










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias vol.36 no.4 Lisboa out. 2013
ARTIGO
Gestão do azoto no solo numa rotação hortícola no modo de produção biológico
Nitrogen management in organic horticultural rotation
Rui Pinto1, Luis Miguel Brito2 e João Coutinho3
1 Escola de Ciências da Vida e do Ambiente (ECVA), Universidade de Trás-os-Montes e Alto Douro, Apartado 1013, 5001-911 Vila Real, Portugal. E-mail: ruicasaldematos@gmail.com, author for correspondence
2 Centro de Investigação de Montanha (CIMO), Escola Superior Agrária, Instituto Politécnico de Viana do Castelo (IPVC), Refóios, 4990-706 Ponte de Lima, Portugal. E-mail: miguelbrito@esa.ipvc.pt
3 Centro de Química, Escola de Ciências da Vida e do Ambiente (ECVA), Universidade de Trás-os-Montes e Alto Douro (UTAD) 5001-911 Vila Real, Portugal. E-mail: j_coutin@utad.pt
RESUMO
Realizou-se uma rotação anual com uma cultura de cobertura de ervilhaca (Vicia sativa L.) e centeio (Secale cereale L.) como adubo verde, seguida por batata (Solanum tuberosum L.) e alface (Lactuca sativa L.) com o objetivo de contribuir para melhorar a gestão do azoto (N) no solo em agricultura biológica. A mineralização de N foi determinada através de uma incubação de campo em resposta à aplicação de adubo verde, compostado (20 e 40 t ha-1) e adubo orgânico (1 e 2 t ha-1) à cultura da batata. A mineralização de N no compostado ocorreu durante todo o período de desenvolvimento das culturas, enquanto no adubo orgânico uma percentagem elevada de N mineralizou-se logo nos primeiros dias após incorporação ao solo, do que resultou uma maior disponibilidade de N e uma produção superior na cultura da batata com 2 t ha-1 de adubo em comparação com a aplicação de 20 t ha-1 de compostado e superior na alface com a aplicação do compostado em comparação com o adubo orgânico.
Palavras-chave - Adubo verde, agricultura biológica, azoto, compostado, incubação de campo
ABSTRACT
A field crop rotation with a cover crop of hairy vetch (Vicia sativa L.) and rye (Secale cereale L.) followed by potato (Solanum tuberosum L.)and lettuce (Lactuca sativa L.) was set up with the aim of improving nitrogen (N) management for organic farming. N mineralization was determined by a field incubation in response to green manure and increasing rates of compost (20 e 40 t ha-1) and organic commercial fertilizer (1 e 2 t ha-1). N mineralization in compost took place during all crop rotation, but most of the organic fertilizer was mineralized during the first days after soil application. Consequently, N availability and potato crop yield was increased with 2 t ha-1 of organic fertilizer compared to 20 t ha-1 of compost whereas lettuce yield increased with compost application in comparison to organic fertilizer.
Keywords - Organic agriculture, compost, field incubation, green manure, nitrogen
Introdução
A fertilidade do solo é fundamental para a agricultura biológica (AB) e depende das propriedades físicas, químicas e biológicas do solo de modo a fornecer os nutrientes essenciais ao crescimento das plantas em quantidades adequadas. As práticas agrícolas para gerir a fertilidade do solo em AB incluem a rotação de culturas, a utilização de culturas de cobertura para sideração e a incorporação nos solos de compostados e outros fertilizantes aceites no modo de produção biológico (MPB). A aplicação de compostados ao solo é uma prática recomendada pelas normas da AB pois recicla os resíduos agrícolas e agro-industriais e aumenta o teor de matéria orgânica (MO) do solo (Flavel e Murphy, 2006). Como muitas culturas são exigentes em azoto (N), os adubos orgânicos comerciais são usados para rapidamente disponibilizar N às culturas, sendo frequentemente constituídos por resíduos de avicultura que são muito mais ricos em N do que, por exemplo da bovinicultura. A limitação agronómica ao uso de compostados, designadamente quando mal maturados, inclui o excesso de salinidade, o elevado teor de amónia e a imobilização de N (Brito, 2001; Wong et al., 1999).
As culturas de cobertura melhoram a estrutura do solo, evitam a sua erosão, ajudam a controlar pragas e doenças e incrementam a atividade biológica do solo (Dabney et al., 2001; Sainju et al., 2006; Hossain et al., 2012). Em AB o uso de adubos verdes é também importante pela libertação de N para a cultura seguinte (Rannels e Wagger 1996). A ervilhaca (Vicia sativa L.) acumula uma quantidade elevada de N durante o período de crescimento (Seo e Lee, 2008) e usada como adubo verde pode fornecer entre 50 a 370 kg N ha-1 à cultura seguinte devido à fixação biológica de N e à sua rápida mineralização (Tonitto, David e Drinkwater, 2006). O centeio (Secale cereale L.) é eficiente na absorção de N impedindo a sua lixiviação e tem um efeito aleopático na supressão de infestantes (Weston, 1996). A mistura de centeio com ervilhaca pode diminuir as perdas por lixiviação e desnitrificação pela presença do centeio (Rosecrance et al., 2000), tendo a quantidade de biomassa sido superior na mistura centeio e ervilhaca em relação às monoculturas de centeio ou ervilhaca (Clark et al., 1997).
O N é um nutriente crucial na fertilização em AB porque a sua disponibilidade no solo depende fundamentalmente da mineralização da matéria orgânica (MO), ao contrário dos outros nutrientes que podem ser incorporados na forma de fertilizantes inorgânicos naturais. O N mineralizado varia com as condições climáticas, cultura, tipo de solo e operações culturais e depende do balanço entre a mineralização e a imobilização (Amlinger et al., 2003; Drinkwater et al., 2000; Mengel, 1996). A previsão da mineralização de N no campo é importante para sincronizar a libertação de N mineral do solo com as necessidades das culturas (Brito, 2001) e evitar perdas de N por lixiviação ou volatilização (Jarvis, 1996). As incubações no laboratório (Gil et al., 2011) permitem prever a mineralização potencial de N, mas não refletem com precisão a mineralização de N no campo devido às perturbações no solo em consequência das operações de armazenamento, mistura e crivagem, que modificam as características físicas e microbiológicas do solo, devido à ausência de absorção de N e de libertação de exsudados pelas plantas e porque são realizadas em condições de temperatura e humidade controladas (Lomander et al., 1998). A incubação do solo no campo em sacos de polietileno (Eno, 1960) e em tubos de PVC garantindo um mínimo de perturbação do solo (Hatch et al., 1990) têm sido realizadas de modo a submeter as amostras às variações de temperatura e deste modo avaliar com maior precisão a mineralização de N no campo. O valor acumulado obtido nos vários períodos de incubação permite estimar o fluxo de N para o reservatório de N mineral do solo. Os principais inconvenientes deste método são o arejamento do solo quando se recolhe a amostra e o isolamento da amostra durante o período de incubação às variações de humidade no campo (Hatch et al., 1990). No entanto, tem sido observada uma forte correlação entre o N acumulado nas plantas e o N mineralizado durante os períodos de incubação (Carsky et al., 1990; Monaco et al., 2010). Este método é simples e provoca uma perturbação mínima no solo sendo considerado apropriado para a investigação agronómica (Monaco et al., 2010).
A batata (Solanum tuberosum L.) e a alface (Lactuca sativa L.) são duas culturas de grande importância no NW de Portugal, sendo no entanto a produção biológica restrita devido à falta de informação sobre fertilização e a proteção das culturas. Por exemplo, os agricultores precisam de conhecer a origem e o estado de maturação dos compostados e o momento e as doses de aplicação dos mesmos em função da cultura e da produção esperada. Com o objetivo de investigar sobre as recomendações de fertilização para estas culturas no MPB realizou-se uma rotação de culturas que teve inicio no Outono com a sementeira de uma cultura de cobertura para sideração (centeio e ervilhaca), seguida da batata e da alface no período de Primavera-Verão, utilizando um compostado bem maturado de estrume de bovinos e um adubo orgânico comercial com elevado teor de N facilmente mineralizável. O principal objetivo deste trabalho consistiu na quantificação da mineralização de N durante estas culturas, através de uma incubação de campo, e na absorção de N pelas mesmas de modo a avaliar a disponibilidade de N mineral em comparação com as necessidades de N destas culturas para rentabilizar as produções e incrementar a fertilidade do solo no MPB de um modo sustentável. Estas culturas inserem-se numa rotação trianual com varias culturas hortícolas de ciclo curto e longo com necessidades de fertilização diferentes, que será prosseguida de modo a compreender melhor a dinâmica do azoto em horticultura biológica.
Materiais e métodos
Realizou-se uma rotação de culturas no modo de produção biológico num solo com textura franco-arenosa, formado com base em granito e rochas afins localizado no NW de Portugal (41º 12´ N, 8º 7´ W, altitude 466 m). A rotação de culturas foi iniciada com uma cultura de cobertura constituída por centeio (Secale cereale L.) consociado com ervilhaca (Vicia sativa L. cv. Nikian) durante o período de Outono-Inverno, seguida pela cultura da batata (Solanum tuberosum L. cv. Desirée) de ciclo longo e calibre 25-28 mm, de Abril a Agosto, e da cultura da alface (Lactuca sativa L. cv. Maravilla de verano) de Agosto a Outubro. Apesar de este estudo ser referente apenas a estas três culturas, as mesmas inserem-se numa rotação trianual, que permitirá continuar esta investigação ao longo de toda a rotação.
A experiência foi conduzida com um delineamento de blocos casualizados com 6 tratamentos e quatro blocos. Os tratamentos incluíram: (i) sideração da cultura de cobertura sem aplicação de outros fertilizantes (AV); (ii) e (iii) sideração da cultura de cobertura e aplicação do compostado nas doses de 20 e 40 t ha-1 respetivamente (C20 e C40); (iv) e (v) sideração e aplicação de um adubo orgânico comercial certificado para o MPB (Mix da Crimolara) nas doses de 1 t ha-1 e 2 t ha-1 respetivamente (AO1 e AO2), e (vi) um tratamento testemunha sem cultura de cobertura para sideração nem aplicação de qualquer outro fertilizante (T0).
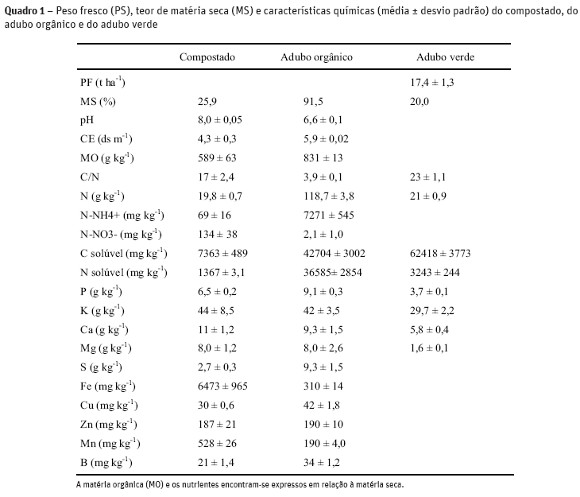
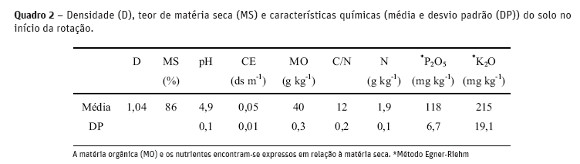
O compostado foi produzido com estrume resultante de dejetos e urinas de bovinos em cama de palha de trigo, numa pilha durante 7 meses de compostagem com revolvimento aos 2 e 4 meses. As características do compostado e do adubo orgânico comercial encontram-se no Quadro 1. O compostado estava com um elevado grau de maturação de acordo com o teor de N-NH4+ (69 mg kg-1 ms) que foi inferior ao máximo recomendado (400 mg kg-1 ms) por Zucconi e de Bertoldi (1987) e com a razão N-NH4+ /N-NO3- (0,51) inferior a 1, valor estabelecido por Larney e Hao (2007) para compostados bem maturados. Pelo contrário o adubo orgânico encontrava-se mal maturado pois continha valores do teor de N-NH4+ (7271 mg kg-1) e da razão N-NH4+ /N-NO3- (3462 mg kg-1) muito superiores aos recomendados para compostados maturados. Os valores da condutividade elétrica do compostado (4,3 ds m-1) e do adubo orgânico (5,9 ds m-1) eram superiores ao máximo de 3 ds m-1 recomendado por Soumaré et al. (2003). A relação C/N = 23 na mistura centeio e ervilhaca foi superior em relação às referidas por outros autores (Kuo e Sainju, 1998; Rosecrance et al., 2000) provavelmente devido ao baixo valor do pH do solo (Quadro 2) que não permitiu o desenvolvimento da ervilhaca (Juscafresa, 1982).
A sementeira do centeio e da ervilhaca (180 g de centeio e 90 g de ervilhaca vilosa por cada talhão de 15 m2) foi realizada manualmente após a mobilização do solo com um escarificador de 7 dentes sendo as sementes enterradas com o auxílio de um ancinho. O adubo verde foi destroçado quando a maior parte do centeio se encontrava em grão pastoso, 158 dias após a sementeira, utilizando um destroçador de martelos. Após uma secagem prévia à superfície durante 2 dias, o adubo verde foi misturado no solo à superfície, juntamente com o compostado ou o adubo orgânico nos talhões respetivos, utilizando uma fresa axial de facas. De seguida realizou-se a incorporação no solo à profundidade de 20 cm com um escarificador.
Nove dias após a incorporação do adubo verde foi semeada a batata em regos separados por 60 cm, e à distância de 25 cm na linha. As infestantes foram sachadas 2 vezes para não competirem com a batateira por nutrientes e foi realizada uma amontoa 52 dias após a sementeira. Realizaram-se dois tratamentos com hidróxido de cobre (2 kg ha-1) e um com calda bordalesa à base de sulfato de cobre (20%) previamente neutralizado com cal (20 kg ha-1) contra o míldio (Phytophthora infestans). A cultura foi regada com um sistema de mini-aspersão sempre que necessário para que a água disponível no solo não fosse limitante para o crescimento da batata.
As alfaces foram semeadas em alvéolos de polietileno de 35 ml com um substrato certificado para o MPB (Tray mix da Bas Van Buuren). Após a mobilização do solo com um escarificador, as alfaces foram transplantadas para os talhões experimentais 24 dias após a sementeira. Foram constituídos 2 camalhões de 3 linhas em cada talhão, com bordaduras de 30 cm sendo a distância de plantação 35 cm x 35 cm. As alfaces foram sachadas 2 vezes e o sistema de irrigação foi igual ao da batata. As características do solo (0-20 cm de profundidade) do campo experimental no início da rotação encontram-se no Quadro 2. O pH do solo foi corrigido com 5 t ha-1 de calcário dolomítico, e o teor de P com a aplicação de 300 kg ha-1 de fosfato de Gafsa (26,5 % P2O5). No final de cada cultura colheram-se amostras (n=6) de solo de cada talhão experimental, até 20 cm de profundidade. Colheram-se amostras (n=2) da cultura de cobertura (adubo verde) em cada talhão (repetição), lançando aleatoriamente um quadrado de arame com 0,5 m de lado no terreno, para determinação do peso fresco, do peso seco, do carbono (C) total e do teor de nutrientes (N, P, K, Ca e Mg). Nas culturas da batateira (folhas e tubérculos separadamente) e da alface colheram-se amostras de 4 plantas de cada talhão 47, 75 e 124 dias após a sementeira da batata e 23, 37 e 51 dias após a plantação da alface e realizaram-se as mesmas análises indicadas para o adubo verde. As temperaturas médias do ar e do solo foram registadas continuamente durante as culturas da batata e da alface com termístores ligados a um data logger (Delta–T Devices). Os sensores de temperatura foram colocados no solo a 10 cm de profundidade na posição horizontal e por baixo de uma placa refletora a 30 cm de altura.
Utilizaram-se as normas europeias para determinação da humidade (EN 13040:1999); pH (EN 13037:1999); condutividade elétrica ( EN 13038:1999). O N e P totais do compostado e adubo orgânico foram medidos por espectrofotometria de absorção molecular e K por fotometria de chama, depois de uma digestão com ácido sulfúrico. Os teores totais de Ca, Mg, Cu, Zn Mn, Fe e S foram determinados por espectrofotometria de absorção molecular após extração por digestão nitro-perclórica. As amostras de solo foram analisadas quanto aos teores de P e K assimiláveis pelo método de Egnér-Riehm, por extração com solução de lactato de amónio e ácido acético, sendo doseados, respetivamente, por colorimetria e fotometria de chama; o N mineral nos solos e nos compostados foi determinado com um espectrofotómetro de absorção molecular em analisador automático de fluxo segmentado após solução em KCL 1M 1:5. O C e o N solúveis dos compostados e adubo verde foram determinados em analisador elementar após extração em CaCl2 0,01M. As plantas foram secas a 65ºC até atingirem um peso constante. Nas plantas o N e o P total foram medidos por espectrofotometria de absorção molecular, o K total com um fotómetro de chama após uma digestão sulfúrica e os teores de Ca e Mg totais por espectrofotometria de absorção molecular após uma digestão nitro-perclórica.
A incubação de campo teve início após a incorporação dos fertilizantes, 9 dias antes da plantação da batata, e consistiu numa adaptação à metodologia aplicada por Hatch et al. (1990) sem etileno. Os tubos PVC utilizados tinham 15 cm de comprimento e 4 cm de diâmetro onde foram realizados 6 furos de 6 mm de diâmetro para permitir o arejamento da amostra. As amostras foram retiradas de cada talhão utilizando os tubos PVC, garantindo um mínimo de perturbação do solo. Posteriormente, as amostras foram colocadas num saco de polietileno microperfurado e enterradas verticalmente numa zona adjacente ao campo de ensaio a 20 cm de profundidade. A colheita de amostras foi realizada com um intervalo de 14 dias, exceto a primeira com um intervalo de 7 dias. Em cada período de incubação foram realizadas duas amostragens de 5 tubos cada, sendo a primeira congelada e a segunda mantida durante o período de incubação no solo. A mineralização de N do solo em cada período de incubação foi calculada diminuindo o N mineral (NH4+ + NHO3-) do solo incubado pelo N mineral da amostra congelada. O N acumulado foi estimado com base no somatório do N mineral nos vários períodos de incubação. O N mineral foi calculado em laboratório seguindo os métodos acima referidos para a determinação do N mineral do solo. Foi calculada a mineralização de N do compostado e do adubo orgânico subtraindo a mineralização de N nos tratamentos C20, C40, AO1 e AO2 pela mineralização de N no tratamento AV. A mineralização do adubo verde foi calculada subtraindo a mineralização de N no tratamento AV pela mineralização de N no tratamento T0.
A modelação dos resultados da mineralização de N do adubo verde, compostado e adubo orgânico e adubo verde foi realizada utilizando as seguintes equações de regressão não linear do modelo utilizado por Cabrera e Kessel (1988) e do modelo de Bonde e Lindberg (1988), que melhor se adaptavam aos valores observados.
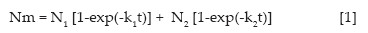
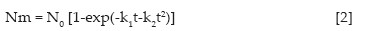
em que Nm (mg kg-1) representa a acumulação de N mineralizável no tempo t, sendo o N potencialmente mineralizável representado por N0 no modelo de um reservatório e por N1 e N2 no modelo com um reservatório de lenta e outro de rápida mineralização, e k, k1 e k2 constantes de mineralização. O tempo de imobilização foi calculado (t = -k1/k2) com base nas constantes de mineralização da equação [2] de acordo com Brito e Santos (1996).
A comparação entre os tratamentos realizou-se utilizando o procedimento para análise de variância com um fator do programa SPSS v. 17.0, e do cálculo da menor diferença significativa (P< 0,05) para comparação entre médias. Os resultados da mineralização do N orgânico foram ajustados às funções cinéticas pelo método do algoritmo de Levenberg-Marquardt, para minimizar as diferenças entre as somas dos quadrados dos valores observados e dos valores previstos da variável dependente, utilizando o mesmo programa estatístico.
Resultados e discussão
O peso fresco (PF) dos tubérculos da batateira (Figura 1) aumentou significativamente (P< 0,05) no tratamento AO2 em comparação com os restantes tratamentos aos 47 e aos 75 dias após a sementeira. No final da produção (124 dias após a sementeira) o PF dos tubérculos nos tratamentos AO1 e AO2 (35,9 e 42,2 t ha-1 respetivamente) e C40 (36,1 t ha-1) não diferiu significativamente. Contudo, no tratamento AO2 a produção foi superior (P< 0,05) aos tratamentos C20, AV e T0, sem que o mesmo fosse verdade entre estes e os tratamentos AO1 e C40. O teor de matéria seca (MS) dos tubérculos manteve-se semelhante entre os tratamentos (23-25 %).
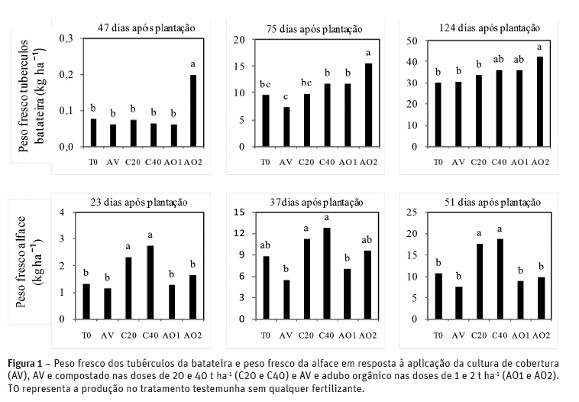
No final da produção da alface, o peso fresco (Figura 1) aumentou significativamente (P< 0,05) nos tratamentos C20 e C40 (18 e 19 t ha-1 respetivamente) em relação aos restantes tratamentos (8 a 11 t ha-1). Não se verificaram diferenças significativas no teor de MS (5-7 %) da alface entre os vários tratamentos. A máxima produção de alface registada nesta experiência com a incorporação de 40 t ha-1 de compostado foi inferior à obtida por Brito et al. (2012) com a mesma variedade e com a incorporação de 40 t ha-1 de composto (33 t ha-1), possivelmente porque esta ultima produção foi obtida com a incorporação do compostado imediatamente antes da plantação da alface.
O teor de nutrientes nos tubérculos da batateira (Quadro 3) não variou de forma significativa entre os tratamentos. No entanto os teores de N (8,1-8,9 g kg-1) e K (5,4-8,2 g kg-1) foram ligeiramente inferiores aos teores determinados por Srek et al. (2012) respetivamente 12,0 e 11,0 g kg -1 com fertilização orgânica. Os teores dos outros elementos revelaram-se semelhantes aos citados por estes autores. Verificaram-se variações ocasionais nos teores de nutrientes da alface. O teor de N foi superior nos tratamentos AV e C20 em relação ao tratamento AO1.
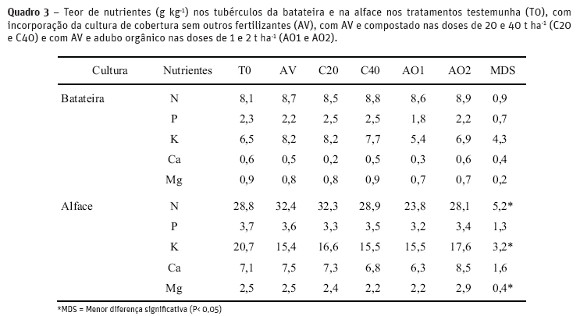
A acumulação de N na batateira (folhas e tubérculos) aumentou com a aplicação de 2 t ha-1 de adubo orgânico em comparação com a batateira não fertilizada, ou fertilizada com 20 t ha-1 de compostado (Quadro 4). A acumulação de P na batateira aumentou com a aplicação 2 t ha-1 de adubo orgânico em comparação com a batateira fertilizada só com adubo verde ou também com 1 t ha-1 de adubo orgânico. A acumulação de Ca e Mg também aumentou com a aplicação de 2 t ha-1 de adubo orgânico em relação ao tratamento testemunha, só com adubo verde ou AV e 20 t ha-1 de compostado. Não se verificaram diferenças significativas na acumulação de potássio entre os vários tratamentos (Quadro 4). O compostado aplicado à cultura da batateira nas doses de 20 e 40 t ha-1 contribuiu para o aumento (P< 0,01) na acumulação de N na cultura da alface em comparação com os restantes tratamentos. A acumulação de P, K, Ca e Mg na cultura da alface também aumentou (p <0,05) com a aplicação de 20 e 40 t ha-1 de compostado à cultura da batateira em relação aos tratamentos com AV ou AO1 (Quadro 4). A máxima acumulação de N, P, K, Ca e Mg na alface que se verificou com a incorporação de 40 t ha-1 de compostado (30,3; 3,9; 16,9; 7,5 e 2,4 kg ha-1 respetivamente) foi contudo inferior aos valores obtidos por Brito et al. (2012) respetivamente 45,7; 6,2; 56,3; 12,8; e 5,5 para a mesma variedade, em consequência da menor produção de matéria seca de alface nesta experiência.
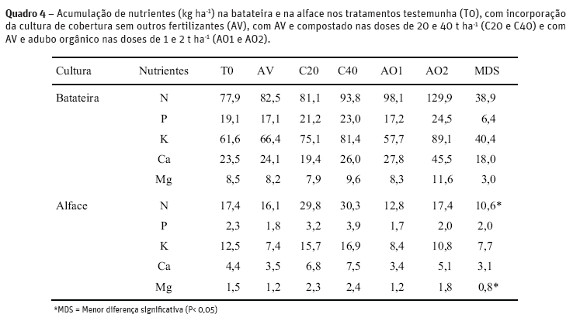
No fim da cultura da batata a mineralização de N acumulada nos tratamentos T0, AV, C20, C40, AO1 e AO2 foi respetivamente 104, 88, 152, 160, 148 e 201 kg ha-1 e no fimda rotação respetivamente 163, 137, 214, 243, 207 e 261 kg ha-1 (resultados não apresentados). A mineralização de N foi superior nos tratamentos sujeitos a fertilização com compostado e adubo orgânico, tendo sido encontrada uma correlação positiva entre a mineralização de N e a acumulação de N na batateira (R2=0,67; P< 0,05) ou a produção (R2= 0,90; P< 0,05), o que sugere que a determinação da mineralização de N na incubação de campo pode ser um bom indicador para uma previsão da recomendação de fertilização (Yan et al., 2006).
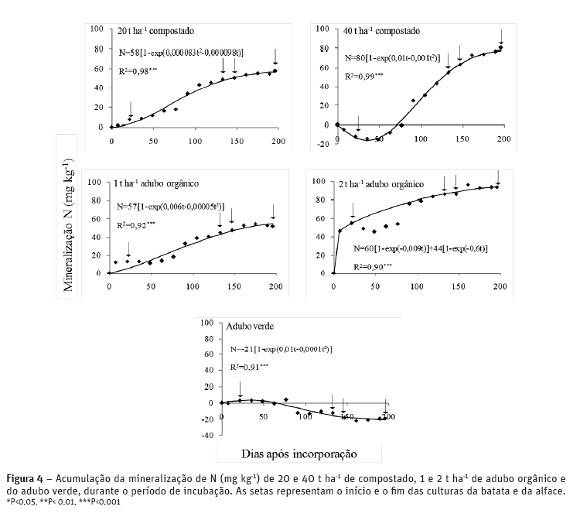
O efeito residual da fertilização à cultura da batata na produção e na acumulação de nutrientes na cultura da alface foi superior com a aplicação de compostado, porque o N mineral do adubo orgânico foi rapidamente disponibilizado durante a cultura da batata, diminuindo o efeito residual para a cultura seguinte. Na figura 2 observa-se que a mineralização de N do adubo orgânico atingiu picos elevados, 23 % (1,75 mg N kg-1 dia-1) e 49 % (6,56 mg N kg-1 dia-1) do azoto total mineralizado durante a rotação, respetivamente com 1 e 2 t ha-1 nos primeiros 7 dias de incubação quando as culturas ainda não estavam instaladas. Este facto deveu-se à baixa relação C/N (3,9) e ao teor elevado de C e N solúveis (42704 e 36585 mg kg-1 respectivamente) disponíveis neste adubo orgânico que aumentou a atividade microbiana libertando uma elevada quantidade de N num curto período de tempo. A incorporação de 40 t ha-1 de compostado resultou numa imobilização inicial de N provavelmente devida ao aumento de C biodegradável (Reddy et al., 2008), tendo a mineralização de N ocorrido durante o período de crescimento das culturas (40 dias após a plantação da batata até à colheita da alface). Probert et al. (2005) num estudo realizado com 9 estrumes de curral verificou a existência de períodos de imobilização de N superiores a 8 semanas em estrumes de curral de C/N inferiores a 20. A incorporação do adubo verde (C/N = 23) resultou numa imobilização de N média de 0,11 mg N kg-1 dia-1 durante a rotação. Normalmente é esperada uma mineralização de N nos adubos verdes com relação C/N < 20 e, eventualmente, uma imobilização de N com uma relação C/N > 20 (Rosecrance et al, 2000). Estes autores registaram uma imobilização média diária de 0,34 mg N kg-1 dia-1 no centeio (C/N = 21,4) durante 49 dias. Este facto também é devido ao teor elevado de C solúvel (62418 mg kg-1) em relação ao teor de N solúvel (3243 mg kg-1) que permitiu aos microrganismos imobilizar o N do solo para decompor o C solúvel do adubo verde.
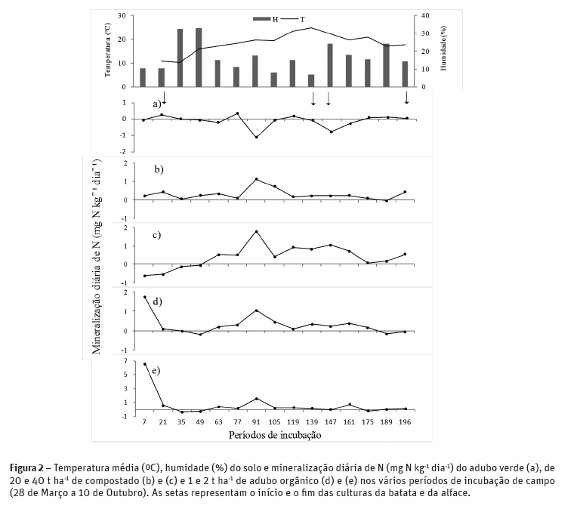
As oscilações na mineralização de N (figura 2) também dependem da interação entre a temperatura, o teor de humidade no solo e o tempo desde a incorporação dos fertilizantes orgânicos (Wang et al., 2006). Verificou-se um pico acentuado de mineralização de N em todos os fertilizantes no período de incubação entre os 77 e 91 dias (1,1; 1,8; 1,1 e 1,6 mg N kg-1 dia-1 respetivamente com 20 e 40 t ha-1 de composto e 1 e 2 t ha-1 de adubo orgânico). Este facto deveu-se em parte ao aumento de humidade do solo de 11,2 para 17,5% a uma temperatura média de 20 ºC que favoreceu as condições para a mineralização. Wang et al. (2006) referiram que com temperaturas superiores a 15ºC a mineralização de N aumenta com o teor de humidade e que o teor de mineralização de N aumentou 50 % num solo de densidade 1,15 quando o teor de humidade aumentou de 15 para 25 %. O aumento do teor de humidade de 6,9 para 24,2 % a 22 ºC entre os dias 139 e 147 após a incorporação dos fertilizantes não resultou num aumento de mineralização de N, porque possivelmente o teor de C e N orgânico potencialmente disponíveis nos vários fertilizantes era baixo.
A elevada mineralização de N no adubo orgânico verificada nos primeiros 7 dias de incubação resultou em perdas de N elevadas nos dias seguintes como se pode observar na figura 3. O teor de N mineral no solo no tratamento AO1 diminuiu de 67,3 para 18,9 mg kg-1 e no tratamento AO2 de 98,7 para 37,1 mg kg-1 dos 7 aos 35 dias após a incorporação dos fertilizantes. Estas perdas não foram devidas à absorção de N pela batateira pois no fim deste período a batateira estava ainda na fase de emergência. Possivelmente as perdas de N foram devidas à lixiviação, considerando que vários autores (Huang et al., 2011; Zhu et al., 2005) referem que a pluviosidade é responsável por perdas de N elevadas se existirem teores elevados de N-NO3- no solo. A percentagem de humidade do solo nos tubos da incubação entre os dias 21 e 49 era superior a 90 % da capacidade de campo (PF 2,0) o que indica a ocorrência de pluviosidade. No período de incubação entre os dias 7 e 21 provavelmente não houve sincronia entre a percentagem de humidade no solo dentro e fora dos tubos de incubação, sendo a percentagem de humidade no solo superior. A diminuição do teor de N mineral no solo entre os dias 77 e 105 após a incorporação dos fertilizantes (68 e 96 dias após a plantação da batata) foi em grande parte devida à absorção de N pela batateira. Gayler et al (2002) por exemplo referiram que a máxima acumulação de N nas folhas da batateira se verificou entre 40 e 90 dias após a plantação da batata. A diminuição de N mineral no solo entre os dias 147 e 196 após a incorporação dos fertilizantes corresponde ao período de absorção de N pela alface. O teor de N mineral no fim da rotação foi baixo (1,5 a 1,8 mg N kg-1) sendo praticamente idêntico ao teor de N mineral no início da rotação, o que diminui os riscos de lixiviação no período Outono-Inverno.
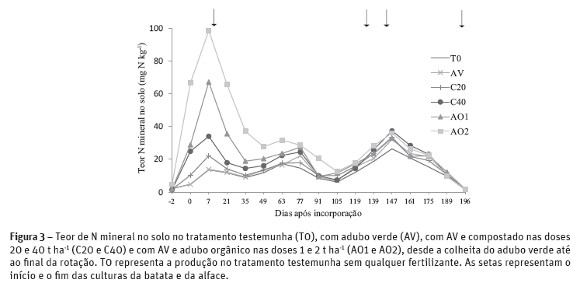
No final da rotação o N potencialmente mineralizável (N0) do compostado e do adubo orgânico foi residual porque a mineralização foi superior a 90 % desta fração de N durante a rotação (196 dias). A mineralização de N dos vários fertilizantes durante a cultura da batata (133 dias) variou entre 46 e 86 mg kg-1 MS (61 a 114 kg ha-1). Na cultura da alface a percentagem de N0 disponível foi insuficiente para obter uma produção de alface adequada a esta variedade. A maior quantidade de N mineral libertado foi 22 mg kg-1 MS (29 kg ha-1) com a incorporação de 40 t ha-1 de compostado o que explica a produção superior (P< 0,05) da alface com compostado em comparação com o adubo orgânico. Este facto deveu-se em parte ao período de imobilização de N de 72 dias com a incorporação de 40 t ha-1 de compostado que permitiu que a mineralização de N tivesse ocorrido durante o período de crescimento das culturas (figura 4).
Conclusões
A mineralização de N do compostado de estrume de bovinos ocorreu durante o período de crescimento das culturas ao contrário do adubo orgânico que libertou 23 e 49 % do N total mineralizado nos primeiros 7 dias após a incorporação ao solo para a aplicação de 1 e 2 t ha-1 respetivamente, o que implicou riscos de lixiviação de N elevados. A produção de batata com a incorporação de 40 t ha-1 de compostado e adubo verde (36 t ha-1) foi igual à produção com adubo orgânico e adubo verde. No entanto a produção de alface foi superior com a aplicação de 40 t ha-1 de compostado e adubo verde em relação aos tratamentos com adubo orgânico e adubo verde devido à maior disponibilidade de N com a aplicação de 40 t ha-1 de compostado na cultura da alface. A mineralização de N foi contudo insuficiente para obter a produção ótima desta variedade. A plantação de uma variedade de batata de ciclo médio (90 a 120 dias) em substituição da variedade de ciclo longo (120 a 150 dias) usada neste estudo poderia ser vantajosa pois, reduzindo o período de cultura da batata, a alface seria plantada mais cedo e o teor de N orgânico no solo e a temperatura média mais elevada do ar aumentariam a mineralização de N durante a cultura da alface e consequentemente a produção desta cultura. É necessário continuar este estudo com a realização de rotações de hortícolas de ciclo curto e longo de modo a otimizar a fertilização de uma rotação no MPB.
Referências bibliográficas
Amlinger, F.; Gotz, B.; Dreher, P.; Gestzi, J. e Weissteiner, C. (2003) - Nitrogen in biowaste compost: dynamics of mobilization and availability- a review. European Journal of Soil Biology, vol. 39, n. 3, p. 107-116. [ Links ]
Bonde, T.A. e Lindberg, T. (1988) - Nitrogen mineralization kinetics in soil during long-term aerobic laboratory incubations: a case study. Journal of Environmental Quality, vol.17, n.3, p. 414-417. [ Links ]
Brito, L.M.; Pinto, R.; Mourão, I. e Coutinho, J. (2012) - Organic lettuce, rye/vetch, and Swiss chard growth and nutrient uptake response to lime and horse manure compost. Organic Agriculture, vol. 2 n. 3-4, p. 163-171. [ Links ]
Brito, L.M. (2001) - Lettuce (Lactuca sativa) and cabbage (Brassica oleracea L. var. capitata L.) growth in soil mixed with municipal solid waste compost and paper mill sludge composted with bark. Acta Horticulturae (ISHS), vol.563, p. 131-137. [ Links ]
Brito, L.M. e Santos, J.Q. (1996) - Descrição quantitativa da mineralização do azoto orgânico em lixos urbanos e em lamas celulósicas. Revista de Ciências Agrárias, vol. 19, n. 1, p 35-49. [ Links ]
Cabrera, M.L. e Kessel, D.E. (1988) - Potencially mineralizable nitrogen in disturbed and undisturbed soil samples. Soil Science Society of America Journal, vol. 52, n. 4, p. 1010-1015. [ Links ]
Carsky, R.J.; Reid., W.S.; Suhet, A.R. e Lathwell, D.J. (1990) - Screening legume green manures as nitrogen sources to succeed non-legume crops. Plant and Soil, vol. 128, n. 2, p. 275-282. [ Links ]
Clark, A.J.; Decker, A.M.; Meisinger, J.J. e Mclntosh, M.S. (1997) - Kill date of vetch, rye and a vetch-rye mixture: I. Cover crop and corn nitrogen. Agronomy Journal, vol. 89, n. 3, p. 427-434. [ Links ]
Dabney, S.M.; Delgado J.A. e Reeves, D.W. (2001) - Using winter cover crops to improve soil quality and water quality. Communications in Soil and Plant Analysis, vol. 32, n. 7-8, p. 1221-1250. [ Links ]
Drinkwater, L.E.; Janke, R.R. e Rossoni-Longnecker. (2000) - Effects of tillage intensity on nitrogen dynamics and productivity in legume-based grain systems. Plant and Soil, vol. 227, n. 1-2, p. 99-113. [ Links ]
Eno, C.F. (1960) - Nitrate production in the field by incubating the soil in polyethylene bags. Soil Science Society of America Proceedings, vol. 24, p. 277-279. [ Links ]
Flavel T.C. e Murphy D.V. (2006) – Carbon and nitrogen mineralization rates after application of organic amendments to soil. Journal of Environment Quality, vol. 35, n. 1, p. 183-193. [ Links ]
Gayler, S.; Wang, E.; Priesack, E.; Schaaf, T. e Maidl, F.-X. (2002) - Modeling biomass growth, N-uptake and phenological development of potato crop. Geoderma, vol. 105, n. 3-4, p. 367-383. [ Links ]
Gil, M.V.; Carballo, M.T. e Calvo, L.F. (2011) - Modelling N mineralization from bovine manure and sewage sludge composts. Bioresource Technology, vol. 102, n. 2, p. 863-871. [ Links ]
Hatch, D.J.; Jarvis, S.C. e Philipps, L. (1990) - Field measurement of nitrogen mineralization using soil core incubation and acetylene inhibition of nitrification. Plant and Soil, vol. 124, n. 1, p. 97-107. [ Links ]
Hossain, S.; Bergkvist, G.; Berglund, K.; Martensson, A. e Persson, P. (2012) - Aphanomyces pea root rot disease and control with special reference to impact of Brassicaceae cover crops. Acta Agriculturae Scandinavica Section B-Soil and Plant Science, vol. 62, n. 6, p. 477-487. [ Links ]
Huang, M.X.; Liang, T.; Ou-Yang, Z.; Wang, L.Q.; Zhang, C.S. e Zhou, C.H. (2011) - Leaching losses of nitrate nitrogen and dissolved organic nitrogen from a yearly two crops system, wheat-maize, under monsoon situations. Nutrient Cycling in Agroecosystems, vol. 91, n. 1, p. 77-89. [ Links ]
Jarvis, S.C. (1996) - Future trends in nitrogen research. Plant and Soil, vol. 181, n. 1, p. 47-56. [ Links ]
Juscafresa, B. (1982) - Forragens. Fertilização e valor nutritivo. [S.l.], Litexa Editora, 201 p. [ Links ]
Kuo, S. e Sainju, U.M. (1998) – Nitrogen mineralization and availability of mixed leguminous and non-leguminous cover crop residues in soil. Biology and Fertility of Soils, vol. 26, n. 4, p. 346-353. [ Links ]
Larney, F.J. e Hao, X. (2007) - A review of composting as a management alternative for beef cattle feedlot manure in southern Alberta Canada. Bioresource Technology, vol. 98, n. 17, p. 3221-3227. [ Links ]
Lomander, A.; Katterer T. e Andren, O. (1998) - Modelling the effects of temperature and moisture on CO2 evolution from top and subsoil using a multi-compartment approach. Soil Biology and Biochemistry, vol. 30, n. 14, p. 2023-2030. [ Links ]
Mengel, K. (1996) - Turnover of organic nitrogen in soils and its availability to crops. Plant and Soil, vol. 181, n. 1, p. 83-93. [ Links ]
Monaco, S.; Sacco, D.;e Borda, T. e Grignani, C. (2010) - Field measurement of net nitrogen mineralization of manured soil cropped to maize. Biology and Fertility of Soils, vol. 46, n. 2, p. 179-184. [ Links ]
Probert, M.E.; Delve, R.J.; Kimani, S.K. e Dimes, J.P. (2005) - Modelling nitrogen mineralization from manures by varying C:N ratio of sub-pools. Soil Biology and Biochemistry, vol. 37, n. 2, p. 279-287. [ Links ]
Ranells, N.N. e Wagger, M.G. (1996) - Nitrogen release from grass and legume cover crop monocultures and bicultures. Agronomy Journal, vol. 88, n. 5, p. 777-782. [ Links ]
Reddy, K.S.; Mohanty, M., Rao, D.L.N.; Singh, M.; Dalal, R.C.; Rao, A.S.; Pandey, M. e Menzies, N. (2008) - Nitrogen mineralization in a Vertisol from organic manures, green manures and crop residues in relation to their quality. Agrochimica, vol. 52, n. 6, p. 377-388. [ Links ]
Rosecrance, R.C.; McCarty, G.W.; Shelton, D.R. e Teasdale, J.R. (2000) - Denitrification and N mineralization from hairy vetch (Vicia villosa Roth) and rye (Secale cereale L.) cover crop monocultures and bicultures. Plant and Soil, vol. 227, n. 1-2, p. 283-290. [ Links ]
Sainju, U.M.; Whitehead, W.F.; Singh, B.P. e Wang, S. (2006) - Tillage cover crops and nitrogen fertilization effects on soil nitrogen and cotton and sorghum yields. European Journal of Agronomy, vol. 25, n. 4, p. 372-382. [ Links ]
Seo, J.H. e Lee, H.J. (2008) - Mineral nitrogen effects of hairy vetch (Vicia villosa Roth) on maize (Zea mays L.) by green manure amounts. Journal of Agronomy, vol. 7, n. 3, p. 272-276. [ Links ]
Soumaré, M.; Tack, F.M.G. e Verloo, M.G. (2003) - Characterisation of Malian and Belgian solid waste composts with respect to fertility and suitability for land application. Waste Management, vol. 23, n. 6, p. 517-522. [ Links ]
Srek, P.; Hejcman, M. e Kunzova, E. (2012) – Effect of long-term cattle slurry and mineral N, P and K application on concentrations of N, P, K, Ca, Mg, As, Cd, Cr, Cu, Mn, Ni, Pb and Zn in peeled potato tubers and peels. Plant Soil and Environment, vol. 58, n. 4, p. 167-173. [ Links ]
Tonitto, C.; David, M.B. e Drinkwater, L.E. (2006) - Replacing bare fallows with cover crops in fertilizer-intensive cropping systems: a meta-analysis of crop yield and N dynamics. Agriculture Ecosystems & Environment, vol. 112, n. 1, p. 58-72. [ Links ]
Wang, C.; Wan, S.; Xing, X.; Zhang, L. e Han, X. (2006) - Temperature and soil moisture interactively affected soil net N mineralization in temperate grassland in Northern China. Soil Biology and Biochemistry, vol. 38, n. 5, p. 1101-1110. [ Links ]
Weston, L.A. (1996) - Utilization of allelopathy for weed management in agroecosystems. Agronomy Journal, vol. 88, n. 6, p. 860-866. [ Links ]
Wong, J.W.C.; Ma, K.K.; Fang, K.M. e Cheung, C. (1999) - Utilization of a manure compost for organic farming in Hong Kong. Bioresource Technology, vol.67, n.1, p. 43–46. [ Links ]
Yan, D.Z.; Wang, D.J.; Sun, R.J. e Lin, J.H. (2006) - N mineralization as affected by long-term N fertilization and its relationship with crop N uptake. Pedosphere, vol. 16, n. 1, p. 125-130. [ Links ]
Zhu, A.; Zhang, J.; Zhao, B.; Cheng, Z. e Li, L. (2005) - Water balance and nitrate leaching losses under intensive crop production with Ochric Aquic Cambosols in North China Plain. Environment International, vol. 31, n. 6, p. 904-912. [ Links ]
Zucconi, F. e de Bertoldi, M. (1987) - Compost specifications for the production and characterization of composts from municipal solid waste. In: de Bertoldi, M. Ferranti, M.P., LHermite, P., Zucconi, F. (Eds.) - Compost: Production, quality and use. London, Elsevier Applied Science, p. 30-50. [ Links ]
Recebido/Received: 2013.09.27
Aceitação/Accepted: 2013.09.27

