y.; Baier, A.C.; Somers, D.J. e Gustafson, J.P. (2001) - Aluminum tolerance in triticale, wheat and rye. Euphytica, 120, 3/3: 329-337. [ Links ]
Martins, C.E.; Miguel, P.S.B.; Rocha, W.S.D.; Souza Sobrinho, F.L.; Gomes, F.T. e Oliveira, A.V. (2010) - Seleção de genótipos de Brachiaria ruziziensis quanto à tolerância ao alumínio em solução nutritiva. Revista de Ciências Agrárias, 34, 1: 154-162. [ Links ]
Mattiello, E.M.; Pereira, M.G.; Zonta, E.; Mauri, J.; Matiello, J. D.; Meireles, P. G. e Silva, I.R. (2008) - Produção de matéria seca, crescimento radicular e absorção de cálcio, fósforo e alumínio por Coffea canephora e Coffea arabica sob influência da atividade do alumínio em solução. Revista Brasileira de Ciência do Solo, 32, 1/6: 425-434. [ Links ]
Mistro, J.C.; Camargo, C.E.O. e Pettinelli-Júnior, A. (2001) - Avaliação de genótipos de trigo, de diferentes origens, em relação à toxicidade de alumínio. Bragantia, 60, 3/6: 177-184. [ Links ]
Nolla, A.; Schlindwein, J.A. e Anghinoni, I. (2007) - Crescimento, morfologia radicular e liberação de compostos orgânicos por plântulas de soja em função da atividade de alumínio na solução do solo de campo natural. Revista Ciência Rural, 37, 1/6: 97-101. [ Links ]
Paterniani, M.E.A.G. e Furlani, P.R. (2002) - Tolerância à toxicidade de alumínio de linhagens e híbridos de milho em solução nutritiva. Bragantia, 61, 1/4: 11-16. [ Links ]
Pupo, N.I.H. (1979) - Manual de pastagens e forrageiras: formação, conservação e utilização. Campinas, Instituto Campineiro de Ensino Agrícola, 341 p. [ Links ]
Sanchez-Chacon, C.D.; Federizzi, L.C.; Milach, S.C.K. e Pacheco, M.T. (2000) - Variabilidade genética e herança da tolerância à toxicidade do alumínio em aveia. Pesquisa Agropecuária Brasileira, 35, 9/12: 1797-1808. [ Links ]
Scott, A.J. e Knot, M.A.A. (1974) – Cluster analysis method for grouping means in the analysis of variance. Biometrics, Arlington, 30, 3/4: 507-512. [ Links ]
Simões, J.W.; Couto, H.T.Z. e Kajyia, S. (1973) - Tolerância do pinheiro do paraná (Araucaria angustifolia (Bert.) O. Ktze) a teores crescentes de alumínio.IPEF, 6, 2/2: 93-102. [ Links ]
Veloso, C.A.C.; Muraoka, T.; Malavolta, E. e Carvalho, J.G. (1995) - Efeitos do alumínio em pimenteiras do reino (Piper nigrum L.) cultivadas em solução nutritiva.Revista Scientia Agricola, 52, 2/3: 368-375. [ Links ]
Recepção/Reception: 2010.06.23
Aceitação/Acception: 2011.03.05

