










Serviços Personalizados
Journal

Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
 Similares em
SciELO
Similares em
SciELO Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias vol.35 no.2 Lisboa jul. 2012
Parasitismo oófago de Traça-do-Tomateiro por Trichograma Evanescens (Hymenoptera: Trichogrammatidae)
Parasitism of south American Tomato Moth eggs by Trichograma Evanescens (Hymenoptera: Trichogrammatidae)
Rosângela Payer[1], António Mexia[1], Dirceu Pratissoli[2] e Elisabete Figueiredo[1]
[1] CEER, Instituto Superior de Agronomia, Universidade Técnica de Lisboa, Tapada da Ajuda, 1349-017 Lisboa. E-mail: gpayer@gmail.com, amexia@isa.utl.pt, elisalacerda@isa.utl.pt.
[2] Dep. Produção Vegetal, Centro de Ciências Agrárias, Univ. Federal do Espírito Santo, Alto Universitário s/ nº, Alegre, ES, 29500-000, Brasil. E-mail: pratissoli@cca.ufes.br
RESUMO
Com o objectivo de utilizar espécies indígenas em protecção biológica contra traça-do-tomateiro, avaliou-se a adequação desta espécie como hospedeiro para Trichogramma evanescens. Após se verificar que este parasitóide oófago aceita como hospedeiro os ovos desta praga e consegue completar o seu desenvolvimento, avaliaram-se: a taxa de parasitismo; a proporção de ovos em que o parasitóide atingiu o estado adulto; e a taxa de viabilidade de desenvolvimento, a três temperaturas (22,0ºC, 24,0ºC e 25,5ºC). A taxa de parasitismo e a proporção de ovos com orifício de emergência foram calculadas como número de ovos parasitados e número de ovos com orifício de emergência, respectivamente, sobre número de ovos viáveis. A viabilidade de desenvolvimento do parasitóide até ao estado adulto foi calculada como número de ovos com emergência sobre número de ovos parasitados. Verificou-se parasitismo de ovos de traça-do-tomateiro em todas as temperaturas ensaiadas. A taxa de parasitismo foi mais elevada a 22,0ºC. A taxa de ovos com orifício de emergência e a viabilidade do desenvolvimento foram significativamente mais elevadas a 22,0ºC e a 24,0ºC, respectivamente.
Palavras-chave: Parasitismo oófago,protecção biológica, Tuta absoluta.
ABSTRACT
Tuta absoluta eggs were evaluated as host for Trichogramma evanescens with the aim of using indigenous species for biological control of the South American tomato moth. After being proved that T. evanescens could parasitize and achieve the complete development inside this host, parasitism rate, proportion of eggs within the parasitoid that reach the adult stage and viability of development rate, at three different temperatures (22.0ºC, 24.0ºC and 25.5ºC) were evaluated. Parasitism rate and the proportion of eggs with emerging hole were calculated as number of parasitized eggs and number of eggs with emerging holes, respectively, by the number of viable eggs. The viability of parasitoid development until adult stage was calculated as number of eggs with emerging hole/number of parasitized eggs. T. evanescens parasitized T. absoluta eggs at all the studied temperatures. Parasitism rate and proportion of eggs with emerging holes were higher at 22.0ºC; viability of development till adult was higher at 24.0ºC.
Keywords: Biological control, egg parasitism, Tuta absoluta.
INTRODUÇÃO
A traça-do-tomateiro, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae), é uma praga que causa importantes prejuízos em solanáceas em vários países da América Latina (Torres et al., 2001; EPPO, 2005; Lietti et al., 2005). De origem Neotropical, foi detectada na Europa, pela primeira vez, em Castellón, Valência, em 2006 (Urbaneja et al., 2007) e tem vindo a dispersar-se por toda a bacia do Mediterrâneo, Europa e Médio Oriente (Desneux et al., 2010, 2011). Em Portugal, a traça-do-tomateiro foi assinalada no Montijo, em Abril de 2009 (Matos, 2011), no Algarve, em estufas na campina de Faro, em Maio de 2009 e, em Julho de 2009, na região Oeste, em estufas na zona da Silveira e A-dos-Cunhados (Anónimo, 2009; Serra et al., 2009; Figueiredo et al., 2010). No final do Verão de 2009 foi detectada em armadilhas iscadas com feromona sexual, na Lezíria do Ribatejo em tomate de indústria e, no Outono, verificou-se a presença em estufas do Entre-Douro e Minho (Figueiredo, dados não public.).
As larvas da traça-do-tomateiro minam as folhas e perfuram caules, gomo apical e frutos verdes e maduros (Lietti et al., 2005; Monserrat, 2009). O tomate é o principal hospedeiro de T. absoluta, mas também ataca outras solanáceas, nomeadamente beringela, tabaco e batata, bem como espécies de solanáceas adventícias como Solanum nigrum L., Solanum elaeagnifolium Cav., Solanum valdiviense Dunal, Datura stramonium L.,Nicotiana glaucaGraham e Datura ferox L.(EPPO, 2005). A espécie Chenopodium murale L. também pode servir de alimento para as larvas (Payer e Figueiredo, dados não public.). Em Itália, foi recentemente identificado o feijão-verde, Phaseolus vulgaris L. (Fabaceae) como hospedeiro (EPPO, 2009).
Tuta absoluta apresenta 9 a 12 gerações/ano na Bacia do Mediterrâneo (EPPO, 2005; Monserrat, 2009).
Os prejuízos causados por esta praga têm sido importantes, quer em tomate protegido quer em tomate de indústria, dado o seu potencial biótico, o ciclo de vida curto, a sobreposição de gerações e não apresentar diapausa (EPPO, 2005), podendo perder-se 80-100% da produção na ausência de medidas de protecção (Desneux et al., 2010). Para limitar as populações de T. absoluta é necessária uma estratégia de integração de vários meios de protecção, culturais, biológicos, biotécnicos e químicos (Monserrat, 2009). Aliás, a utilização repetida das substâncias activas homologadas, além dos problemas toxicológicos e ecotoxicológicos decorrentes, e da incompatibilidade da protecção química com as cada vez maiores exigências do mercado relativamente à ausência de resíduos de pesticidas, têm provocado desenvolvimento de resistências da traça-do-tomateiro a várias substâncias activas (e.g. Pratissoli e Parra, 2000; Lietti et al., 2005; Silva et al., 2011).
Considerando estes aspectos, é importante estudar os agentes de protecção biológica que poderão ser usados na Bacia Mediterrânica. Os inimigos indígenas que possam adaptar-se a esta nova presa/hospedeiro devem ser privilegiados (Ehler, 2000; van Lenteren et al., 2006). Parasitóides oófagos do género Trichogramma (Hymenoptera: Trichogrammatidae) foram já experimentados, com sucesso, na protecção contra traça-do-tomateiro, em laboratório e no campo, no Brasil e, em laboratório, em Espanha, respectivamente, T. pretiosum Riley (e.g. Miranda et al., 1998; Pratissoli e Parra, 2001; Medeiros et al., 2009) e T. achaeae Nagaraja e Nagarkatti (Cabello et al., 2009 e 2010). O facto destes parasitóides oófagos poderem ser facilmente produzidos em hospedeiros alternativos e a preços compatíveis com largadas inundativas, mesmo em culturas de ar livre, como tomate de indústria, torna-os bastante interessantes como agentes de protecção biológica. Trichogramma evanescens Westwood é uma espécie que existe espontaneamente no Ribatejo e que limita naturalmente noctuídeos em searas de tomate de indústria (Silva et al., 1999; Gonçalves et al., 2006a e b), sendo o parasitóide oófago mais frequente no complexo de parasitóides da lagarta do tomate, Helicoverpa armigera (Hbn.) (Lepidoptera: Noctuidae), outra praga-chave de tomate de indústria, nestas searas (Gonçalves et al., 2006b), além de ser usado como agente de protecção biológica em tratamentos biológicos contra várias espécies de lepidópteros (Hassan, 1993; Schöller et al., 1996; Smith, 1996).
Neste trabalho, verificou-se a adequação de T. absoluta como hospedeiro de T. evanescens e avaliaram-se as taxas de parasitismo e de viabilidade do desenvolvimento até adulto, com vista a futura utilização deste parasitóide oófago indígena em tratamento biológico contra a traça-do-tomateiro.
MATERIAL E MÉTODOS
A traça-do-tomateiro foi criada e mantida em gaiolas de acrílico (30x30x40 cm3) à temperatura de 25 ± 2ºC, 60 ± 10% HR e um fotoperíodo de 16 h, em plântulas de diversas variedades de tomateiro provenientes de viveiros situados na região Oeste. Para dar início à criação de T. absoluta, recolheram-se larvas, pupas e adultos nas estufas da Silveira (Torres Vedras). As larvas e pupas provenientes do campo foram condicionadas em caixas pequenas de plástico e mantidas até a emergência dos adultos, sendo, posteriormente, introduzidos nas gaiolas de acrílico. Os adultos foram alimentados com uma solução de mel diluído em água a 50%.
Trichogramma evanescens foi fornecido pela empresa Andermatt Biocontrol, sob a forma de ovos de Ephestia kuehniella Zeller parasitados. A empresa comercializa esta espécie para utilização como agente de protecção biológica em produtos armazenados.
Com o objectivo de verificar se T. evanescens é capaz de parasitar ovos de traça-do-tomateiro, procedeu-se a marcação de alguns ovos de T. absoluta com diversas idades sobre as plântulas de duas gaiolas e colocou-se o cartão contendo pupas de T. evanescens (em ovos de E. kuehniella) em cada uma das gaiolas, bem como cartões contendo solução de 50% de mel em água, para a alimentação dos adultos do parasitóide.
Para avaliação das taxas de parasitismo e de viabilidade de desenvolvimento até adulto, adaptou-se a metodologia de Pratissoli e Parra (2000). No primeiro ensaio, Ensaio I, retiraram-se os ovos de T. absoluta das folhas do tomateiro com um pincel humedecido em água. Em seguida, isolaram-se 10 ovos para um cartão que se colocou num tubo de vidro de 13 cm de comprimento por 0,9 cm de diâmetro selado com Parafilm®, onde se tinha previamente introduzido cartão contendo gotas de mel puro para a alimentação dos adultos de T. evanescens. Colocaram-se, em cada tubo, duas fêmeas com o auxílio de um pincel, tendo o cuidado de lhes causar o mínimo de danos mecânicos. Realizaram-se oito repetições. Os tubos foram colocados em câmaras climatizadas às temperaturas de 22,0 ± 1ºC, 24,0 ± 1ºC, 25,5 ± 1ºC, sendo a HR de 70 ± 10 % e o fotoperíodo de 14h. Realizaram-se observações diárias para retirar ovos em que se estivessem a desenvolver larvas e após 10 dias contabilizou-se o número de ovos de T. absoluta parasitados bem como o número de ovos inviáveis em cada um dos tubos. No Ensaio II, procedeu-se de igual forma, à excepção de ter sido colocada apenas uma fêmea por tubo, para evitar eventual superparasitismo e os tricogramas foram atraídos para o cartão com uma gota de mel e, de seguida, inseridos nos tubos. Neste ensaio foram efectuadas 10 repetições.
Para determinar o tempo de desenvolvimento de T. evanescens em T. absoluta, procedeu-se como no Ensaio II. Foram usados 30 tubos. Estes foram colocados em câmara climatizada regulada a 24,0ºC, durante 6 h. Passado este período, retiraram-se as fêmeas e sujeitou-se cada conjunto de 10 tubos a diferentes temperaturas: 22,0 ± 1ºC, 24,0 ± 1ºC, 25,5 ± 1ºC. A humidade relativa (HR) foi de 70 ± 10 % em qualquer das câmaras e o fotoperíodo de 14 h. Realizaram-se observações diárias, sempre no mesmo horário, cerca das 11h, para verificar a emergência dos adultos.
A percentagem de parasitismo e a percentagem de ovos com orifício de emergência foram calculadas como número de ovos parasitados e número de ovos com orifício de emergência, respectivamente, sobre o número de ovos viáveis. A viabilidade do desenvolvimento até adulto foi calculada através do número de ovos com emergência sobre o número de ovos parasitados.
A comparação das taxas de parasitismo obtidas sob as temperaturas referidas foi efectuada através de análise de variância não paramétrica utilizando o teste de Kruskal-Wallis, dado que a normalidade dos dados foi rejeitada mesmo após transformação angular (arcsen Öx) das variáveis. Adoptou-se 0,05 como nível de significância. Recorreu-se ao programa SPSS (Statistical Package for the Social Sciences) Versão 17.0.
RESULTADOS E DISCUSSÃO
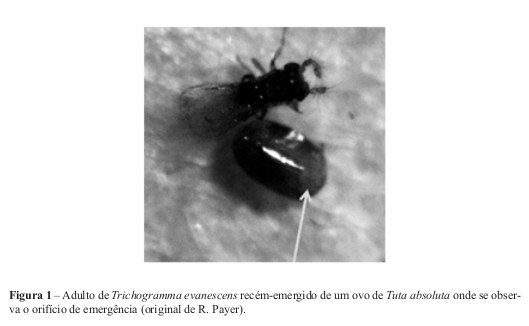
No ensaio preliminar, largando T. evanescens na gaiola, foi possível verificar que este parasitóidenão só parasitou ovos de T. absoluta, mas também conseguiu completar o seu ciclo de desenvolvimento (Figura 1). No entanto, os resultados revelaram baixo parasitismo (com o máximo de 34%). Possíveis causas deste baixo parasitismo são: as diferentes idades dos ovos marcados, sendo alguns provavelmente demasiado velhos para serem parasitados; e/ou baixa qualidade do parasitóide. De facto, verificou-se que a quantidade de ovos de Ephestia kuehniella parasitados no cartão fornecido pelo produtor de tricogramas, usado no ensaio, não era muito superior à de ovos não parasitados. A taxa de parasitismo no cartão com pupas de E. kuehniella foi de 51%, sendo bastante baixa em relação ao que é expectável (C. Gonçalves, com. pes.). Gonçalves et al. (2005) verificaram taxas mais elevadas de T. evanescens sobre E. kuehniella. Tratando-se de um produto procedente de uma biofábrica, este valor indica a baixa performance do parasitóide ou reduzido número de fêmeas usadas no parasitismo. É necessário frisar que estes insectos são sensíveis a variações bruscas de temperatura e que esta influencia aspectos biológicos inerentes ao normal desenvolvimento do parasitóide (Pratissoli e Parra, 2000; Torres, 2005). Na altura da aquisição destes insectos a temperatura média do ar sofreu uma subida brusca e parte dos adultos de T. evanescens já haviam emergido quando a encomenda foi recebida. Foi pedido um novo cartão de T. evanescens para repetir este ensaio mas, desta vez, dos ovos parasitados nem chegaram a emergir adultos. Gonçalves et al. (2006b) referem taxas elevadas de parasitismo por T. evanescens, sobre outras espécies de lepidópteros; as baixas taxas obtidas nestes ensaios podem, também, dever-se a estar a usar-se uma raça diferente. Tabone et al. (in press) referem maior diferença na eficácia de parasitismo entre raças de Trichogramma spp. do que entre espécies.
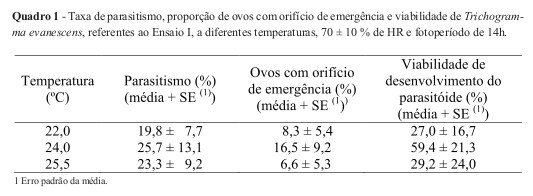
No Ensaio I, verificou-se que a percentagem de parasitismo, a proporção de ovos com orifício de emergência e a de viabilidade do parasitóide foram mais elevadas a 24,0ºC, do que a 22,0ºC e 25,5ºC, embora com valores bastante baixos (Quadro 1). É de notar que a variância associada a cada um destes valores foi elevada. Contudo, o facto de, no Ensaio I, se terem colocado duas fêmeas em cada tubo, pode ter conduzido a superparasitismo e, consequentemente, dada a dimensão reduzida do ovo da traça-do-tomateiro, a baixa percentagem de ovos em que houve desenvolvimento até ao estado adulto, expressa em baixa percentagem de ovos com orifício de emergência. Por outro lado, no Ensaio I, observou-se baixo número de ovos viáveis em alguns tubos. Este facto pode ter sido causado por estes ovos terrem sofrido danos mecânicos pela manipulação, especialmente pela dificuldade em retirá-los de folhas que, na variedade de tomateiro utilizada neste Ensaio, apresentavam demasiado indumento.
No Ensaio II, obtiveram-se maiores taxas de parasitismo do que no Ensaio I, sobretudo a 22,0ºC (Quadro 2), mas os valores foram ainda relativamente baixos para as restantes temperaturas. Esta diferença entre os dois ensaios pode dever-se ao aperfeiçoamento da metodologia laboratorial, como, por exemplo, à colocação de fêmeas nos tubos, atraindo-as para os cartões contendo mel, em vez de as seleccionar com um pincel. No Ensaio II, verificou-se que a taxa de parasitismo e a proporção de ovos com orifício de emergência foram mais elevadas à temperatura de 22,0ºC (Quadro 2). No entanto, a 24,0ºC, a percentagem de viabilidade foi mais elevada e próxima de 100%.

Na análise conjunta dos dados dos Ensaios I e II (Quadro 3), a taxa de parasitismo por T. evanescens decresceu com a temperatura, de 50,8 % a 22,0ºC, para 28,1% a 25,5ºC. Entre os ovos parasitados, verifica-se que o número de ovos que deram origem a adultos foi maior a 24,0ºC. Este menor parasitismo, a temperaturas mais elevadas, pode derivar de má adaptação desta estirpe a estas temperaturas dado que foi desenvolvida para utilização para cereais armazenados.

A análise de variância não revelou diferenças significativas nas variáveis estudadas em função da temperatura, excepto no Ensaio II. Neste último caso, em que a metodologia estava mais afinada, verificou-se a existência de diferenças estatisticamente significativas para a proporção de ovos com orifício de emergência (c2=6,324; g.l.=2; p=0,042) e viabilidade de desenvolvimento do parasitóide (c2=6,376; g.l.=2; p=0,041) mas, apenas, tendência no caso da taxa de parasitismo (c2=4,719; g.l.=2; p=0,094).
As primeiras emergências ocorreram 10, 11 e 12 dias após o parasitismo, respectivamente, para as temperaturas de 25,5ºC, 24,0ºC e 22,0ºC.
CONCLUSÕES
Trichogramma evanescens parasita e consegue completar o ciclo de desenvolvimento em ovos de Tuta absoluta. As taxas de parasitismo observadas não foram muito elevadas, sobretudo para temperaturas acima de 22,0ºC. Contudo, a estirpe usada nos ensaios, comercializada para protecção biológica em produtos armazenados, pode não estar adaptada a temperaturas mais elevadas, como as que ocorrem nas estufas ou nas searas de tomate em Portugal. Este estudo deverá ser repetido com estirpes recolhidas nas searas de tomate do Ribatejo, onde esta espécie é frequente.
AGRADECIMENTOS
Os autores agradecem ao Sr. José Firmino, agricultor da Silveira (Torres Vedras) por ter disponibilizado as suas estufas para recolha de T. absoluta para criação em laboratório. Este trabalho foi financiado pelo Financiamento Plurianual, referente a 2010, da FCT ao CEER (Centro de Engenharia dos Biossistemas).
REFERÊNCIAS BIBLIOGRÁFICAS
Anónimo (2009) - Traça-do-tomateiro - Tuta absoluta. Lourinhã, Associação Interprofissional de Horticultura do Oeste (AIHO), 2 p. [ Links ]
Cabello, T.; Gallego, J.R.; Vila, E.; Soler, A.; del Pino, M.; Carnero, A.; Hernández-Suárez, E. e Polaszek, A. (2009) - Biological control of the South American Tomato Pinworm, Tuta absoluta (Lep.: Gelechiidae), with releases of Trichogramma achaeae (Hym.: Trichogrammatidae) on tomato greenhouse of Spain. IOBCwprs Bulletin, 49: 225-230. [ Links ]
Cabello, T.; Gallego, J.R.; Fernández, F.J.; Vila, E.; Soler, A. e Parra, A. (2010) - Aplicación de parasitoides de huevos en el control de Tuta absoluta en España. Phytoma-España, 217: 53-59. [ Links ]
Desneux, N.; Wajnberg, E.; Wyckhuys, K.A.G.; Burgio, G.; Arpaia, S.; Narváez-Vasquez, C.A.; González-Cabrera, J.; Ruescas, D.C.; Tabone, E.; Frandon, J.; Pizzol, J.; Poncet, C.; Cabello, T. e Urbaneja, A. (2010) - Biological invasion of European tomato crops by Tuta absoluta: ecology, geographic expansion and prospects for biological control. Journal of Pest Science, 83:197–215. [ Links ]
Desneux, N.; Luna, M.G.; Guillemaud, T. e Urbaneja, A. (2011) - The invasive South American tomato pinworm, Tuta absoluta, continues to spread in Afro-Eurasia and beyond: the new threat to tomato world production. Journal of Pest Science, 84: 403-408. [ Links ]
Ehler, L.E. (2000) – Critical issues related to nontarget effects in classical biological control of insects. In: Follett, P.A. e Duan, J.J.(Eds.) - Nontarget effects of biological control. Norwell, Massachusetts, EUA, Kluwer Academic Publishers, p. 3-13. [ Links ]
EPPO (European and Mediterranean Plant Protection Organization) (2005) - Tuta absoluta - Data sheets on quarantine pests. Bulletin EPPO, 35: 434-435. [ Links ]
EPPO (European and Mediterranean Plant Protection Organization) (2009) - Tuta absoluta found on Phaseolus vulgaris in Sicilia (IT). EPPO Reporting Service, nº 8, 2009/154. (Acesso em 25 de Agosto de 2010). Disponível em http://archives.eppo.org/EPPOReporting/2009/Rse-0908.pdf . [ Links ]
Figueiredo, E.; Rodrigues, S.; Payer, R. e Mexia, A. (2010) - Situación actual de Tuta absoluta en Portugal. Phytoma España, 217: 118-120. [ Links ]
Gonçalves, C.I.; Amaro, F.; Figueiredo, E.; Godinho, M.C. e Mexia, A. (2005) - Productivity and quality aspects concerning the laboratory rearing of Trichogramma spp. (Hym.. Trichogrammatidae) and its factitious host, Ephestia kuehniella Zeller (Lep.: Pyralidae). Boletín de Sanidad Vegetal Plagas, 31: 19-23. [ Links ]
Gonçalves, C.; Duarte, S.; Figueiredo, E. e Amaro, F. (2006a) - Luta biológica: para quando o tratamento biológico? In: Amaro, F. e Mexia, A. (eds.) (2006) - Protecção integrada em tomate de indústria. Oeiras, EAN/INIAP: 61-68. [ Links ]
Gonçalves, C.I.; Huigens, M.E.; Verbdarschoot, P.G.H.; Duarte, S.; Mexia, A. e Tavares J. (2006b) - Natural occurence of Wolbachia-infected and uninfected Trichogramma species in tomato fields in Portugal. Biological Control, 37: 375-381. [ Links ]
Hassan, S. A. (1993) - The mass rearing and utilization of Trichogramma to control lepidopterous pests: achievements and outlook. Pest Management Science, 37, 4: 387–391. [ Links ]
Lietti, M.M.; Botto, E. e Alzogaray, R.A. (2005) - Insecticide resistance in Argentine populations of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Neotropical Entomology, 34, 1: 113-119. [ Links ]
Matos, T.M.F.L.G. (2011) - Evolução das populações de Tuta absoluta (Meyrick) e meios de protecção em ambiente empresarial vocacionado para exportação. Dissertação de mestrado. Lisboa, Instituto Superior de Agronomia /Universidade Técnica de Lisboa, 72 p. [ Links ]
Medeiros, M.A.; Villas-Bôas, G.L.; Vilela, N.J. e Carrijo, A.O. (2009) - Estudo preliminar do controle biológico da traça-do-tomateiro com o parasitóide Trichogramma pretiosum em ambientes protegidos. Horticultura Brasileira, 27: 80-85. [ Links ]
Miranda, M.M.; Picanço, M.; Zanuncio, J.C. e Guedes, R.C. (1998) - Ecological life table of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Biocontrol Science and Technology, 8: 597-606. [ Links ]
Monserrat, A.D. (Ed.) (2009) - La polilla del tomate "Tuta absoluta" en la región de Murcia: Bases para su control. Murcia, Consejería de Agricultura y Agua, Comunidad Autónoma de la Región de Murcia, 112 p. (Série Técnica y de Estudios nº 34). [ Links ]
Pratissoli, D. e Parra, J.R.P. (2000) - Desenvolvimento e exigências térmicas de Trichogramma pretiosum Riley, criados em duas traças-do-tomateiro. Pesquisa Agropecuária Brasileira, 35, 7: 1281-1288. [ Links ]
Pratissoli, D. e Parra, J.R.P. (2001) - Seleção de linhagens de Trichogramma pretiosum Riley (Hymenoptera: Trichogrammatidae) para o controle de traças Tuta absoluta (Meyrick) e Phthorimaea operculella (Zeller) (Lepidoptera: Gelechiidae). Neotropical Entomology, 30, 2: 277-282. [ Links ]
Schöller, M.; Hassan, S.A. e Reichmi, C. (1996) - Efficacy assessment of Trichogramma evanescens and T. embryophagum (Hym.: Trichogrammatidae), for control of stored products moth pests in bulk wheat. Entomophaga, 41, 1: 125-132. [ Links ]
Serra, C.; Tavares, H.; Soares, C.; Fernandes, J.E.; Ramos, N. e Figueiredo, E. (2009) - Traça-do-tomateiro (Tuta absoluta). Uma nova praga em Portugal. Direcção Geral de Agricultura e Desenvolvimento Rural, Lisboa, 2 p. [ Links ]
Silva, I.M.M.S.; Honda, J.; Van Kan, F.; Hu, J.; Neto, L.; Pintureau, B. e Stouthamer, R. (1999) - Molecular differentiation of five Trichogramma species occurring in Portugal. Biological Control, 16: 177-184. [ Links ]
Silva, G.A.; Picanço, M.C.; Bacci, L.; Crespo, A.L.B.; Rosado, J.F. e Guedes, R.N.C. (2011) - Control failure likelihood and spatial dependence of insecticide resistance in the tomato pinworm, Tuta absoluta. Pest Management Science, 67: 913-920. [ Links ]
Smith, S.M.1996.Biological control with Trichogramma: advances, successes, and potential of their use. Annual Review of Entomology, 41: 375-406. [ Links ]
Tabone, E.; Bardon, C. e Desneux, N. (em public.) - Dispersal study as Trichogrammatidae selection criteria for biological control in cauliflower greenhouses. Acta Horticulturae (ISHS). [ Links ]
Torres, M.L.G. (2005) - Controle biológico de Ecdytolopha aurantiana (Lima, 1927) (Lepidoptera: Tortricidae) com Trichogramma atopovirilia Oatman & Platner, 1983. Dissertação de mestrado, Escola Superior de Agricultura Luiz de Queiroz / Universidade de S. Paulo, 101 p. [ Links ]
Torres, J.B.; Faria, C.A.; Evangelista, W.S. e Pratissoli, D. (2001) - Within plant distribution of the leaf miner Tuta absoluta (Meyrick) immatures in processing tomatoes, with notes on plant phenology. International Journal of Pest Management, 47: 173–178. [ Links ]
Urbaneja, A.; Vercher, R.; Navarro, V.; García-Marí, F. e Porcuna, J.L. (2007) - La polilla del tomate, Tuta absoluta. Phytoma- España, 194: 16–23. [ Links ]
van Lenteren, J.C.; Bale, J.; Bigler E.; Hokkanen, H.M.T. e Loomans, A.M. (2006) -Assessing risks of releasing exotic biological control agents of arthropod pests. Annual Review of Entomology, 51: 609–634. [ Links ]

