










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias vol.38 no.1 Lisboa mar. 2015
ARTIGO
Efeito dos reguladores giberelina e citocinina na quebra de dormência de sementes de capim-andropogon
Effect of gibberellin and cytokinin regulators in overcoming dormancy of grass andropogon
Francielle M. Feitosa1; Ildeu O. A. Júnior1; Andréia Márcia S. S. David1*; Bruno Rafael A. Rodrigues1; Nayara S. Damascena1; Edcássio D. Araújo1 e Hugo Tiago R. Amaro2
1 Departamento de Ciências Agrárias, Universidade Estadual de Montes Claros – Unimontes, CEP 39440-000, Janaúba – MG, Brasil. E-mails: franciellefeitosa@hotmail.com; ildeujunior.moc@hotmail.com; rafabrunoalves@hotmail.com; nayasouza22@hotmail.com; agroeda@yahoo.com.br; andreia.david@unimontes.br, author for correspondence
2 Departamento de Fitotecnia, Universidade Federal de Viçosa – UFV, CEP 36570-900, Viçosa – MG, Brasil. E-mail: htiagoamaro@yahoo.com.br
RESUMO
Nas gramíneas forrageiras tropicais, a expressão da dormência associa-se a causas físicas e fisiológicas presentes em sementes recém-colhidas. O objetivo deste trabalho foi estudar os efeitos de reguladores vegetais do grupo das giberelinas e citocininas na quebra da dormência de sementes de capim-andropogon. O delineamento experimental utilizado foi inteiramente casualizado, em arranjo fatorial (2 x 4) + 2, constituídos pela combinação de dois reguladores vegetais (giberelina e citocinina) e quatro concentrações (75; 150; 225 e 300 mg.L-1 i.a.), mais duas testemunhas (com e sem embebição em água destilada), com quatro repetições de 50 sementes por tratamento. As sementes foram imersas nas diferentes soluções por um período de 2-h. As variáveis analisadas foram: pureza física, teor de água, massa de mil sementes, germinação, primeira contagem de germinação, emergência de plântulas e índice de velocidade de emergência. A imersão das sementes de capim-andropogon em giberelina, na concentração de 225 mg L-¹, proporciona incrementos na germinação e no vigor. A citocinina, nas concentrações estudadas afeta negativamente a qualidade fisiológica das sementes.
Palavras-chave: Andropogon gayanus, germinação, reguladores vegetais, vigor.
ABSTRACT
In the tropical forage grasses, the expression of dormancy is associated with physical and physiological causes present in freshly harvested seeds. The purpose of this work was to study the effects of vegetal growth regulators in the group of gibberellins and cytokines in overcoming dormancy of seeds of grass-andropogon. The experimental delimitation was completely randomized in factorial design (2 x 4) + 2, made by the combination of two vegetal growth regulators (gibberellins and cytokines) and four concentrations (75; 150; 225 and 300 mg L-1 a.i.), two evidences (with and without soaking in distilled water), with four replications of 50 seeds per treatment. The seeds were immersed in the solution for a period of 2-h. The variables analyzed were: physical purity, water content, weight of thousand seeds, germination, first germination, abnormal seedlings, dormant seeds, seedling emergence and index of speed emergence. The immersion of seeds of grass-andropogon provides increment in gibberellins, at a concentration of 225 mg L-1, on germination and vigor. The cytokinin, the concentrations studied negatively affects seed quality.
Keywords: Andropogon gayanus, germination, plant growth regulators, vigor.
Introdução
No Brasil, cerca de 23,2% do território corresponde ao cultivo de pastagens permanentes, sendo o cerrado o segundo maior bioma do Brasil, ocupando aproximadamente em torno de 26,8% do território nacional (Sano et al., 2009). Entretanto os solos dessa região são de baixa fertilidade e elevada acidez, desta forma entende-se a necessidade de introdução de forrageiras adaptadas a esse tipo de solos e que consigam uma alta produção de biomassa, atendendo às demandas do setor agropecuário.
Neste contexto, o capim-andropogon (Andropogon gayanus Kunth var. bisquamulatus (Hochst.) Hack cv. Baetí) se apresenta como uma gramínea forrageira perene, ereta, de grande importância para pecuária brasileira, devido à elevada produção de fitomassa, resistência a seca e a baixa fertilidade, sendo cultivado nos mais variados ecossistema (Costa et al., 2001). Contudo, a maioria das espécies forrageiras tropicais não expressa todo o desempenho genético e fisiológico devido principalmente a fatores internos, com destaque a dormência, pois embora considerada um mecanismo para a sobrevivência, esta pode dificultar a expressão do potencial produtivo e a emergência das plântulas no estabelecimento de pastagens.
A dormência do capim-andropogon é fisiológica, ocorrendo quando o embrião apresenta algum mecanismo fisiológico específico que impede a protrusão da raiz primária (Vivian et al., 2008). Portanto, torna-se importante o estudo de alternativas que contribuam na descoberta de métodos que permitam a quebra de dormência dessas sementes.
O uso de hormonas como as giberelinas (Bevilaqua et al., 1993) e as citocininas (Cunha e Casali, 1989) na fase de germinação podem melhorar o desempenho de sementes de várias espécies, principalmente em condições adversas. A giberelina faz com que a raiz primária rompa os tecidos que restringem o seu crescimento, como o endosperma, o tegumento da semente ou do fruto, enquanto que as citocininas, essenciais para complementar a ação das giberelinas, são caracterizadas pela habilidade em induzir a divisão celular e na promoção do crescimento da radícula (Crozier et al., 2001; Taiz e Zeiger, 2004). Silva et al. (2013) verificaram que a utilização de giberelina proporcionou um aumento na porcentagem de germinação de sementes de Brachiaria brizantha MG 5 e Marandu.
Segundo Castro et al. (2005), a giberelina estimula a produção que enzimas hidrolíticas, as quais quebram o amido e outras substâncias, permitindo a retomada do crescimento do eixo embrionário, quebrando os mecanismos de dormência fisiológica. Adicionalmente, Garcia e Cícero (1992) ressaltam que a dormência encontrada em gramíneas é devido à impermeabilidade ao oxigénio de estruturas como o pericarpo, o tegumento e as paredes celulares, restringindo as trocas gasosas.
A aplicação e a eficiência desses tratamentos dependem da intensidade de dormência, que é bastante variável entre espécies, procedências e anos de colheita (Albuquerque et al., 2007). Além disso, dentro de um mesmo lote, pode haver sementes dormentes e não dormentes, de modo que o método deve ser efetivo na quebra da dormência, sem prejudicar as sementes não dormentes (Eira et al., 1993).
Face às considerações, objetivou-se com este estudo avaliar os efeitos de reguladores vegetais do grupo das giberelinas e citocininas na quebra de dormência de sementes de capim-andropogon.
Materiais e Métodos
O experimento foi conduzido no Laboratório de Análise de Sementes do Departamento de Ciências Agrárias (DCA), da Universidade Estadual de Montes Claros (UNIMONTES), Janaúba - Minas Gerais - Brasil, cujas coordenadas geográficas são: 15°4750 S e 43°1831 W, e altitude de 540 m. O clima da região segundo classificação de Koppen é do tipo AW (tropical com inverno seco). As condições climáticas da região são representadas por temperaturas médias que variam de 21 a 25oC, umidade relativa diária de 60 a 70% e precipitação média anual de 900 mm.
Para a condução do experimento foram utilizadas sementes de capim-andropogon, safra 2013, provenientes de uma pastagem situada no município de Janaúba-Minas Gerais. Após a colheita e processamento, as sementes foram acondicionadas em embalagem de papel e mantidas em condições ambientais, por um período de 30 dias até o início do experimento.
O delineamento experimental utilizado foi inteiramente casualizado, em arranjo fatorial (2 x 4) + 2, constituído pela combinação de dois reguladores vegetais, giberelinas (GA3) e citocinina (6-Benzilaminopurina-BAP) e quatro concentrações (75; 150; 225 e 300 mg L-1 i.a), mais duas testemunhas (com e sem embebição em água destilada), com quatro repetições de 50 sementes por tratamento. As concentrações do presente trabalho foram escolhidas com base nos resultados obtidos com outras espécies (Vieira et al., 1998; Picolotto et al., 2007; Santoe et al., 2013; Silva et al., 2013). As sementes foram imersas nas soluções contendo os reguladores vegetais e em água destilada, por um período de 2-h.
As sementes foram submetidas aos seguintes testes: pureza física, teor de água, massa de mil sementes, primeira contagem de germinação, germinação, emergência de plântulas e índice de velocidade de emergência.
Inicialmente, foi realizada a análise de pureza, conforme indicado nas Regras para Análise de Sementes - RAS (Brasil, 2009). Numa amostra de 10 g de sementes foram separados os componentes: sementes puras, outras sementes e material inerte. Cada porção foi pesada e os valores expressos em porcentagem. As demais avaliações foram realizadas a partir da porção de sementes puras obtidas nesse teste. São consideradas puras todas as sementes e/ou unidades de dispersão pertencentes à espécie em exame, declarada pelo requerente, ou como sendo a predominante na amostra e deve incluir todas as variedades botânicas e cultivares da espécie. Em outras sementes devem ser incluídas as unidades de dispersão de qualquer outra espécie de planta que não aquela da semente pura, enquanto que material inerte deve incluir unidades de dispersão e todos os outros materiais e estruturas não definidas como semente pura ou outras sementes, segundo recomendações das RAS (Brasil, 2009).
No teste de pureza física, o lote apresentou valores médios de 76% de sementes puras, sendo esse resultado acima do padrão nacional para a produção e comercialização de sementes de capim-andropogon, que é de 40% de pureza (Brasil, 2013).
O teor de água foi determinado, pelo método da estufa, a 105 ± 3oC por 24-h, com quatro repetições de cinco gramas de sementes, conforme as RAS (Brasil, 2009), indicando que as sementes apresentaram-se com 8,1% de umidade. Esses valores são adequados para a conservação de sementes de andropogon, em conformidade com os obtidos por Novembre et al. (2006) em sementes de braquiária, espécie pertencente à mesma família do andropogon.
A massa de mil sementes (g) foi obtida pelo peso de oito repetições de 100 sementes, segundo as determinações das RAS (Brasil, 2009).
A germinação das sementes foi determinada conforme as RAS (Brasil, 2009), sendo utilizadas quatro repetições de 50 sementes por tratamento. As sementes foram semeadas sobre papel mata-borrão, umedecido com um volume de água destilada equivalente a 2,5 vezes o peso do papel, em caixas plásticas tipo gerbox mantidas em germinador sob temperatura alternada de 20 °C por 16 h no escuro e 35 °C por 8-h sob luz. As avaliações foram realizadas aos 7 e 28 dias após a semeadura, quando foram avaliadas as plântulas normais, anormais e sementes dormentes, sendo os resultados expressos em porcentagem.
Os resultados do teste de primeira contagem foram obtidos pela porcentagem de plântulas normais, determinada por ocasião da primeira contagem do teste de germinação, ou seja, no sétimo dia após a semeadura (Brasil, 2009).
O teste de emergência de plântulas foi conduzido em condições ambientais de laboratório, utilizando como substrato areia lavada e esterilizada em estufa à 200 ºC, durante duas horas. As sementes foram semeadas a 0,5 cm de profundidade, em caixas plásticas tipo gerbox, contendo areia lavada e esterilizada, umedecida a 50% da capacidade de retenção (Brasil, 2009). Foram utilizadas quatro repetições de 50 sementes e os resultados foram obtidos pelo número de plântulas normais emersas, que apresentaram parte aérea visível, até a estabilização do estande ao vigésimo oitavo dias.
O índice de velocidade de emergência foi conduzido em conjunto com o teste de emergência de plântulas, anotando-se diariamente, no mesmo horário, o número de plântulas normais emersas, que apresentaram parte aérea visível, até a estabilização do estande. Ao final do teste, com os dados diários do número de plântulas emergidas, foi calculado o índice, empregando-se a fórmula proposta por Maguire (1962).
Os dados foram submetidos à análise de variância utilizando o programa estatístico SISVAR (Ferreira, 2010). Quando significativa, a comparação dos efeitos dos diferentes reguladores vegetais foi realizada por meio do teste Tukey a 5% de significância, já os estudos dos efeitos das diferentes concentrações foi realizado por meio da análise de regressão. Os modelos de regressão utilizados foram escolhidos com base na significância dos coeficientes, coeficiente de determinação (R²) e comportamento biológico. A comparação das médias dos tratamentos em relação às testemunhas foi realizada por meio da aplicação do teste Dunnett a 5% de significância.
Resultados e Discussão
Os resultados médios obtidos na massa de mil sementes foram de 3,33 g, valores próximos aos encontrados por Italiano (2000), com a mesma espécie em estudo.
No Quadro 1 encontra-se o resumo da análise de variância dos dados referentes a ação das giberelinas e citocininas na quebra da dormência das sementes de capim-andropogon. Houve efeito significativo da fonte de variação de hormona vegetal (H) sobre as variáveis emergência de plântulas e índice de velocidade de emergência. A emergência de plântulas foi ainda influenciada significativamente pelas concentrações estudadas (C), enquanto a interação entre os fatores H x C foi significativa para as variáveis emergência de plântulas e germinação. As giberelinas e citocininas nas concentrações estudadas não influenciaram nos resultados das variáveis primeira contagem de germinação e sementes dormentes.
Comparando estatisticamente as médias dos tratamentos com as médias das testemunhas (Quadro 2) observou-se que, para a variável germinação, a adição das giberelinas na concentração de 225 mg L-1 foi superior à testemunha sem embebição em água destilada, resultando num incremento de 50%, com valores de 26% de germinação, valor este superior ao mínimo exigido para todas as classes de sementes de capim-andopogon (Brasil, 2013). Enquanto que, para a variável emergência de plântulas, as sementes quando imersas em solução de giberelina na concentração de 225 mg L-1 exibiram resultados estatisticamente diferentes das testemunhas (com e sem embebição em água destilada), atingindo uma percentagem máxima de emergência de 30%, resultando em um desempenho de 68,4 e 45% superior em relação às testemunhas com e sem embebição em água destilada, respectivamente.
Com o desdobramento da interação H x C, verifica-se que apenas na concentração de 225 mg L-1 houve diferença estatística entre as hormonas vegetais estudadas (Quadro 3). Observa-se que nesta concentração (225 mg L-1) o tratamento com giberelinas mostrou-se superior em relação à citocinina, pois promoveu incrementos na germinação das sementes de andropogon. As citocininas teriam um papel permissivo na regulação da dormência, permitindo que as sementes se tornem mais sensíveis a ação das giberelinas, não sendo constatado nas sementes de andropogon (Khan, 1971).
Na cultura da soja, Leite et al. (2003) concluíram que a aplicação conjunta de giberelina e citocinina tendeu a diminuir os efeitos da giberelina, visto que o número de plântulas emergidas aos 15 dias foi reduzido com o tratamento de sementes, resultados estes compatíveis com os apresentados neste estudo, em que o biorregulador apresenta em sua formulação ambas as hormonas.
Os resultados obtidos no presente estudo concordam com os encontrados por Braun et al. (2010) com Beta vulgaris L.; Oliveira et al. (2010) com atemoia (Annona cherimola Mill. x Annona squamosa L.); Peixoto et al. (2011) com Ricinus communis L. e Silva et al. (2013) com B. brizantha Marandu e MG 5, os quais constataram a quebra da dormência de sementes de diversas espécies vegetais quando tratadas com giberelinas. A giberelina na fase de germinação das sementes melhora o desempenho das plântulas, acelerando a velocidade de emergência e realçando o seu potencial (Lopes e Souza, 2008).
Na Figura 1 são apresentados os dados relativos à emergência de plântulas oriundas de sementes de capim-andropogon tratadas com diferentes concentrações de giberelinas e citocininas. Nota-se comportamento diferenciado entre as concentrações analisadas apenas para as giberelinas, cujos resultados de emergência de plântulas se adequaram a um modelo quadrático de regressão, com percentuais máximo e mínimo de 13,5 e 30% nas concentrações de 75 e 225 mg L-1. Dentro da faixa das concentrações testadas, a emergência de plântulas foi crescente até de 225 mg L-1, atingindo o seu máximo percentual. Entretanto, a partir dessa concentração houve redução na emergência de plântulas, encerrando com 14% na concentração de 300 mg L-1.
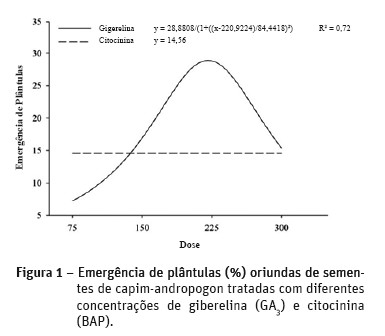
Em trabalho semelhante, Dantas et al. (2001) verificaram que sementes de Brachiaria plantaginea escarificadas e mergulhadas em solução contendo giberelinas na concentração de 0,5 mmol L-1, tiveram um aumento na emergência de plântulas em relação ao controle, e que concentrações elevadas de giberelina podem reduzir a porcentagem de plântulas emergidas. No entanto, em B. brizantha o tratamento com ácido giberélico não resultou na completa emergência de plântulas, sugerindo que outro ponto de controle da dormência fisiológica possa estar atuando juntamente à giberelina, ou que mais uma substância promotora de germinação possa estar envolvida na quebra da dormência destas sementes além da giberelina (Vieira et al.,1998).
Em relação ao efeito das citocininas, segundo Cohn e Butera (1982), esses reguladores têm se mostrado eficientes na quebra de dormência de arroz vermelho, desde que a casca tenha sido retirada. Supõe-se que a casca não atue apenas limitando a entrada, mas como um segundo fator inibidor da germinação. O modo de ação das citocininas na quebra da dormência é incerto, nesse sentido, Abeles (1986) supôs que a promoção da germinação se dava por aumento do crescimento potencial do embrião, via alongamento do hipocótilo. Segundo Vivian et al. (2008), as citocininas propiciam a superexpressão de outros compostos que atuam em mecanismos de quebra da dormência. Um dos efeitos primários das citocininas é alterar a expressão de diversos genes, incluindo aqueles que codificam a enzima redutase do nitrato e reguladores da luz
A avaliação da emergência de plântulas nos testes de vigor é importante, pois demonstram o potencial das sementes, sob condições de campo, de resistirem às condições adversas impostas pelo meio. Sementes de baixo vigor determinam redução, retardamento e desuniformidade na emergência. Nesse sentido observa-se que pelo teste de emergência de plântulas a concentração de 225 mg L-1 de giberelina promoveu um incremento na emergência de plântulas, apresentando um maior potencial fisiológico no estabelecimento e desenvolvimento do estande.
Conclusão
A imersão das sementes de capim-andropogon em giberelina, na concentração de 225 mg L-¹, proporciona incrementos na germinação e no vigor.
A citocinina, nas concentrações estudadas afeta negativamente a qualidade fisiológica das sementes.
Referências Bibliográficas
Abeles, F.B. (1986) - Role of ethylene in Lactuca sativa cv. Grand Rapids seed germination. Plant Physiology, vol. 81, n. 3, p. 780-787. [ Links ]
Aguiar, L.M.S.; Machado, B.M. e Marinho-filho, J. (2004) - A diversidade biológica do cerrado. In: Cerrado: ecologia e caracterização (L.M.S. Aguiar e A.J.A Camargo, eds.). Embrapa Cerrados, Planaltina, p.17-40. [ Links ]
Albuquerque, K.S.; Guimarães, R.M.; Almeida, Í.F. e Clemente, A.C.S. (2007) - Métodos para a quebra da dormência em sementes de sucupira-preta (Bowdichia virgilioides KUNTH.). Ciência e Agrotecnologia, vol. 31, n. 6, p. 1716-1721. [ Links ]
Bevilaqua, G.A.P.; Peske, S.T.; Santos Filho, B. G. e Baudet, L. (1993) - Desempenho de sementes de arroz irrigado tratadas com regulador de crescimento. II. Efeito na germinação. Revista Brasileira de Sementes, vol. 15, n. 1, p. 75-80. [ Links ]
Brasil. (2013) - Ministério da Agricultura, Pecuária e Abastecimento. Portaria nº 16, de 25 de janeiro de 2013, DOU de 28/01/2013, nº 19, Seção 1, pág. 2. [ Links ]
Brasil. (2009) - Ministério da Agricultura, Pecuária e Abastecimento. Regras para Análise de Sementes. Brasília: SNAD/DNDV/CLAV, 365 p. [ Links ]
Braun, H.; Lopes, J.C.; Souza, L.T.; Schmildt, E.R.; Cavatte, R.P.Q. e Cavatte, P.C. (2010) - Germinação in vitro de sementes de beterrabas tratadas com ácido giberélico em diferentes concentrações de sacarose no meio de cultura. Semina: Ciências Agrárias, vol. 31, n. 3, p. 539-546. [ Links ]
Castro, P.R.C.; Kluge, R.A. e Peres, L.E.P. (2005) - Manual de fisiologia vegetal: teoria e pratica. Piracicaba: Agronômica Ceres, 640 p. [ Links ]
Cohn, M.A., Butera, D.L. (1982) - Seed dormancy in red rice (Oryza sativa). II. Response to cytokinins. Weed Science, vol. 30, p. 200-205. [ Links ]
Costa, N.L.; Townsend, C.R.; Magalhães, J.A. e Pereira, R.G. de A. (2001) - Formação e manejo de pastagens de capim-andropogon em Rondônia. Embrapa Porto Velho. Rondônia. Recomendações Técnicas, 25. 2 pp. [ Links ]
Crozier, A.; Kamiya, K.; Bishop, G. e Yokota, T. (2001) - Biosynthesis of hormones and elicitor molecules. In: Buchanan, B.B.; Gruissem, W. e Russel, L.J. (eds). Biochemistry & molecular biology of plants. Courier Companies Inc., New York, USA, p.850-929. [ Links ]
Cunha, R. e Casali, V.W. (1989) - Efeito de substâncias reguladoras do crescimento sobre a germinação de sementes de alface. Revista Brasileira de Fisiologia Vegetal, vol.1, n.2, p.121-132. [ Links ]
Dantas, B.F.; Alves, E.; Aragão, C.A.; Tofanelli, B.D.; Corrêa, M.R.; Rodrigues, J.D.; Cavariani, C. e Nakagawa, J. (2001) - Germinação de sementes de capim-marmelada (Brachiaria plantaginea (link) hitchc.) tratadas com ácido giberélico. Revista Brasileira de Sementes, vol. 23, n. 2, p. 27-34. [ Links ]
Eira, M.T.S.; Freitas, R.W.A. e Mello, C.M.C. (1993) - Quebra da dormência de sementes de Enterolobium contortisiliquum (VELL.) Morong.-Leguminosae. Revista Brasileira de Sementes, vol. 15, n. 2, p. 177-182. [ Links ]
Eira, M.T.S. (1983) - Comparação de métodos de quebra de dormência em sementes de capim andropogon. Revista Brasileira de Sementes, vol. 5, n. 3, p. 37-50. [ Links ]
Ferreira, D. F. (2010) - Sisvar: Versão 5.3. Lavras: UFLA [ Links ] J. e Cícero, S.M. (1992) - Quebra de dormência em sementes de Brachiaria brizantha cv. Marandu. Scientia Agrícola, vol. 49, n.l, p. 9-13. [ Links ]
Italiano, E.C.C. (2000) - Determinação da época de colheita de sementes do Andropogon gayanus Kunth para a região Meio-Norte do Brasil. Pastura Tropicales, vol. 22, n. 2, p. 29-33. [ Links ]
Khan, A.A. (1971) - Cytokinins: Permissive role in seed germination. Science, vol. 171, n. 3974, p. 853-859. [ Links ]
Lopes, H.M. e Souza, C.M. (2008) - Efeitos da giberelina e da secagem no condicionamento osmótico sobre a viabilidade e o vigor de sementes de sementes de mamão (Carica papaya L.). Revista Brasileira de Sementes, vol. 30, n. 1, p. 181-189. [ Links ]
Maguire, J.D. (1962) - Speed of germination - aid in selection and evaluation for seedling emergence and vigor. Crop Science, vol. 2, p. 176-177. [ Links ]
Novembre, A.D.L.C.; Chamma, H.M.C.P. e Gomes, R.B.R. (2006) - Viabilidade das sementes de braquiária Pelo teste de tetrazólio. Revista Brasileira de Sementes, vol. 28, n.2, p. 147-151. [ Links ]
Oliveira, M.C.; Ferreira, G.; Guimarães, V.F. 3 Dias, G.B. (2010) - Germinação de sementes de atemoia (annona cherimola Mill. x a. squamosa L.) cv gefner submetidas a tratamentos com ácido giberélico (GA3) e ethephon. Revista Brasileira de Fruticultura, vol. 32, n. 2, p. 544-554. [ Links ]
Peixoto, C.P.; Sales, F.J.S.; Vieira, E.L.; Passos, A.R. e Santos, J.M.S. (2011) - Ação da giberelina em sementes pré-embebidas de mamoneira. Comunicata Scientiae, vol. 2, n. 2, p. 70-75. [ Links ]
Silva, A.B.; Landgraf, P.R.C. e Machado, G.W.O. (2013) - Germinação de sementes de braquiária sob diferentes concentrações de giberelina. Semina: Ciências Agrárias, vol. 34, n. 2, p. 657-662. [ Links ]
Taiz, L. e Zeiger, E. (2004) - Fisiologia vegetal. Art-med Editora S.A., Porto Alegre, Brasil, 719 p. [ Links ]
Vieira, H.D.; Silva, R.F. e Barros, R. S. (1998) - Efeito de substâncias reguladoras de crescimento sobre a germinação de sementes de braquiarão cv. Marandu. Revista Brasileira de Fisiologia Vegetal, vol. 10, n. 2, p. 143-148. [ Links ]
Vivian, R.; Silva, A.A.; Gimenes, Jr., M.; Fagan, E.B.; Ruiz, S.T. e Labonia, V. (2008) Dormência em sementes de plantas daninhas como mecanismo de sobrevivência –Breve revisão. Planta Daninha, vol. 26, n. 3, p. 695-706. [ Links ]
Recebido/Received. 2014.06.01
Aceite/Accepted: 2014.12.16

