










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias vol.35 no.2 Lisboa jul. 2012
Elaboração de um plano de amostragem para Cigarrinha-Verde (Homoptera: Cicadellidae) em vinha no Vale do Lima
Sampling plan for Green-Leafhoper (Homoptera: Cicadellidae) in vineyard at Lima Valley
Raúl Rodrigues[1], Jorge Pereira[2] , Isabel Mourão[1], Miguel Brito[1]e João Garrido[3]
[1] Centro de Investigação de Montanha (CIMO) - Escola Superior Agrária/IPVC. Refóios, 4990-706 Ponte de Lima. E-mail: raulrodrigues@esa.ipvc.pt
[2] Escola Superior Agrária/IPVC
[3] EVAG - Comissão de Viticultura da Região dos Vinhos Verdes, Arcos de Valdevez
RESUMO
Este trabalho teve como objetivo: estudar o padrão espacial das ninfas de cigarrinha-verde Empoasca vitis, através dos modelos da Lei da Potência de Taylor e da regressão Iwao, nas castas Arinto e Vinhão conduzidas nos modos de produção biológico e de produção integrada; e elaborar um plano de amostragem binomial (presença-ausência). As conclusões evidenciaram que; (i) a lei da potência de Taylor mostrou-se mais adequada para descrever o padrão espacial das cigarrinhas-verdes, que foi aleatório, não havendo diferenças estatisticamente significativas entre castas e modos de produção; (ii) a dimensão da amostra aumenta com o grau de precisão; (iii) é possível simplificar o plano de amostragem enumerativo; e (iv) a dimensão da amostra requerida para estimar densidades populacionais correspondentes ao nível económico de ataque proposto para Portugal, varia entre 30 a 57 folhas.
Palavras-chave : Amostragem binomial,dimensão da amostra, Empoasca vitis , padrão espacial.
ABSTRACT
The aims of this work were: to study the spatial patterns of nymphal stages of the green-leafhopper Empoasca vitis using the regression models of Taylor's Power Law and Iwao, in vineyard cvs. Arinto and Vinhão in organic farming and integrated production systems; to develop a binomial sampling (presence-absence). The results showed that: i) Taylor's power law was more appropriate to describe the spatial pattern of E. vitis and indicated a random pattern; no statistically differences were detected between varieties and production systems, ii) the sample size required increases with the degree of accuracy, iii) it is possible to simplify the enumerative sampling plan; and iv) the sample size required to estimate population densities of this pest corresponding to the economic thresholds, ranges from 30 to 57 leaves.
Keywords : Binomial sampling, Empoasca vitis , sample size, spatial pattern.
INTRODUÇÃO
A cigarrinha-verde (Homoptera: Cicadellidae) constitue um grupo de insetos picadores-sugadores, que afetam além da vinha, grande número de espécies selvagens e cultivadas. Atualmente, assiste-se à sua consolidação como praga em território nacional, sendo estes insetos considerados um novo grupo de inimigos-chave nas regiões de clima marcadamente mediterrânico, designadamente Douro e Alentejo (Freitas e Amaro, 2001; Quartau e Simões, 1995). Do complexo de espécies associadas ao ecossistema vinha em Portugal Continental, apenas as espécies Empoasca vitis Goethe e Jacobiasca lybica (Bergenin e Zanon) são consideradas pragas da vinha, ocorrendo a primeiranas regiões vitícolas do norte e centro do país e a segunda mais a sul (Quartau e Simões, 1995; Freitas e Amaro, 2001). O interesse prático do conhecimento do padrão espacial das populações de cigarrinha-verde reside na possibilidade de estabelecer sistemas de amostragem, os quais não serão apropriados a não ser que esta distribuição seja determinada no campo. Por sua vez, a determinação do tamanho da amostra é uma componente chave para a elaboração de planos de amostragem em proteção integrada das culturas, dependendo estes do grau de precisão requerido, resultando a sua dimensão do compromisso entre os objetivos de manter o mais baixo possível o esforço amostral requerido e a necessidade de considerar o número de unidades amostrais, que se enquadre satisfatoriamente com os requisitos do método estatístico adotado.
Este trabalho teve como objetivos: (i) estudar o padrão espacial das ninfas de cigarrinha-verde E. vitis pelos modelos da Lei da Potência de Taylor e do modelo de regressão Iwao, nas castas Arinto e Vinhão conduzidas nos modos de produção biológico (MPB) e de produção integrada (PRODI); e (ii) elaborar um plano de amostragem binomial (presença-ausência), em alternativa ao plano enumerativo.
MATERIAL E MÉTODOS
O trabalho foi realizado em duas vinhas, conduzidas em PRODI e em MPB, na Estação Vitivinícola Amândio Galhano da Comissão de Viticultura da Região dos Vinhos Verdes, localizada no concelho de Arcos de Valdevez (Lat: 41º4844 N e Long: 8º24343 W). As observações incidiram nas castas Arinto e Vinhão, em ambos os modos de produção, e decorreram entre os estados fenológicos cachos visíveis e início da queda da folha, no período compreendido entre 7 de Abril e 30 de Setembro de 2008. Em cada vinha, delimitou-se uma parcela por casta. Em cada parcela, a amostra consistiu no conjunto de 50 folhas escolhidas aleatoriamente com base em duas folhas por videira. Procedeu-se à observação direta das folhas, contando-se todas as formas móveis de ninfas de cigarrinha-verde existentes na página inferior das folhas.
Análise do padrão espacial
A determinação do padrão espacial de cigarrinha-verde foi feita através de dois modelos de regressão linear: a lei da potência de Taylor (Taylor, 1961); e o modelo da regressão de Iwao (Lloyd, 1967; Iwao, 1968). Tais modelos baseiam-se na relação existente entre a variância e a média amostral. Para ambos os modelos, o ajustamento dos dados foi feito através do coeficiente de determinação (r2).
O modelo de Taylor descreve a relação existente entre a média ( m ) e a variância ( S2 ) através de uma função exponencial (Equação1):

em que: A é um fator escalar, que depende do tamanho da amostra (Southwood e Henderson, 2000), correspondendo ao antilogarítmo de a, ou seja, 10ª; e b é o índice de dispersão. Quando o parâmetro b for significativamente superior, igual ou inferior à unidade, o padrão espacial diz-se que é, respetivamente, agregado, aleatório e regular ou uniforme (Taylor, 1961).
Os parâmetros A e b da lei da potência de Taylor foram estimados através da análise de regressão linear do logaritmo da variância em função do logaritmo da média, após transformação logarítmica (log10) (Equação 2):
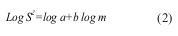
em que m é número médio de formas móveis de ninfas de cigarrinha-verde por folha; S2 a variância amostral; a a interseção com o eixo dos yy e b o declive da reta de regressão.
Para verificar se b diferiu significativamente da unidade, utilizou-se o teste t-Student (teste bilateral), dado pela Equação 3, aplicada a um nível de significância de n-2 graus de liberdade:
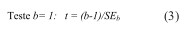
em que SEb é o erro padrão de b .
O modelo de regressão de Iwao quantifica a relação entre o agregado médio de Lloyd ( m* ) e a média. Para calcular os coeficientes de regressão de Iwao, estimou-se em primeiro lugar o agregado médio ( m* ) para cada amostragem, mediante a Equação 4:
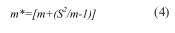
em que m e S2 representam respectivamente a média e a variância para o número médio de formas móveis de ninfas cigarrinhas-verdes por folha em cada data de amostragem (Lloyd, 1967). A relação entre m * e m foi feita por análise de regressão linear (Equação 5)

em que: 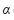 é a interseção com o eixo dos y e refere-se à média do número de indivíduos que vivem na mesma unidade amostral, sendo denominado índice básico de contágio do indivíduo (
é a interseção com o eixo dos y e refere-se à média do número de indivíduos que vivem na mesma unidade amostral, sendo denominado índice básico de contágio do indivíduo (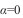 , o componente básico é o indivíduo;
, o componente básico é o indivíduo; 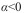 indica repulsão entre os indivíduos e
indica repulsão entre os indivíduos e 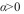 mostra que o componente básico é a colónia). O tamanho da colónia (agrupamento) é dado por
mostra que o componente básico é a colónia). O tamanho da colónia (agrupamento) é dado por 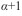 .
.
ß é o declive da reta, que indica se os agregados estão distribuídos de maneira uniforme ( ß <1), aleatória ( ß =1) ou agregada ( ß >1) (Iwao, 1968; Southwood e Henderson, 2000).
Para verificar se e ß eram significativamente diferentes de, respetivamente, zero e um, utilizou-se o teste t-Student (teste bilateral), dado pelas Equações 6 e 7 aplicadas a um nível de significância de n-2 graus de liberdade :
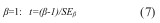
onde 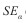 e
e 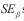 são o erro padrão de respectivamente e ß . Para valores de e ß =1 o padrão é considerado aleatório, pelo que segue a distribuição de Poisson.
são o erro padrão de respectivamente e ß . Para valores de e ß =1 o padrão é considerado aleatório, pelo que segue a distribuição de Poisson.
Para verificar se existiam diferenças entre os declives das retas de regressão obtidas a partir de ambos os modelos utilizados, utilizou-se o teste t-Student (teste bilateral) para o nível de significância de 5%.
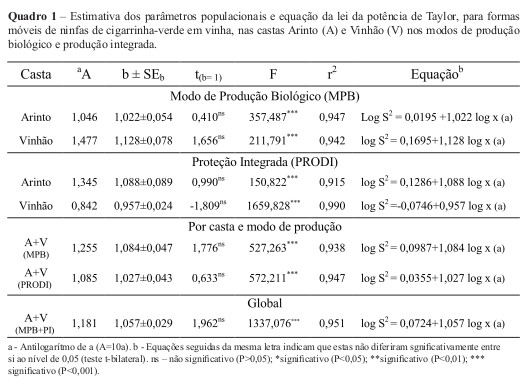
Relação entre a percentagem de folhas ocupadas e o número médio de formas móveis por folha
A determinação do número de formas móveis de cigarrinha-verde por folha (amostragem enumerativa) é um processo moroso e que se pode tornar impreciso para densidades elevadas, dada a forte mobilidade que o inseto apresenta. Por esta razão, estudou-se a possibilidade de realização de um método de amostragem presença/ausência (amostragem binomial), através do estudo da relação entre a percentagem de folhas ocupadas e o número médio de formas móveis de ninfas de cigarrinha-verde por folha, de forma a reduzir o custo da amostragem sem prejuízo para a fiabilidade e precisão do método.
Para tal, estudou-se a relação entre a percentagem de folhas ocupadas por pelo menos um indivíduo e o número de formas móveis por folha, através de regressão linear. O critério utilizado para determinar a validação do modelo foi o valor do coeficiente de determinação (r2), obtido através da regressão entre a percentagem de folhas ocupadas calculada e a percentagem de folhas ocupadas observada.
Determinação do tamanho da amostra
Amostragem enumerativa
O número de folhas necessário para estimar a população das ninfas de cigarrinha-verde, foi calculado através do modelo baseado no erro padrão da média (Equação 8) (Wilson e Room, 1983; 1980; 1994):
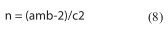
em que: n é o número folhas a amostrar, a e b são os parâmetros de Taylor , m o número médio de formas móveis de ninfas de cigarrinhas-verdes por folha e c o grau de precisão desejado (erro padrão da média, expresso como uma porção fixa da média). Os níveis de significância adotados foram de 15%, 20% e 25%.
Amostragem binomial ou presença/ausência
Uma alternativa ao método enumerativo e que visa maior economia de tempo nas observações das amostras, mantendo, no entanto, a precisão do método, consiste no recurso à amostragem binomial. Para tal, torna-se necessária a existência de uma correlação elevada entre a percentagem de folhas ocupadas calculada e a densidade populacional de um determinado organismo (Villagrán et al., 1999).
Assim, a partir dos resultados obtidos na amostragem enumerativa, fez-se a respetiva correspondência entre a densidade populacional de ninfas de cigarrinha-verde e a percentagem de folhas ocupadas por, pelo menos, uma forma móvel do fitófago.
RESULTADOS E DISCUSSÃO
Análise do padrão espacial
Lei da Potência de Taylor
Considerando a análise por casta e modo de produção, na vinha conduzida em MPB, a análise do conjunto de pares logaritmo da variância-logaritmo da média revelou ajustamentos significativos ao modelo de regressão para ambas as castas (Quadro 1), ou seja Arinto (94,7%) e Vinhão (94,2%). O índice de agregação de Taylor em ambas as castas, designadamente Arinto (b=1,022) e Vinhão (b=1,128), não diferiu significativamente da unidade, evidenciando que as ninfas de cigarrinha-verde se distribuíam de acordo com um padrão aleatório. O declive das retas de regressão não diferiu significativamente entre as casas em estudo (p>0,05). Na vinha conduzida em PRODI, o estudo da referida relação de pares, indicou ajustamentos significativos à regressão, com coeficientes de determinação de 91,5% na casta Arinto e de 99,1% na Vinhão (Quadro 1). O índice de agregação de Taylor em ambas as castas, designadamente Arinto (b=1,088) e Vinhão (b=0,957), não diferiu significativamente da unidade, evidenciando que as ninfas de cigarrinha-verde, se distribuíam de acordo com um padrão aleatório. O declive das retas de regressão não diferiu significativamente entre as casas em estudo (p>0,05) (Quadro 1).
De acordo com a análise conjunta para as castas em cada modo de produção, na vinha conduzida em MPB, a análise do conjunto de pares demonstrou ajustamentos significativos ao modelo de regressão (93,8%). O índice de agregação de Taylor para o conjunto Arinto+Vinhão (b=1,084) não diferiu significativamente da unidade, sugerindo que as ninfas de cigarrinha-verde, se distribuíram de acordo com um padrão aleatório. Na vinha conduzida em PRODI o conjunto de pares em análise proporcionou ajustamentos altamente significativos à regressão, expressos pelo coeficiente de determinação de 94,7%. O índice de agregação de Taylor para ambas as castas em conjunto (b=1,027) não diferiu significativamente da unidade, evidenciando que as ninfas de cigarrinha-verde se distribuíram de acordo com um padrão aleatório. O declive das retas de regressão não diferiu significativamente entre os modos de produção considerados (Quadro 1).
Na análise global dos dados para as castas e modo de produção, para os conjuntos modo de produção e casta, o estudo da relação de dependência logaritmo da variância-logaritmo da média para o conjunto de pares em análise, proporcionou ajustamentos significativos à regressão, expressos pelo coeficiente de determinação de 95,1%. O índice de agregação de Taylor em ambas as castas e modo de produção em conjunto (b=1,057), não diferiu significativamente da unidade, evidenciando que as ninfas de cigarrinha-verde se distribuíam de acordo com um padrão aleatório (Quadro 1).
Modelo de Regressão de Iwao
Na análise por casta e modo de produção, tanto na vinha conduzida em MPB como em PRODI, os pares agregado médio-média não se ajustaram significativamente ao modelo de Iwao, apresentando, em ambos os casos, coeficientes de determinação não significativos de 4,8% (Vinhão) e 18,4% (Arinto) na vinha conduzida em MPB e de 17,4% (Vinhão) e 10,7% (Arinto) na vinha conduzida em PRODI.
Na análise conjunta para as castas em cada modo de proteção, os pares agregado médio-média para ambas as castas em conjunto não se ajustaram ao modelo de regressão de Iwao, tanto na vinha conduzida em MPB (r2=8,9%) como em PRODI (r2=14,9%).
Por sua vez, na análise global para as castas e modo de produção,e tal como nos casos anteriores, os pares em estudo não se ajustaram de modo significativo ao modelo de Iwao (r2=12,3%).
A distribuição espacial das ninfas de cigarrinha-verde foi aleatória em todas as situações consideradas. No entanto, os coeficientes de determinação foram distintos para ambos os modelos utilizados. O modelo de Taylor proporcionou ajustamentos significativos à regressão, que se traduziram em coeficientes de determinação oscilando entre 91,5% e 99,0%. Em contrapartida, o modelo de regressão de Iwao não proporcionou ajustamentos significativos à regressão, cujos coeficientes de determinação variaram entre 4,8% e 14,9%. Como consequência, serão neste trabalho utilizados, para a elaboração do plano de amostragem, os coeficientes a e b de Taylor obtidos para o conjunto das castas e modos de produção.
Os resultados obtidos são consistentes com os valores obtidos em diversos países. No trabalho realizado na Suiça, Ceruti et al., (1991) obtiveram valores de agregação entre 1,11 e 1,44, enquanto Maixner (2003) no trabalho realizado na Alemanha, em três vinhas da casta Riesling, verificou a existência do padrão agregado (b=1,21) para a mesma espécie. Possíveis justificações para estas diferenças podem estar relacionadas com a heterogeneidade do vigor das plantas nas próprias vinhas, com diferentes valores médios de densidade populacional e com a própria distribuição geográfica (Delrio et al., 2001).
Relação entre a percentagem de folhas ocupadas e o número médio de formas móveis de ninfas por folha.
O ajustamento do modelo utilizado, avaliado pela análise de regressão entre a proporção de folhas ocupadas por pelo menos uma forma móvel de E. vitis e o número médio de formas móveis por folha foi significativo (r2=94,6%; n=72; p<0,001)) (Figura A). Por sua vez, a relação entre a proporção de folhas ocupadas observadas no campo e as estimadas pelo modelo proporcionou o ajustamento significativo (n=72; p<0,001), explicando em 93,86% a variação do modelo (Figura 1B). Estes resultados indicam que a proporção de folhas ocupadas pelo inseto pode ser usada como indicador do número médio de cigarrinha-verde por folha.
Determinação do tamanho da amostra
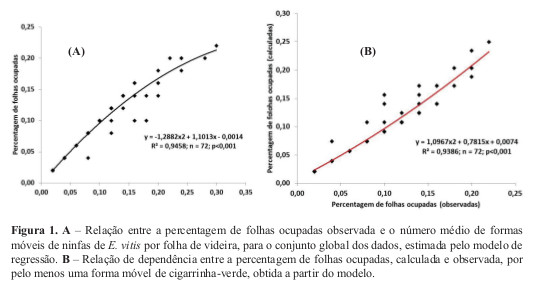
Amostragem enumerativa. A dimensão da amostra necessária para obter estimativas
populacionais de ninfas de cigarrinha-verde aumenta com o aumento do grau de precisão requerido e com a redução da densidade populacional do inseto (Figura 2). No entanto, verificou-se também que à medida que a densidade populacional da praga aumentou, o tamanho da amostra tendeu a estabilizar.
Amostragem binomial ou presença/ausência. Os resultados obtidos neste estudo revelaram que o modelo obtido proporcionou uma boa descrição da relação entre a densidade média de cigarrinha-verde por folha observada e a calculada. Por sua vez, os coeficientes de determinação elevados, obtidos a partir da relação entre a percentagem de folhas ocupadas observadas e o número médio de formas móveis de ninfas por folha (94,58%) e da correlação entre a percentagem de folhas ocupadas observada e prevista pelo modelo (93,86%) possibilitam o recurso à amostragem binomial, na qual se estima a densidade populacional de cigarrinha-verde através da simples observação da sua presença/ausência.
Estes resultados indicam que a proporção de folhas ocupadas pelo inseto pode ser usada como indicador do número médio de cigarrinha-verde por folha.
A avaliação da estimativa de risco para a cigarrinha-verde proposta para Portugal (Félix e Cavaco, 2009) baseia-se na observação de 100 folhas de videira e o nível económico de ataque é de 50 a 100 ninfas por folha, o que corresponde a 0,5% a 1,0% de folhas ocupadas. Considerando que o erro admissível para efeitos de tomada da decisão é de 20% (Soutwood e Henderson, 2003) e de acordo com os resultados obtidos, o número de folhas a observar para detectar as referidas densidades populacionais é de 57 folhas para o primeiro caso e de 30 folhas no segundo.
CONCLUSÕES
O modelo da Lei da Potência de Taylor mostrou-se mais adequado para descrever o padrão espacial de cigarrinha-verde, em comparação com o modelo de Iwao.
A cigarrinha verde apresentou o padrão espacial aleatório, em todas as situações analisadas, não se verificando diferenças estatisticamente significativas entre castas e modos de produção.
O tamanho da amostra requerido para obter estimativas populacionais de ninfas de cigarrinha-verde aumenta com o aumento do grau de precisão adotado e com a redução da densidade populacional de cigarrinha-verde.
É possível simplificar o plano de amostragem enumerativo, recorrendo apenas à observação da percentagem de folhas ocupadas pelo inseto, reduzindo desta forma o esforço amostral, sem perda de fiabilidade, constituindo importante economia de tempo na realização desta tarefa.
REFERÊNCIAS BIBLIOGRÁFICAS
Delrio, G., Lentini, A. e Serra, G. (2001) – Spatial distribution and sampling of Jacobiasca lybica on grapevine. IOBC/wprs Bulletin 24, 7: 211-219. [ Links ]
Félix, P.A. e Cavaco, M. (2009) – Manual de Proteção Fitossanitária para Proteção Integrada e Agricultura Biológica da Vinha . MADRP/DGADR, 90 p. [ Links ]
Freitas J. e Amaro, P. (2001) – Explosion de cicadelle verte dans la region du Douro au Portugal en Juillet/Août 1998. IOBC/wrps Bulletin , 24,7: 217-220. [ Links ]
Iwao, S. (1968) – A new regression method for analyzing the aggregation pattern of animal populations. Res. Popul. Ecol. ,X: 1-20. [ Links ]
Lloyd, M. (1967) – Mean crowding. The Journal of Animal Ecology, 36, 1: 1-30. [ Links ]
Maixner, M. (2003) – A sequential sampling procedure for Empoasca vitis Goethe (Homoptera: Auchenorrhyncha). IOBC/wprs Bulletin, 26, 8: 209-215. [ Links ]
Quartau, J. A. e Simões, P. (1995) – Aplicação de métodos numéricos na separação das espécies das cigarrinhas verdes da vinha no Alentejo (Homoptera: Cicadelidae). In: Actas do 3º Simpósio de Viticultura do Alentejo, 1:145-156. [ Links ]
Southwood, T.R.E. e Henderson, P.A. (2000) – Ecological Methods. 3rd Edition. Oxford, Ed. Blackwell Science, 575 p. [ Links ]
Taylor, L.R. (1961) – Aggregation, variance and the mean. Nature, 189: 732-735. [ Links ]
Villagrán, M.; Martín, P.; Soria, F.J. e Ocete, M.E. (1999) – Population dinamics of Asterolecanium ilicicola (Targioni, 1892) (Homoptera: Asterolecaniidae) in the Huelva and the development of a sampling method, Zool. Baetica, 10: 49-61. [ Links ]
Wilson, L.T. e Room, P.M. (1983) – Clumping patterns of fruit and arthropods in cotton, with implications for binomial sampling. Environ. Entomol., 21, 1: 50-54 [ Links ]

