










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias vol.34 no.2 Lisboa jul. 2011
Actividades enzimáticas antioxidativas em Cistus Ladanifer L. provenientes de áreas não contaminadas em elementos vestigiais
Erika Santos1,2, Mª Manuela Abreu2, Jorge Saraiva3 e Cristina Nabais4
1Centro de Investigação em Ciências do Ambiente e Empresariais (CICAE), Instituto Superior Dom Afonso III, Convento Espírito Santo, 8100-641 Loulé - Portugal. erika.santos@inuaf-studia.pt;
2Unidade de Investigação Química Ambiental (UIQA), Instituto Superior de Agronomia, Universidade Técnica de Lisboa (TULisbon), Tapada da Ajuda, 1399-017 Lisboa – Portugal;
3Departmento de Química, Unidade de Investigação de Química Orgânica, Produtos Naturais e Agro-alimentares (QOPNA), Universidade de Aveiro, Campus Universitário de Santiago, 3810-193 Aveiro – Portugal;
4Centro de Ecologia Funcional, Departamento de Botânica, Universidade de Coimbra, Calçada Martim de Freitas, Arcos do Jardim, 3000 Coimbra - Portugal
RESUMO
Avaliou-se a actividade das enzimas antioxidativas (catalase, peroxidase e superóxido dismutase) e as concentrações em As, Cu, Pb e Zn nas folhas de duas populações de Cistus ladanifer L. colhidas, na Primavera e no Verão, em solos não contaminados em elementos vestigiais (Caldeirão e Pomarão) e com condições climáticas diferentes.
Nos solos das duas áreas, as concentrações totais e disponíveis (extracção com DTPA) em elementos vestigiais foram baixas. A distribuição dos elementos pelas folhas novas e maduras foi semelhante entre áreas e estações do ano. As actividades enzimáticas variaram consoante a população. As folhas colhidas nas duas estações do ano, excepto as folhas novas de Verão do Pomarão, apresentaram actividade enzimática na fracção solúvel e iónica. Comparando as estações do ano e áreas de amostragem, constatou-se que as actividades enzimáticas representam um mecanismo de tolerância a vários factores de stresse (radiação UV, temperatura do ar elevada no verão, défice hídrico no verão, baixa-média fertilidade do solo e conteúdo de elementos vestigiais no solo), o que confere uma elevada plasticidade à espécie.
Palavras-chave: Cistus ladanifer L., condições climáticas, elementos vestigiais, enzimas antioxidativas, solos não contaminados.
Antioxidant enzymes activity of Cistus Ladanifer L. from areas non contaminated in trace elements
ABSTRACT
The aim of this study was to evaluate the antioxidant enzymes activity (catalase, peroxidase and superoxide dismutase) and the concentrations of As, Cu, Pb and Zn in leaves from two populations of Cistus ladanifer growing on soils non-contaminated with trace elements (Caldeirão and Pomarão), but with different climatic conditions and in two different seasons (spring and summer).
In both areas, total and available concentrations (DTPA extraction) of trace elements in soils were low. The distribution of elements for young and mature leaves was similar between areas and seasons. The enzyme activities varied with the plant population. Leaves collected in the two seasons, except young leaves from Pomarão collected in summer, showed enzymatic activity in the soluble and ionic fractions. Comparing the seasons and sampling areas, C. ladanifer enzymatic activities represent a tolerance mechanism to different stress factors (UV radiation, high air temperature in summer, drought, low-medium fertility in soils, trace element concentrations in soil), giving it a high plasticity.
Key-words: Antioxidative enzymes, Cistus ladanifer L., climatic conditions, non-contaminated soils, trace elements.
INTRODUÇÃO
Cistus ladanifer L. (esteva) é um arbusto que cresce naturalmente no mediterrâneo. Esta espécie cresce em áreas sujeitas a diferentes condições de stresse tais como: elevada radiação solar e temperatura, baixos conteúdos de água e matéria orgânica no solo, elevadas concentrações de elementos vestigiais no solo e baixo pH.
Embora a "verdadeira" tolerância esteja associada a mecanismos fisiológicos controlados geneticamente, existem evidências de que alguns factores ambientais adversos ou induzidos pela acção do Homem podem provocar processos de evolução ou mudanças na adaptação das plantas (Schmid, 1992). A plasticidade existente na esteva pode estar relacionada com o genótipo e/ou condições ambientais (Núñez-Olivera et al., 1996). Assim, de modo a poderem colonizar e desenvolver-se em áreas degradadas e com variações climáticas sazonais, estas plantas desenvolveram uma série de mecanismos fisiológicos e morfológicos de tolerância. Dimorfismo foliar, variações nas concentrações de clorofilas e taxa fotossintética, regulação do ângulo foliar, translocação dos nutrientes para as folhas novas antes da senescência foliar, aumento da espessura da epiderme e da concentração de flavonóides são alguns exemplos de mecanismo de tolerância da esteva face às condições climáticas sazonais, stresse hídrico, térmico e luminoso (radiação UV) (Núñez-Olivera et al., 1996; Chaves et al., 1997; Correia, 2002).
Como consequência de flutuações significativas de intensidade e duração da exposição às condições de stresse ambiental, as plantas podem aumentar a concentração de espécies reactivas de oxigénio (ROS, reactive oxygen species) e ficarem sujeitas a stresse oxidativo. Quando as plantas estão sob este tipo de stresse, podem produzir ou estimular enzimas antioxidativas, como catalase (CAT), peroxidase (POD) e superóxido dismutase (SOD), e/ou componentes não-enzimáticos que eliminam e neutralizam ROS de modo a protegerem as células de potenciais danos (Alexieva et al., 2001; Pang et al., 2003, Santos et al., 2009).
A função da SOD é catalisar a transformação do radical superóxido gerado na planta em situações de stresse, o que leva à formação de H2O2, na remoção do qual estão envolvidas as enzimas CAT e POD (Cao et al., 2004). Portanto, o equilíbrio entre a actividade da SOD e da POD e/ou CAT nas células é considerado crucial para determinar a homeostasia do O2 e H2O2. Assim, considera-se que é importante estudar o efeito dos elementos químicos, em especial os vestigiais, e outros tipos de stresse no nível da actividade dessas enzimas antioxidativas, como parte da adaptação das plantas ao meio.
Algumas enzimas podem ocorrer em duas fracções activas diferentes consoante a sua localização celular: solúvel no meio aquoso da célula (Ingham et al., 1998) ou ligadas através de interacções electrostáticas à parede celular e a alguns organelos (Moulding et al., 1987; McDougall e Morrison, 1995). A diferenciação entre estas duas formas é baseada no processo de extracção (McDougall e Morrison, 1995). Assim, formas solúveis podem ser extraídas com um tampão de baixa força iónica, enquanto que as formas ionicamente ligadas são extraídas com um tampão de extracção com força iónica elevada contendo, normalmente, 1 M de NaCl ou KCl (Dunand et al., 2002). A diferente localização e actividade destas duas formas enzimáticas podem reflectir diferentes funções fisiológicas, mas é reduzido o conhecimento relativo a esta situação (Saraiva et al., 2007).
Sob condições de campo, as plantas estão sujeitas à co-existência de vários factores de stresse, porém pouco é sabido sobre as respostas enzimáticas aos mesmos. Assim, este estudo teve como objectivo comparar as actividades enzimáticas da catalase, peroxidase e superóxido dismutase de duas populações espontâneas de C. ladanifer colhidas em locais cujos solos não estão contaminados por elementos vestigiais, mas com condições climáticas relativamente diferentes e em duas estações do ano (Primavera e Verão).
MATERIAIS E MÉTODOS
Área de amostragem e materiais
Para este estudo foram seleccionadas duas áreas cujos solos não estão contaminados por elementos vestigiais: Pomarão e Serra do Caldeirão. O Pomarão localiza-se no Concelho de Mértola (SE de Portugal) na margem direita do rio Guadiana, tendo sido o porto a partir do qual o minério proveniente da área mineira de São Domingos era exportado. O local de amostragem situa-se aproximadamente a 2 km do porto do Pomarão e a 25 m da antiga linha-férrea, por onde era transportado o minério. Os solos nesta área apresentam espessura inferior a 15 cm, desenvolveram-se a partir de xistos incluídos na Formação de Mértola, do Grupo do Flysh do Baixo Alentejo (Oliveira et al., 1990) e são classificados como Litossolos (Cardoso, 1974). O clima, de acordo com a classificação de Thornthwaite, é semiárido, mesotérmico sem excesso de água e com pequena concentração estival de eficiência térmica. Caracteriza-se por verões longos, quentes e secos e por invernos moderadamente frios e húmidos (temperatura média diária do ar >17,5 ºC; SMN, 1974). A precipitação média anual (INMG, 1990) foi de 456 mm e ocorreu maioritariamente no inverno e de uma forma irregular. A insolação média anual foi superior a 3100 horas (SMN, 1974).
A Serra do Caldeirão (S de Portugal) é delimitada pelo Barrocal algarvio e as planícies do Baixo Alentejo. Os solos desta área, com espessura inferior a 20 cm, desenvolveram-se sobre o complexo de xistos argilosos e grauvaques incluídos na Formação de Mira, do Grupo do Flysh do Baixo Alentejo (Oliveira et al., 1992). A área de amostragem situa-se na aldeia de Barranco do Velho, Concelho de Loulé, e os solos são também classificados como Litossolos (Cardoso, 1974). Embora o clima desta área também seja considerado mediterrâneo, as temperaturas são mais amenas (15-17,5 ºC de temperatura média diária do ar; SMN, 1974), relativamente ao Pomarão, pois está situada a 475 m de altitude. A precipitação média anual (INMG, 1990) é de 991 mm e a insolação média anual pode variar entre 2600 a 2800 horas (SMN, 1974).
Em cada uma das áreas delimitaram-se três parcelas contíguas de 150 m2 onde se colheram amostras compósitas de solo (Primavera de 2005), até ao máximo de profundidade conseguida (<20 cm), e folhas novas e maduras de C. ladanifer, de 15 plantas por parcela, em duas estações do ano (Primavera e Verão de 2005). Ambas as áreas de amostragem estão orientadas a sul contudo, a área da Serra do Caldeirão apresenta coberto arbóreo esparso de Quercus suber L. (sobreiro).
Métodos
Após homogeneização e secagem à temperatura ambiente, os solos foram crivados. A fracção <2 mm do solo foi caracterizada física e quimicamente (Póvoas e Barral, 1992): pH em água na proporção 1:2,5 (m:v); análise granulométrica; C orgânico por oxidação por via húmida; capacidade de troca catiónica (CTC) e catiões de troca (método do acetato de amónio a pH 7); N total (método de Kjeldahl); P e K extraíveis (método de Egner-Riehm).
A análise química total de As, Cu, Pb e Zn nos solos (fracção <2 mm) foi realizada por análise instrumental por activação de neutrões (INAA) ou espectrometria de emissão óptica com plasma acoplado indutivamente (ICP-OES) após digestão ácida com HF+HClO4+HNO3+HCl (ActLabs, 2010). A fracção disponível (solúvel em água e a fracção associada ao complexo de troca do solo) destes mesmos elementos foi extraída com uma solução de ácido dietileno triamino pentacético - DTPA (0,005 mol/L DTPA + 0,1 mol/L trietanolamina - TEA + 0,01 mol/L CaCl2; Lindsay e Norvell, 1978) sendo posteriormente analisado o Cu, Pb e Zn por espectrofotometria de absorção atómica em chama (F-AAS) e em câmara de grafite (GF-AAS) e o As por geração de hidretos (GH-AAS).
O material vegetal de Cistus ladanifer (folhas novas e maduras) foi lavado várias vezes com água destilada à qual se seguiram duas passagens com água milli-Q, seco a 70 ºC e finamente moído. Posteriormente, foi analisado pelas mesmas técnicas que a fracção disponível dos elementos no solo (F-AAS, GF-AAS e GH-AAS) após extracção, através de digestão ácida com HNO3 concentrado, sob pressão durante 10 horas a 150 ºC. Para controlo analítico, usaram-se folhas de choupo (NCS DC 73350) como material de referência.
Para a obtenção dos extractos enzimáticos (fracção solúvel e fracção ligada ionicamente), realizou-se uma extracção sequencial baseada em Ingham et al. (1998) e Pang et al. (2003). Assim, para a extracção adicionou-se a 0,50 g de folhas, previamente liofilizadas e moídas finamente, 10 mL de tampão fosfato (pH 7,2) a 50 mM, contendo ácido etilenodiamino tetra-acético (EDTA) a 1 mM e 1 % (p/v) de polivinilpolipirrolidona insolúvel (PVPP). A extracção realizou-se durante 15 minutos a 4 ºC com agitação, sendo depois o homogeneizado centrifugado (10 minutos a 22000 g e 4 ºC). O sobrenadante resultante foi congelado com azoto líquido, sendo posteriormente utilizado nos ensaios de quantificação da actividade enzimática da catalase (CAT), peroxidase (POD) e superóxido dismutase (SOD) na fracção solúvel. Ao resíduo resultante da extracção da fracção solúvel adicionou-se 10 mL de tampão fosfato (pH 7,2) a 50 mM que continha 1 mM de EDTA, 1 % (p/v) PVPP e 1 M de NaCl. A extracção decorreu também durante 15 minutos a 4 ºC, a que se seguiu centrifugação durante 10 minutos a 22000 g e 4 ºC. O sobrenadante resultante foi congelado com azoto líquido, sendo utilizando posteriormente nos ensaios de quantificação da actividade das mesmas enzimas, na fracção iónica.
A quantificação da actividade da CAT foi realizada com base no método descrito por Chance e Maehly (1955) e Wong e Whitaker (2003). Soluções de peróxido de hidrogénio (H2O2) a 200 mM, tampão fosfato a 0,1 M e pH 7,0 e água Milli-Q foram previamente incubadas a 25 ºC. Em cuvetes de quartzo foram adicionados 2 ml de tampão fosfato, 50 a 150 µL de extracto enzimático, 150 µL de H2O2 e um volume de água Milli-Q de modo a perfazer 3 mL, sendo a reacção iniciada pela adição de H2O2. O consumo de H2O2 foi seguido a 240 nm (espectrofotómetro UV-VIS) durante 2 min. O declive da porção linear da curva que relaciona a absorvância com o tempo foi calculado (ΔAbs240 min-1) e utilizado para determinar a actividade da CAT (µmol H2O2 min-1 g-1 peso fresco), utilizando o coeficiente de extinção (e) do H2O2 de 36,0 M-1 cm-1 (Verma e Dubey, 2003).
A actividade da POD foi quantificada usando guaiacol como substrato, como descrito por Chance e Maehly (1955) e Yuan e Jiang (2003). Soluções de 135 mM de guaiacol, 0,1 M de tampão fosfato (pH 7,0), 200 mM de H2O2 e água Milli-Q foram previamente incubadas a 25 ºC. Em cuvetes de quartzo adicionaram-se 1,2 mL de tampão fosfato, 500-1000 μL de extracto enzimático, 30 µL de H2O2, 200 ml de guaiacol e água Milli-Q para completar 3 mL de volume, sendo a reacção iniciada pela adição do guaiacol. O aumento da absorvância foi seguido a 420 nm (espectrofotómetro UV-VIS) durante 5 min. O declive da porção linear da curva, que relaciona a absorvância com o tempo, foi calculado (ΔAbs420 min-1) e utilizado para determinar a actividade da POD (mmol de H2O2 consumidos min-1 g-1 peso fresco), utilizando o coeficiente de extinção (e) de 2,60 mM-1 cm-1 que corresponde ao produto de oxidação do guaiacol, tetraguaiacol (Verma e Dubey, 2003).
A actividade da SOD foi determinada de acordo com Sun e Zigman (1977) e Khopde et al. (2001). Soluções de 0,1 M de tampão hidrogenocarbonato de sódio (pH 10,0), 5 mM de epinefrina (pH 2,0) e água Milli-Q foram incubadas a 25 ºC. Em cuvetes de quartzo adicionaram-se 1,5 mL de tampão, 25-100 μL da amostra, 300 μL de epinefrina e água Milli-Q para completar 3 mL de volume, sendo a reacção iniciada pela adição de epinefrina. O aumento da absorvância foi seguido a 320 nm (espectrofotómetro UV-VIS) durante 1 minuto e a taxa de oxidação da epinefrina foi calculada pelo declive da porção linear da curva que relaciona a absorvância com o tempo (ΔAbs320 min-1). A taxa de auto-oxidação da epinefrina foi calculada nas mesmas condições, mas sem extracto enzimático. Uma unidade de SOD foi definida como a quantidade de enzima necessária para reduzir a taxa de auto-oxidação da epinefrina em 50 %.
Para todos os casos foram realizadas réplicas, excepto para os teores totais. A comparação dos parâmetros bioquímicos entre as duas áreas foi analisada por ANOVA e teste de Tukey (p<0,05), utilizando o programa estatístico SPSS v17.0. As correlações bivariadas de Pearson foram utilizadas para relacionar a influência de elementos vestigiais nas actividades enzimáticas das folhas.
RESULTADOS E DISCUSSÃO
As características dos solos amostrados constam do Quadro 1. Os solos de ambas as áreas apresentaram valores semelhantes de pH e textura franco-limosa. A capacidade de troca catiónica foi maior nos solos da Serra do Caldeirão, o que está de acordo com os valores do carbono orgânico (r=0,98). Em ambas as áreas, o Ca e o Mg foram os catiões de troca maioritários.
Quadro 1 – Caracterização química dos solos colhidos nas áreas do Caldeirão e Pomarão.
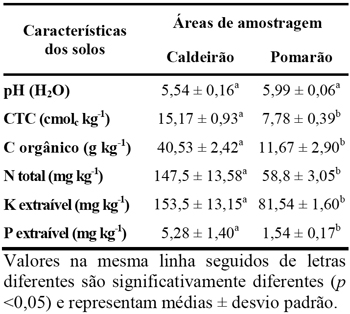
Relativamente à fertilidade, os solos na serra do Caldeirão apresentaram concentrações de C orgânico, N total e P e K extraíveis significativamente superiores às dos solos do Pomarão, o que pode estar relacionado com a maior cobertura de plantas existente nesses solos e às condições mais húmidas. De uma maneira geral, os solos do Caldeirão apresentaram fertilidade alta ou média-alta, embora o P extraível <10 mg/kg para ambas as áreas possa indicar fertilidade baixa. Os solos do Pomarão apresentaram fertilidade média ou baixa (Anónimo, 2000).
As concentrações em As, Cu, Pb, e Zn nos solos (total e da fracção disponível) e nas folhas de C. ladanifer colhidas no Caldeirão e Pomarão em ambas as estações do ano apresentam-se no Quadro 2. Os solos do Caldeirão e Pomarão não apresentaram diferenças significativas nas concentrações totais de todos os elementos vestigias. As concentrações totais de As nos solos de ambas as áreas e de Cu no Pomarão ultrapassaram os valores de referência da legislação canadiana para qualquer tipo de uso de solo (12 mg As kg-1, 63 e 91 mg Cu kg-1; CCME, 2007). Porém, as concentrações de todos os elementos químicos estudados, mesmo o As e Cu, apresentam valores considerados normais para as litologias a partir das quais os solos se desenvolveram (Quental et al., 2002; Abreu et al., 2004; Tavares et al., 2008) correspondendo por isso ao fundo geoquímico das áreas. Assim, os solos das áreas de estudo podem ser considerados como não contaminados nestes elementos vestigiais.
Quadro 2 – Concentrações em As, Cu, Pb e Zn nos solos (total e na fracção disponível) e nas folhas novas e maduras de Cistus ladanifer, colhidas na Primavera e Verão, no Caldeirão e Pomarão.
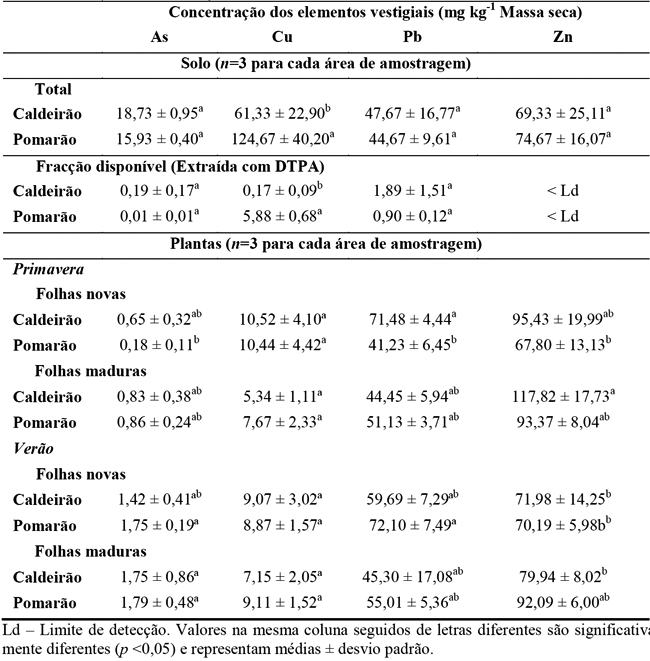
Relativamente à fracção disponível (extraída com DTPA), os solos de ambas as áreas não apresentaram diferenças significativas nas concentrações de As e Pb. A fracção de Cu disponível para os organismos nos solos do Pomarão foi superior, porém só representa 4,7 % da concentração total. Ambas as áreas apresentaram valores para a fracção de Zn disponível inferiores ao limite de detecção do aparelho analítico.
Embora as folhas maduras apresentem maiores concentrações em elementos vestigiais do que as folhas novas, esta variação não foi significativa. De uma maneira geral, as concentrações destes elementos nas folhas colhidas em ambas as estações do ano e áreas de amostragem foram semelhantes. Estes factos podem relacionar-se com as concentrações totais e da fracção disponível nos solos (excepto para o Cu) das duas áreas que são semelhantes. Em ambas as áreas, as concentrações de As, Cu e Zn nas folhas estiveram dentro da gama de valores considerada normal ou suficiente e abaixo dos valores fitotóxicos para as plantas no geral (Kabata Pendias e Pendias, 2001).
Apesar das concentrações de Cu nos solos (total e disponível) serem maiores no Pomarão, estes valores não reflectem o Cu nas folhas, uma vez que não foram observadas diferenças significativas para a concentração deste elemento entre as folhas, áreas estudadas e estações do ano. A translocação do Cu na planta é crucial pois este elemento desempenha um papel essencial nos processos fisiológicos da planta. Assim, sob condições de excesso, os tecidos das raízes podem demonstrar uma elevada capacidade para manter o nível de Cu em vez de o translocar para a parte aérea (Kabata Pendias e Pendias, 2001).
Apesar das baixas as concentrações de Pb no solo (Quadro 2), ambas as populações concentram quantidades consideradas fitotóxicas nas suas folhas (30-300 mg Pb kg-1; Kabata Pendias e Pendias, 2001), contudo não demonstraram sinais visíveis de toxicidade (folhas verdes muito escuras, murchamento das folhas maduras, folhagem atrofiada; Kabata Pendias e Pendias, 2001) sugerindo uma elevada tolerância da planta a este elemento.
Os valores obtidos para a actividade enzimática da CAT, POD e SOD nas folhas novas e maduras de C. ladanifer, colhidas na Primavera e Verão, nas duas áreas de amostragem constam da Figura 1.
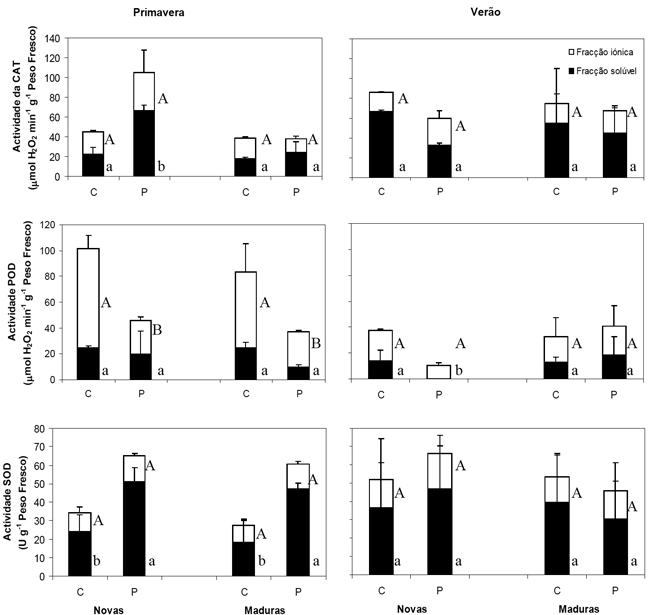
Figura 1 – Actividade das enzimas CAT, SOD e POD nas fracções solúvel e iónica em folhas de Cistus ladanifer, colhidas na Primavera e Verão, no Caldeirão (C) e Pomarão (P), (média±desvio padrão). Valores na mesma fracção seguidos de letras diferentes (maiúsculas para a fracção iónica e minúsculas para a fracção solúvel) são significativamente diferentes (p<0,05).
Em ambas as áreas e estações do ano, as folhas novas e maduras apresentaram actividade da CAT nas fracções solúvel e iónica sendo a fracção solúvel a que demonstrou maior percentagem da actividade total (50-78 %). Na Primavera, as folhas novas do Pomarão apresentaram uma actividade da CAT na fracção solúvel cerca de três vezes superior à das folhas maduras da mesma área e às folhas do Caldeirão, sugerindo que a actividade enzimática no Pomarão decorre também como mecanismo de tolerância ao stress hídrico, térmico e/ou à radiação solar nas folhas fotossinteticamente mais activas. O aumento da actividade total de CAT face ao stresse UV foi documentado por Yannarelli et al. (2006) em estudos feitos com Helianthus annuus. Em ensaios realizados com Pisum sativum L. e Triticum aestivum L. sujeitos a stresse hídrico e à radiação solar simultaneamente, a actividade de CAT total também aumentou relativamente ao controlo (Alexieva et al., 2001).
No Pomarão o decréscimo das actividades da CAT na fracção solúvel das folhas novas para as maduras na Primavera pode relacionar-se com o aumento das concentrações em Zn (r=-0,75) nas folhas, que apesar de não ocorrerem em concentrações consideradas tóxicas, estão muito próximas, nas folhas maduras, do valor mínimo da gama de fitotoxicidade (100 mg kg-1; Kabata-Pendias e Pendias, 2000). O decréscimo da actividade desta enzima face ao Zn foi também observado em Salix viminalis (Landberg e Greger, 2002) cultivado em hidroponia. As actividades da CAT na fracção iónica das folhas (novas e maduras) colhidas na Primavera não apresentaram diferenças significativas entre as áreas.
No Verão, as actividades da CAT nas fracções solúvel e iónica foram semelhantes entre as áreas e estádios de desenvolvimento das folhas. As actividades enzimáticas, na fracção solúvel das folhas de ambas as áreas e na fracção iónica das folhas do Pomarão, não parecem estar associadas a nenhum dos elementos químicos estudados. Porém, a actividade da fracção iónica nas folhas do Caldeirão pode ser estimulada caso as concentrações de As aumentem para níveis tóxicos (r=0,84). Comparando as duas estações do ano, constatou-se que as folhas de Verão demonstraram maior actividade da CAT na fracção solúvel que as da Primavera da mesma área (excepto folhas novas do Pomarão colhidas na Primavera), o que será devido, possivelmente, à intervenção da CAT como mecanismo de tolerância ao stresse hídrico e luminoso. O estímulo da actividade da CAT total face ao aumento da seca e radiação UV, em coexistência ou não, foi também observado para outras plantas (Alexieva et al., 2001; Yannarelli et al., 2006).
Relativamente à POD, a actividade da fracção iónica foi superior à da actividade da fracção solúvel em todos os casos, variando entre 54 e 100 % da actividade total. Na Primavera, as actividades da POD na fracção solúvel foram similares entre áreas e estádios de desenvolvimento das folhas, não estando correlacionadas com nenhum dos elementos estudados.
Relativamente à fracção iónica, verificaram-se diferenças significativas entre as áreas mas não entre as folhas novas e maduras da mesma área colhidas na Primavera. No Caldeirão, a actividade da POD na fracção iónica nas folhas pode relacionar-se com a concentração de Zn (r=-0,82) e/ou radiação UV existente. O estímulo da actividade da POD face ao aumento da radiação foi também verificado com Helianthus annuus L. e Pisum sativum L. (Alexieva et al., 2001; Yannarelli et al., 2006). Em contraste, no Pomarão a actividade da POD na fracção iónica parece não se relacionar com nenhum dos elementos químicos estudados. Assim, a existência de menores valores de POD nesta fracção no Pomarão, relativamente à do Caldeirão, pode relacionar-se com o stresse hídrico como reportado por Alexieva et al. (2001) em ensaios com Triticum aestivum L.
No Verão, a ausência de actividade da POD na fracção solúvel das folhas novas do Pomarão distinguiu-se significativamente e está, possivelmente, relacionada com o aumento da radiação UV absorvida pelas folhas de esteva e indisponibilidade de água. A similaridade entre as actividades da POD na fracção iónica de ambas as áreas de amostragem, os menores valores relativamente aos de Primavera e a não correlação com nenhum dos elementos químicos estudados, sugere que a actividade desta enzima poderá estar apenas associada ao aumento da temperatura e radiação. É de realçar que as folhas novas de Primavera apresentaram maior actividade de POD relativamente às de Verão o que pode sugerir ainda um acréscimo na defesa contra o stresse oxidativo, já que a planta está na fase de desenvolvimento e, consequentemente, fotossinteticamente mais activa.
Em ambas as áreas observou-se actividade da SOD nas fracções solúvel e iónica, independentemente do tipo de folha e estação do ano, sendo a fracção solúvel a que demonstrou maior percentagem da actividade total (57-78 %). Na Primavera, a actividade da SOD na fracção solúvel foi diferente entre as áreas, mas não entre tipo de folhas da mesma área. Porém, na fracção iónica a actividade da SOD foi semelhante entre as áreas e estádios de desenvolvimento das folhas.
Na Primavera, só as actividades da SOD na fracção solúvel e iónica do Caldeirão se relacionaram com um dos elementos vestigiais nas folhas (Zn: r=-0,84 e r=-0,76, respectivamente). As maiores actividades da SOD solúvel no Pomarão, relativamente ao Caldeirão, e a não relação com nenhum dos elementos vestigiais estudados pode sugerir a intervenção desta enzima contra o stresse UV e hídrico. Em ensaios com Triticum aestivum L e Pisum sativum L. sujeitas a stress UV e UV+hídrico também foi observado um aumento da actividade da SOD total (Alexieva et al., 2001).
No Verão, as actividades da SOD nas fracções solúvel e iónica foram semelhantes entre as áreas e tipos de folhas não se relacionando com nenhum dos elementos vestigiais estudados. Assim, esta enzima pode estar relacionada com a tolerância às condições de temperatura elevada, baixa humidade relativa do ar e com a maior incidência da radiação solar como observado em outros estudos (Alexieva et al., 2001). Para o Caldeirão verificou-se ainda uma inter-relação entre a actividade da CAT e SOD na fracção solúvel das folhas colhidas na Primavera (r=0,98) e na fracção iónica das folhas colhidas no Verão (r=0,83).
Para além das razões apresentadas, a variação das actividades enzimáticas observada nas folhas de C. ladanifer pode também ser devida ao défice/excesso de algum elemento químico não estudado, nomeadamente micronutrientes, ou à possível interacção de vários factores de stress.
CONCLUSÕES
Apesar de as concentrações de Pb no solo serem baixas ambas as populações apresentaram nas suas folhas quantidades deste elemento consideradas fitotóxicas para as plantas no geral. No entanto, não foram visíveis sinais de toxicidade. As plantas de Cistus ladanifer parecem estar bem adaptadas à coexistência de um vasto conjunto de factores de stresse ambiental (radiação UV, temperatura do ar elevada no verão, défice hídrico no verão, baixa-média fertilidade do solo e conteúdo de elementos vestigiais no solo) presentes nas áreas do Caldeirão e Pomarão. Esta tolerância pode estar relacionada com o funcionamento eficaz de diferentes isoenzimas de CAT, POD e SOD. A menor actividade das enzimas antioxidativas, nomeadamente da POD na fracção solúvel, pode ser compensada pela actividade de outras enzimas ou funcionamento de outros sistemas antioxidativos.
REFERÊNCIAS BIBLIOGRÁFICAS
Abreu, M.M.; Tavares, M.T.; Vairinho, M.; Joaquim, C. e Quental, L. (2004) – Geoquímica comparada dos solos da área mineira de São Domingos, Alentejo: Fundo geoquímico Versus zona de exploração. Revista de Ciências Agrárias, 27, 1: 301-313. [ Links ]
ActLabs (2010a) - Code 1H, Total Digestion, ICP, INAA (em linha). Ontario, Activation Laboratories Ltd., 1 p. (Acesso em 2010.05.21). Disponível em: < http://www.actlabs.com/page.aspx?page=506&app=226&cat1=549&tp=12&lk=no&menu=64&print=yes > [ Links ].
Anónimo (2000) - Manual de fertilização de culturas. Lisboa, Laboratório Químico Agrícola Rebelo da Silva, INIA, Ministério da Agricultura do Desenvolvimento Rural e das Pescas, 221 p.
Alexieva, V.; Sergier, I.; Mappelli, S. e Karanov, E. (2001) - The effect of drought and ultraviolet radition on growth and stress marked in pea and wheat. Plant Cell Environment, 24: 1337-1344. [ Links ]
Cao, X.; Ma, L.Q. e Tu, C. (2004) - Antioxidative responses to arsenic in the arsenic hyperaccumulator chinese brake fern (Pteris vittata L.). Environmental Pollution, 128: 317-325. [ Links ]
Cardoso, J. C. (1974) - A Classificação dos Solos de Portugal – Nova Versão. Boletim de Solos 17: 14-46. Lisboa, SROA, Secretaria de Estado da Agricultura. [ Links ]
CCME (2007) - Canadian soil quality guidelines for the protection of environmental and human health: summary tables (em linha). Winnipeg, Canada, Canada Council of Ministers of the Environment, 6 p. (Acesso em 2010.05.21). Disponível em < http://www.ccme.ca/assets/pdf/rev_soil_summary_tbl_7.0_e.pdf >.
Chance, B. e Maehly, A.C. (1955) - Assay of catalases and peroxidases. Methods in Enzymology, 2: 764-817. [ Links ]
Chaves, N.; Escudero, J.C. e Gutiérrez-Merino, C. (1997) - Quantitive variation of flavonoids among individuals of a Cistus ladanifer poulation. Biochemical Sytematics and Ecology, 25, 5: 429-435.
Clemens, S.; Bloss, T.; Vess, C.; Neumann, D.; Nies, D. H. e zur Nieden, U. (2002) - A transporter in the endoplasmic reticulum of Schizosaccharomyces pombe cells mediates zinc storage and differentially affects transition metal tolerance. Journal of Biological Chemistry, 277: 18215-18221. [ Links ]
Correia, O. (2002) - Os Cistus: as espécies do futuro?. In: Loução, K.A. (Eds) - Fragmentos de Ecologia. Lisboa, Escolar Editora, p. 97-119. [ Links ]
Dunand, C; Tognolli, M.; Overney, S.; von Tobel, L.; Meyer, M.; Simon, P. e Penel C. (2002) - Identification and characterisation of Ca2+-pectate binding peroxidases in Arabidopsis thaliana. Journal of Plant Physiology, 159, 11:1165-1171. [ Links ]
Grant, J.J. e Loake, G.J. (2000) - Role of active oxygen intermediates and cognate redox signaling in disease resistance. Plant Physiology, 124:21-29. [ Links ]
Ingham, L.M.; Parker, M.L. e Waldron, W. (1998) - Peroxidase: changes in soluble and bound forms during maturation and ripening of apples. Physiologia Plantarum, 102: 93-100. [ Links ]
INMG (1990) - O clima de Portugal. Normais climatológicas da região de "Alentejo e Algarve" correspondentes a 1951-1980. Lisboa, Instituto Nacional de Meteorologia e Geofísica, 98 p. (Fascículo XLIX, Vol 4 – 4ª Região). [ Links ]
Landberg, T. e Greger, M. (2002) - Differences in oxidative stress in heavy metal resistant and sensitive clones of Salix viminalis. Journal of Plant Physiology, 159: 69-75. [ Links ]
Lindsay, W.L. e Norvell, W.A. (1978) - Development of a DTPA soil test for zinc, iron, manganese and copper. Soil Science, 42: 421-428. [ Links ]
Kabata-Pendias A. e Pendias, H. (2001) - Trace Elements in Soils and Plants. 3rd ed. CRC Press, Boca Raton, 413 p. [ Links ]
Khopde, S.M.; Priyadarsini, K.I.; Mohan, H.; Gawandi, V.B.; Satav, J.G.; Yakhmi, J.V.; Banavaliker, M.M.; Biyani, M.K. e Mittal, J.P. (2001) - Characterizing the antioxidant activity of amla (Phyllanthus emblica) extract. Current Science, 81: 185-190. [ Links ]
McDougall, G.J. e Morrison, I.M. (1995) - Ionically-bound and covalently-bound wall peroxidases differ in their substrate specificity. Biochemical Society Transaction, 23: 150S. [ Links ]
Moulding, P.H.; Grant, H.F.; McLeilan, K.M. e Robinson, D.S. (1987) - Heat stability of soluble and ionically bound peroxidases extracted from apples. Internacional Journal of Food Science and Technology, 22: 391-391. [ Links ]
Núñez-Olivera, E.; Martínez-Abaigar, J. e Escudero, J.C. (1996) - Adaptability of leaves of Cistus ladanifer to widely varying environmental conditions. Functional Ecology, 10: 636-646.
Pang, J.; Chan, G.S.Y.; Zhang, J.; Liang, J. e Wong, M.H. (2003) - Physiological aspects of vertiver grass for rehabilitation in abandoned metalliferous mine wastes. Chemosphere, 52: 1559-1570. [ Links ]
Oliveira, J.T.; Brandão Silva, J.; Romão, J.A.; Carvalho, D.; Van den Boogaard, M. e Ribeiro, A. (1990). Carta Geológica de Portugal na escala de 1:50 000, Folha 46-D-Mértola. Lisboa, Serviços Geológicos de Portugal.
Oliveira, J.T.; Pereira, E.; Ramalho, M.; Antunes, M.T. e Monteiro, J.H. (1992) - 5ª Edição da Carta Geológica de Portugal na escala de 1:500 000. Lisboa, Serviços Geológicos de Portugal.
Póvoas, I. e Barral, M.F. (1992) - Métodos de análise de solos. Lisboa, Instituto de Investigação Científica Tropical, Ministério do Planeamento e da Administração do Território, 61 p. (Comunicações do IICT, Série de Ciências Agrárias, Nº 10). [ Links ]
Quental, L.; Bourguignon, A.; Sousa, A.J.; Batista, M.J.; Brito, M.G.; Tavares, T.; Abreu, M.M.; Vairinho, M. e Cottard, F. (2002) - MINEO Southern Europe environment test site., contamination impact mapping and modeling, Final Report. Lisboa, Instituto Geológico e Mineiro, 131 p. (IST-1999-10337). [ Links ]
Saraiva, J.; Nunes, C. e Coimbra, M. (2007) - Purification and characterization of olive (Olea europaea L.) peroxidase - Evidence for the occurrence of a pectin binding peroxidase. Food Chemistry, 101: 1571-1579. [ Links ]
Schmid, B. (1992) - Phenotypic variation in plants. Evolutionary Trends in Plants, 6: 45-60. [ Links ]
Sun, M. e Zigman, S. (1977) - An improved spectrophotometic assay for superoxide dismutase based on epinephrine autoxidation. Analytical Biochemistry, 90: 81-89. [ Links ]
Tavares, M.T.; Sousa, A.J. e Abreu, M.M. (2008) - Ordinary kriging and indicator kriging in the cartography of trace elements contamination in São Domingos mining site (Alentejo, Portugal). Journal of Geochemical Exploration, 98: 43-56. [ Links ]
Wong, D.W.S. e Whitaker, J.R. (2003) - Catalase. In: Whitaker, J. R.; Voragen, A. G. J. e Wong, D. W. S. (Eds) Handbook of Food Enzymology. New York, Marcel Dekker Inc., p. 389-401. [ Links ]
Yannarelli, G.; Gallego, S.M. e Tomaro, L.M. (2006) - Effect of UV-B radiation on the activity and isoforms of enzymes with peroxidase activity in sunflower cotyledons. Environmental and Experimental Botany, 56: 174-181. [ Links ]
Yruela, I. (2005) - Copper in Plants. Brazilian Journal of Plant Physiology, 17: 145-156. [ Links ]
Yuan e Jiang (2003) - Peroxidase. In: Whitaker, J.R.; Voragen, A.G.J.; Wong, D.W.S. (Eds) Handbook of Food Enzymology. New York, Marcel Dekker Inc., 389 p. [ Links ]

