










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias vol.38 no.2 Lisboa jun. 2015
ARTIGO
Aspectos biológicos de Tuta absoluta (Lepidoptera: Gelechiidae) criada em folhas de diferentes genótipos de tomate
Biological aspects of Tuta absoluta (Lepidoptera: Gelechiidae) reared on leaves of different tomato genotypes
Daline B. Bottega1, Arlindo L. Boiça Junior2, Bruno H. S. Souza1 e André L. Lourenção3
1 Departamento de Fitossanidade, Instituto Federal de Educação, Ciência e Tecnologia Goiano - Campus Iporá, 76,200-000, Iporá-GO, Brasil, Avenida Oeste s/n, saída para Piranhas. E-mail: daline.bottega@ifgoiano.edu.br, author for correspondence
2Departamento de Fitossanidade, Faculdade de Ciências Agrárias e Veterinárias, Universidade Estadual Paulista, 14884-900, Jaboticabal, Brasil. E-mails: aboicajr@fcav.unesp.br, souzabhs@gmail.com.
3 Centro de Pesquisa e Desenvolvimento de Fitossanidade, Entomologia, Instituto Agronómico de Campinas, 13020-902, Campinas-SP, Brasil. E-mail: andre@iac.sp.gov.br.
RESUMO
Avaliou-se, em condições de laboratório, a resistência de genótipos de tomateiro, Lycopersicum esculentum(Miller) (cv. ‘Santa Clara' e híbridos: Saladete Italiano Ty Tyna, Santa Cruz Débora Ty, Salada Lumi e Saladete Italiano Andrea Victory), Lycopersicum hirsutum f. glabratum (C.H. Mull.) (linhagens PI 134417 e PI 134418) e Lycopersicum pimpinellifolium (Just) (linhagens NAV 1062 e PI 126931) sobre Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). A linhagem PI 134418 foi, dentre os genótipos avaliados, a que apresentou efeito mais negativo sobre T. absoluta, em relação à testemunha (Santa Clara), alongando a fase larval, reduzindo a sobrevivência larval e a sobrevivência global. A linhagem PI 134417 também prejudicou o desenvolvimento de T. absoluta, alongando a fase larval e reduzindo a sobrevivência larval. Os demais genótipos não apresentaram efeito negativo sobre a biologia de T. absoluta. Concluiu-se que PI 126931 é suscetível, e NAV 1062, Santa Clara, Tyna, Débora, Lumi e Andrea Victory são altamente suscetíveis a T. absoluta, enquanto PI 134418 e PI 134417 destacaram-se com alta resistência e moderada resistência, respectivamente, a T. absoluta, do tipo antibiose.
Palavras-chave: resistência de plantas, traça-do-tomateiro, antibiose.
ABSTRACT
The resistance of tomato genotypes, Lycopersicon esculentum (Miller) (cv. Santa Clara, hybrids: Saladette Italian Ty Tyna, Santa Cruz Débora Ty, Salad Lumi and Saladette Italian Andrea Victory), Lycopersicon hirsutum f. glabratum (C,H, Mull.) (lines PI 134417 and PI 134418) and Lycopersicum pimpinellifolium (Just) (lines NAV 1062 and PI 126931) to Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) was evaluated under laboratory conditions. The line PI 134418 had the most negative effect on T. absoluta in comparison to the control (Santa Clara), lengthening the larval stage and reducing larval and global survival. The line PI 134417 also affected the development of T. absoluta lengthening the larval stage and reducing the larval survival. The other genotypes showed no negative effect on the biology of T. absoluta. We concluded that PI 126931 is susceptible and NAV 1062, Santa Clara, Tyna, Débora, Lumi and Andrea Victory are highly susceptible to T. absoluta, whereas PI 134418 and PI 134417 highlighted with high resistance and moderate resistance, respectively, to T. absoluta of antibiosis-type.
Keywords: host plant resistance, tomato leaf miner, antibiosis
Introdução
O tomateiro (Lycopersicum esculentum Miller) tem como centro de origem a região andina, e, no Brasil, sua introdução deve-se a imigrantes europeus no final do século XIX (Alvarenga, 2004). Esta é a hortícola mais cosmopolita e amplamente disseminada no mundo, entretanto, não há na agricultura brasileira outra cultura tão complexa, do ponto de vista agronómico e de riscos económicos tão elevados, em virtude principalmente da infestação por diversas pragas, tanto na cultura para consumo in natura, como para indústria (Filgueira, 2005; Gonçalves Neto et al., 2010).
A infestação por insetos-praga pode ocorrer durante todo o ciclo, e até mesmo em culturas protegidas e os ataques podem causar danos consideráveis (Silva e Carvalho, 2004). A praga-chave da cultura é a traça-do-tomateiro, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) (Gonçalves Neto et al., 2010), que na fase imatura ataca toda a planta em qualquer estádio de desenvolvimento, como as folhas, brotos apicais, caules, botões florais e até os frutos, formando galerias transparentes, principalmente nas gemas apicais, nas quais destroem brotações novas, além dos frutos, que são depreciados para comercialização (Silva e Carvalho, 2004).
Com o objetivo de diminuir os prejuízos ocasionados por esta praga, têm-se utilizado inseticidas de forma indiscriminada, ocasionando problemas no controle deste inseto, como a resistência de T. absoluta aos seus ingredientes ativos (Siqueira et al., 2001), além de exercerem grande impacto sobre inimigos naturais, afetando principalmente as áreas onde o controle biológico é praticado.
Diante deste fato, é de fundamental importância o desenvolvimento de variedades resistentes a T. absoluta a fim de diminuir o número de aplicações de inseticidas nessa cultura. Assim, torna-se necessário o conhecimento das possíveis fontes de resistência do tomateiro ao inseto (Suinaga et al., 2004), pois a resistência de plantas mostra-se eficiente para diminuir os riscos económicos e ambientais que prejudicam a produção e apresentam várias vantagens, como baixo custo, facilidade de utilização e compatibilidade com outras táticas de controle (Lara, 1991).
Várias fontes de resistência a diversas pragas do tomateiro têm sido identificadas, sendo que, dentre essas, a espécie Lycopersicon hirsutum f. glabratum (C. H. Mull.) tem-se destacado por apresentar o aleloquímico 2-tridecanona (2-TD), substância considerada tóxica a vários insetos (Linden, 1996). Trabalhos realizados com a traça-do-tomateiro demonstram que o 2-TD confere resistência da planta a essa praga. Giustolin e Vendramim (1994) observaram, em laboratório, alongamento das fases larval e pupal, redução da sobrevivência larval, menor peso de pupas e menor fecundidade das fêmeas na linhagem PI134417 de L. hirsutum f. glabrum. Thomazini et al. (2001) também constataram que PI134417 afeta tanto a biologia como o comportamento de oviposição de T. absoluta.
Algumas variedades de L. esculentum, como Gigante Orita e Príncipe Gigante, também apresentam menor suscetibilidade a algumas pragas, tais como Helicoverpa zea (Boddie), T. absoluta, Macrosiphum euphorbiae (Thomas), Spodoptera exigua (Hübner) e Tetranychus urticae (Koch)(Fornazier et al., 1986; Eigenbrode e Trumble 1993, 1994). Essa resistência se deve, em alguns casos, aos tricomas glandulares e/ou à lamela foliar. Além disso, características físicas (espessura da camada cuticular) e químicas dos frutos e hábito de crescimento (Leite et al., 2003) também influenciam na resistência de L. esculentum a pragas.
Assim, teve-se por objetivo, avaliar a resistência de nove genótipos de tomateiro, sendo cinco comerciais e quatro linhagens selvagens, sobre o desenvolvimento de T. absoluta, em condições de laboratório.
Material e Métodos
O estudo foi desenvolvido na Faculdade de Ciências Agrárias e Veterinárias – FCAV/UNESP, Jaboticabal, SP, no Departamento de Fitossanidade, Laboratório de Resistência de Plantas a Insetos, à temperatura de 25±1 ºC, UR de 70±10% e fotofase de 12-h, com as espécies de tomateiro L. esculentum (cv. ‘Santa Clara' (testemunha) e quatro híbridos: Saladete Italiano Ty Tyna, Santa Cruz Débora Ty, Salada Lumi e Saladete Italiano Andrea Victory), L. hirsutum f. glabratum (linhagens PI 134418 e PI 134417) e Lycopersicon pimpinellifolium (Just) (linhagens NAV 1062 e PI 126931), as quais foram cultivadas em vasos (40 x 20 cm2) mantidos em estufa. As plantas foram utilizadas com 60 dias após a emergência.
A praga foi criada em gaiolas de vidro retangular (0,4 m x 0,4 m de base e 0,5 m de altura) com uma abertura lateral, revestida com tecido voile. Folíolos de tomateiro da cultivar ‘Santa Cruz Kada Paulista' foram retirados das plantas cultivadas em estufa e suas hastes mergulhadas em potes de vidro (10 cm de altura e 4 cm de diâmetro) contendo água e fechados com algodão hidrófogo. Estes foram colocados no interior das gaiolas, onde os adultos foram libertados, obtendo-se os ovos. As hastes com ovos foram transportadas para outra gaiola de mesmas dimensões, para a eclosão das lagartas. Ao atingir a fase de pupa, estas foram deixadas no interior das gaiolas com as hastes secas, até a emergência dos adultos, que novamente foram libertados nas gaiolas de oviposição (Miranda et al., 1998). Os adultos foram alimentados com solução de mel a 10%..
O delineamento experimental utilizado foi inteiramente casualizado, composto por nove tratamentos e 20 repetições, cada uma contendo três lagartas.
Primeiramente, obtiveram-se os ovos da criação, os quais foram colocados, com o auxílio de um pincel, em placas de Petri (6 cm de diâmetro) até à eclosão das lagartas. As lagartas recém-eclodidas foram colocadas em placas de Petri (6 cm de diâmetro) contendo papel filtro levemente umedecido e folhas de cada genótipo. As folhas fornecidas às lagartas estavam completamente desenvolvidas e a cada dois ou três dias foram fornecidas folhas novas dos genótipos em estudo (Giustolin et al., 2002), retiradas da parte mediana e apical da planta (Leite et al., 1999).
As placas de Petri foram observadas diariamente, anotando-se a ocorrência de pupas que, 24h após a formação, foram pesadas e individualizadas em tubos de vidro (8,5 cm de altura x 2,5 cm de diâmetro), onde permaneceram até a emergência dos adultos, que foram sexados.
Os parâmetros avaliados foram: duração (dias) e sobrevivência (%) das fases larval e pupal, sobrevivência (%) de e lagartas de primeiro instar à emergência dos adultos (global), duração (dias) global, longevidade dos adultos sem alimento, massa total de pupas (medida com 24 horas de idade) e razão sexual.
Os resultados obtidos foram submetidos à análise de variância (ANOVA), seguindo a distribuição de F, sendo as médias comparadas pelo teste de Tukey, ao nível de 5% de probabilidade. Os dados de duração larval, pupal e global, longevidade dos adultos sem alimento, massa de pupas e razão sexual foram transformados em (x + 0,50)1/2 e os dados de sobrevivência transformados em arcsen [(x + 0,50)/100]1/2.
Realizou-se, também, a análise de agrupamento hierárquico, utilizando-se o método Single Linkage e a distância euclidiana como medida de dissimilaridade, além da análise dos componentes principais, a fim de se classificar os genótipos que apresentassem a máxima similaridade e a mínima dissimilaridade entre os grupos. Para estas duas análises foram usados os parâmetros: Sobrevivência larval (SL), pupal (SP) e global (ST), duração larval (DL), pupal (DP) e global (DG), massa de pupas (MP), longevidade de adultos(L) e razão sexual (RS), com o uso do programa Statistica versão 7.0 (Statsoft, 2004).
Resultados e Discussão
A duração da fase larval foi significantemente mais alongada nos genótipos PI 134417 (20,0 dias) e PI 134418 (20,3 dias) (L. hirsutum f. glabratum) em relação aos criados em Andrea Victory (16,3 dia) e Tyna (16,4 dia) (L. esculentum) (Quadro 1).
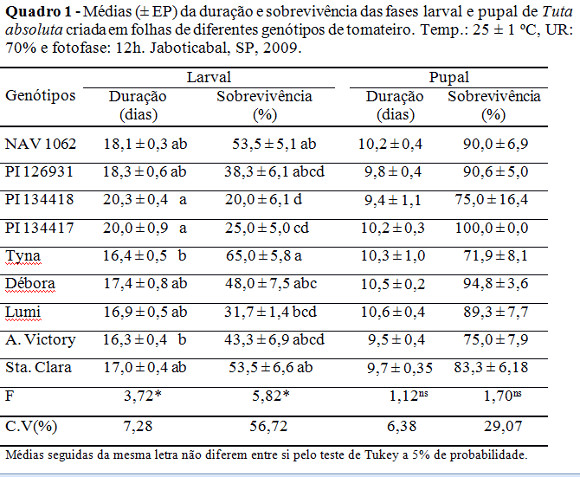
A sobrevivência da fase larval foi significativamente reduzida quando o inseto foi criado nos genótipos PI 134417 (25,0%) e PI 134418 (20,0%) em comparação àqueles criados em ‘Santa Clara' (testemunha) (53,5%), Tyna (65,0%) e NAV 1062 (53,5%)(Quadro 1). Nos outros genótipos os valores não diferiram do encontrado na testemunha.
Com relação à duração e sobrevivência da fase pupal, não houve diferenças significativas entre os tratamentos. A longevidade dos adultos apresentou diferenças significativas entre os genótipos PI 126931 (11,3 dias) e Andrea Victory (7,8 dias), sugerindo proporcionar, neste último, insetos menos longevos (Quadro 2).
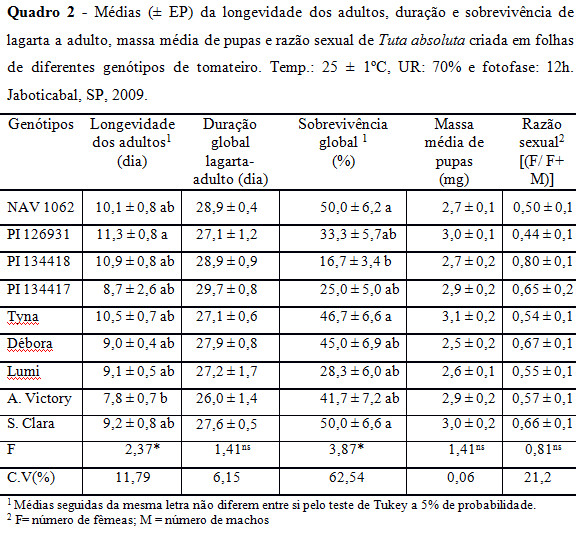
A sobrevivência global de T. absoluta criada nos genótipos ‘Santa Clara' (50,0%), NAV 1062 (50,0%) e Tyna (46,7%) diferenciaram-se das criadas em PI 134418 (16,7%) (Quadro 2). A duração do desenvolvimento desde a eclosão das lagartas até à emergência dos adultos (duração global), a massa de pupas e a razão sexual não obtiveram diferenças significativas entre os tratamentos, mostrando não terem sido afetados pelos genótipos (Quadro 2).
O genótipo PI 134418 foi, dentre os demais avaliados, o que apresentou efeito mais negativo sobre T. absoluta, alongando a fase larval e reduzindo as sobrevivências larval e global. Eigenbrode e Trumble (1993), também em um estudo sobre o mecanismo de resistência por antibiose, constataram alta mortalidade de S. exigua por L. hirsutum f. glabratum (PI 134418). Segundo Rosero (2010) essas espécies silvestres de tomateiro são importantes fontes de genes para resistência genética a insetos pragas.
O genótipo PI 134417 também prejudicou o desenvolvimento de T. absoluta, proporcionando um aumento da fase larval e reduzindo a sobrevivência larval, o que confirma a resistência deste material, também constatada por Thomazini et al. (2001), onde o genótipo alongou a fase larval e pupal e reduziu a massa de pupas fêmeas desta espécie. Giustolin e Vendramim (1994) também observaram a resistência deste genótipo, provocando alongamento das fases larval e pupal, redução da sobrevivência larval, menor massa de pupas e menor fecundidade das fêmeas de T. absoluta.
Os efeitos adversos sobre a praga indicam ser a antibiose um dos tipos responsáveis pela menor suscetibilidade das linhagens PI 134417 e PI 134418 de L. hirsutum f. glabratum. Leite (2004) relata que as causas que podem estar envolvidas nesta menor suscetibilidade devem estar associadas aos tricomas glandulares e não glandulares e àquelas associadas com lamela média, folha, frutos, hábito de crescimento ou idioblastos cristalíferos.
Além disso, a espécie L. hirsutum f. glabratum apresenta o aleloquímico 2-TD, substância considerada tóxica a vários insetos, o que prejudica, assim, o seu desenvolvimento (Linden, 1996). Gilardón et al. (2001) consideram apenas em parte o 2-TD responsável pela resistência do tomateiro à T. absoluta, considerando que alguma podem ter outras causas de resistência. No entanto, segundo Willians et al. (1980), Lin et al. (1987) e Pereira et al. (2000), essa substância tem sido citada como responsável pela resistência a várias pragas.
Fatores outros que também podem influenciar a suscetibilidade dos genótipos são a idade da planta e o estágio de desenvolvimento da folha, pois os mesmos podem proporcionar variações nas concentrações de 2-TD. Segundo Leite et al. (1999), a parte apical do dossel de L. hirsutum é menos atacada por lagartas de T. absoluta, e o teor de 2-TD aumenta da base para o ápice do dossel. Kennedy et al. (1981) relataram que, com o aumento da fotofase, ocorre acréscimo no teor de 2-TD.
Com relação aos genótipos de L. pimpinellifolium (NAV 1062 e PI 126931), constatou-se que não afetaram nenhum dos aspectos biológicos estudados, discordando dos relatos apresentados por Lourenção et al. (1984), em que NAV 1062 se destacou como o genótipo menos danificado (em comparação com genótipos de L. esculentum e L. peruvianum Mill).
Dentre os genótipos comerciais de tomateiro, Andrea Victory interferiu apenas na longevidade dos adultos, reduzindo-a, porém, não houve mais qualquer outro aspecto biológico em que esta se diferenciou da testemunha, sendo assim considerada suscetível ao ataque de T. absoluta, semelhante aos demais genótipos comerciais.
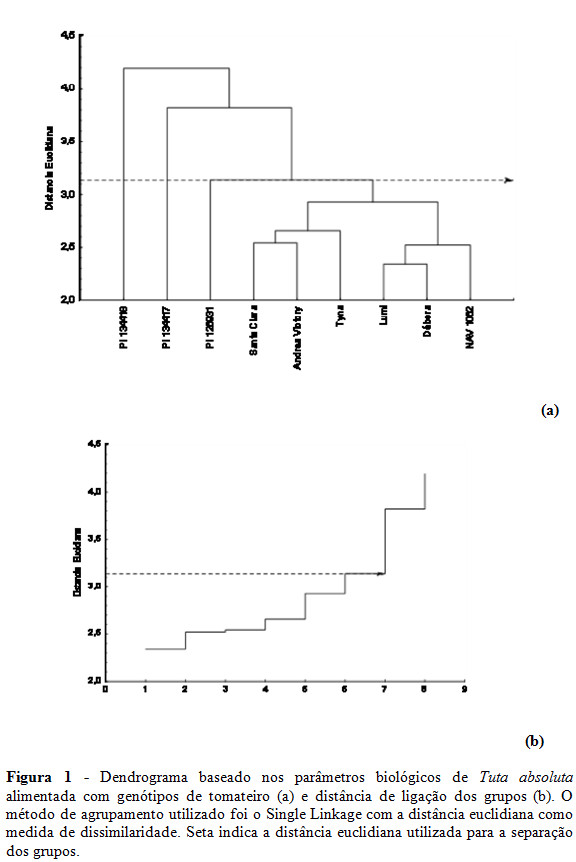
Por meio da análise de agrupamento hierárquico, observou-se que houve uma distinção entre os genótipos, dividindo-os em grupos de acordo com o grau de similaridade entre os mesmos (Figura 1). A partir do gráfico da distância de ligação dos grupos (Figura 1b), fixou-se a distância euclidiana em 3,14, sugerindo a divisão dos genótipos em quatro grupos distintos: o primeiro é formado pelo genótipo PI 134418; PI 134417 forma o segundo grupo; o terceiro grupo é formado pelo genótipo PI 126931; e os genótipos Santa Clara, Andrea Victory, Tyna, Lumi, Débora e NAV 1062 se isolam dos demais genótipos e formam um único grupo (Figura 1a).
Pela análise dos componentes principais, o primeiro componente principal (CP1) concentrou 40,4% da variabilidade contida nas variáveis originais, sendo os seguintes parâmetros aqueles que mais influenciaram esse componente principal: duração larval (0,93), sobrevivência larval (-0,88), duração global (0,76), e sobrevivência global (-0,83) (Figura 2a). O segundo componente principal (CP2) concentrou 22,4% da variabilidade presente nas variáveis originais, e os parâmetros que mais influenciaram o CP2 foram o período pupal (0,87) e sobrevivência pupal (0,78) (Figura 2a). Longevidade, razão sexual e massa de pupas foram os parâmetros biológicos que menos influenciaram na diferenciação e classificação dos genótipos de tomateiro quanto aos graus de resistência a T. absoluta (Figura 2a).
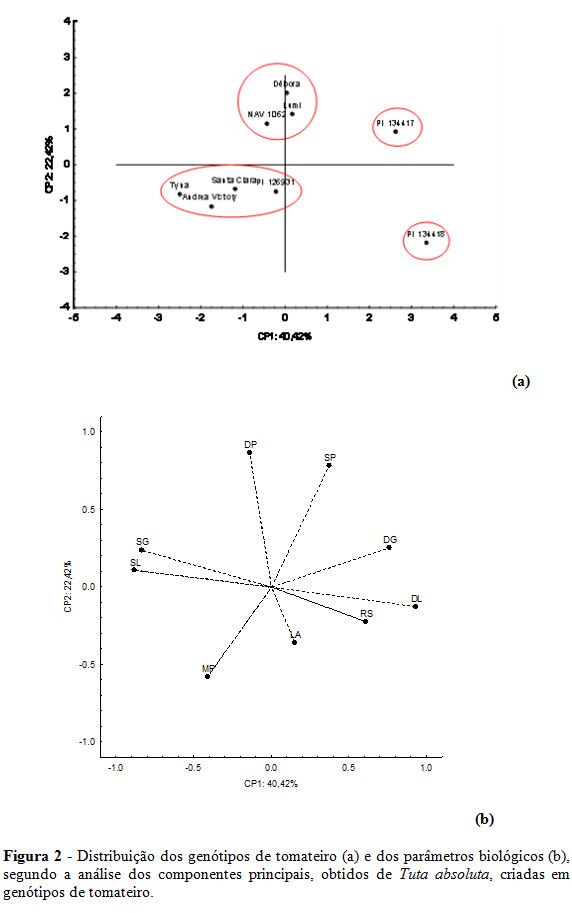
Os resultados obtidos por meio da análise dos componentes principais foram semelhantes aos obtidos pela análise de agrupamento hierárquica quanto ao número de grupos formados pelos genótipos de tomateiro (Figura 2a). Os genótipos PI 134418 e PI 134417 ficaram isolados dos demais, posicionando-se no 4º e 1º quadrantes, respectivamente (Figura 2a). No 3º quadrante, formou-se um grupo composto pelos genótipos PI 126931, Santa Clara, Andrea Victory e Tyna. Por fim, um quarto grupo foi constituído por Débora, Lumi e NAV 1062, posicionando-se próximo ao eixo do CP2 (Figura 2a).
Nota-se, no entanto, que houve algumas alterações quanto aos genótipos integrantes de cada grupo formado pela análise de componentes principais em relação à análise de agrupamento hierárquica (Figura 2a). Contudo, em ambas as análises multivariadas os genótipos PI 134418 e PI 134417 foram isolados individualmente dos demais por serem influenciados principalmente pelas maiores durações do período larval e menores sobrevivência larval e sobrevivência global (Figura 2a e 2b).
Bottega (2013), constatou que o genótipo PI134417 apresentou alto teor de tricomas glandulares, principalmente do tipo IV, e efeitos negativos sobre a duração larval e global, e baixa sobrevivência larval e global de T. absoluta. De acordo com Gonçalves et al. (2006) e Rodríguez-Lopes et al. (2011) os tipos de tricomas glandulares VI e IV em tomateiro, são os dois tipos mais citados como influência negativa a artrópodes praga.
Gonçalves et al. (2006) realizando testes para verificar a relação entre zingibereno, tricomas foliares e repelência a Tetranychus evansi (Koch), constataram que o zingibereno está associado especialmente aos tricomas tipo VI e mostram a sua maior presença na superfície abaxial dos folíolos dos genótipos de tomateiro avaliados. Esses dados concordam em parte com Freitas et al. (2002), que atribuíram a presença do sesquiterpeno aos tricomas glandulares, especialmente os de tipo VI e IV.
Baldin et al. (2005) constataram que PI134418 e PI134417 alongaram o ciclo de B. tabaci, indicando a ocorrência de não-preferência para alimentação e/ou antibiose. Constataram também que os tricomas glandulares desempenham papel fundamental para a não-fixação ou mortalidade do inseto na planta, devendo ser devidamente explorados nos programas de melhoramento, a fim de oferecer novas ferramentas para o controle de insetos-praga.
Dessa forma pode-se considerar que das linhagens de L. hirsutum f. glabratum são material muito interessante para introgressão de genes de resistência.
Conclusões
De maneira geral, conclui-se, a partir dos resultados obtidos nas análises de agrupamento hierárquica e dos componentes principais, que é possível classificar os genótipos de tomateiro da seguinte forma, de acordo com os graus de resistência do tipo antibiose demonstrado nesse experimento: PI 134418, altamente resistente, (AR); PI 134417, moderadamente resistente (MR); PI 126931, ‘Santa Clara', Tyna e Andrea Victory, suscetíveis (S); e Débora, Lumi e NAV 1062, altamente suscetíveis (AS).
Agradecimentos
Ao CNPq, pela concessão da bolsa de doutorado à primeira autora e de produtividade em pesquisa ao segundo autor, e a CAPES, pela concessão de bolsa de doutorado ao terceiro autor.
Referências Bibliográficas
Alvarenga, M.A.R. (2004) - Tomate: Produção em campo, em casa-de-vegetação e em hidroponia. Lavras, UFLA. 400 p. [ Links ]
Baldin, E.L.L.; Vendramim, J.D. e Lourencao, A.L. (2005) - Resistência de genótipos de tomateiro à mosca-branca Bemisia tabaci (Gennadius) biótipo B (Hemiptera: Aleyrodidae). Neotropical Entomology, vol. 34, n. 3, p. 435-441. [ Links ]
Bottega, D.B. (2013) - Resistência de genótipos de tomateiro a Tuta absoluta (Meyrick, 1917) (Lepidoptera: Gelechiidae) e efeito no comportamento e desenvolvimento de Podisus nigrispinus (Dallas, 1851) (Hemiptera: Pentatomidae). Tese de Doutorado. Jaboticabal, Universidade Estadual Paulista, Faculdade de Ciências Agrárias e Veterinárias de Jaboticabal. 83p. [ Links ]
Eeigenbrode, S.D. e Trumble, J.T. (1994) - Fruit-based tolerance to damage by beet armyworm (Lepidoptera: Noctuidae) in tomato. Environmental Entomology, vol. 23, n. 4, p. 937-942. [ Links ]
Eigenbrode, S.D. e Trumble. J.T. (1993) - Antibiosis to beet armyworm (Spodoptera exigua) in Lycopersicon acessions. HortScience, vol. 28, n, 9, p. 932-934. [ Links ]
Filgueira, F.A.R. (2005) - Novo manual de olericultura: agrotecnologia moderna na produção e comercialização de hortaliças. 2ª ed. Viçosa: Universidade Federal de Viçosa, 412 p. [ Links ]
Fornazier, M.J.; Dessaune Filho, N. e Pereira, E.B. (1986) - Perdas ocasionadas pela traça-do-tomateiro Scrobipalpula absoluta em diversas variedades de tomate. Cariacica: EMCAPA, Comunicado Técnico, 46 p. [ Links ]
Freitas, J.A.; Nonato, M.F.B.; Souza, V.S.; Maluf, W.R.; Ciociola Júnior, A.I.; Leite, G.L.D . (2002) - Relações entre acilaçúcares, tricoma glandular e resistência do tomateiro à mosca branca. Acta Scientiarum, vol.24, n. 5, p. 1313-1316. [ Links ]
Gilardón, E.; Pocovi, M.; Hernández, C.; Collavino, G. e Olsen, A. (2001) - Papel da 2-tridecanona e dos tricomas glandulares tipo VI na resistência do tomateiro a Tuta absoluta. Pesquisa Agropecuária Brasileira, vol. 36, n. 7, p. 929-933. [ Links ]
Giustolin, T.A. e Vendramim, J.D. (1994) - Efeito de duas espécies de tomateiro na biologia de Scrobipalpuloides absoluta (Meyrick). Anais da Sociedade Entomológica do Brasil, vol. 23, n. 3, p. 511-517. [ Links ]
Giustolin, T.A.; Vendramim, J.D. e Parra, J.R.P. (2002) - Number of larval instars of Tuta absoluta (Meyrick) in tomato genotypes. Scientia Agricola, vol. 59, n. 2, p. 393-396. [ Links ]
Gonçalves Neto, A.C.; Silva, V.F.; Maluf, W.R.; Maciel, G.M.; Nizia, A.C.D.; Gomes, L.A.A. e Azevedo, S.M. (2010) - Resistência à traça-do-tomateiro em plantas com altos teores de acilaçúcares nas folhas. Horticultura Brasileira, vol. 28, n. 2, p. 203-208. [ Links ]
Gonçalves, L.D.; Maluf, W. R.; Cardoso, M. G.; Resende, T. V. R.; Castro, E. M.; Santos, N. M.; Nascimento, I. R.; Faria, M. V (2006) - Relação entre zingibereno, tricomas foliares e repelência de tomateiros a Tetranychus evansi. Pesquisa Agropecuária Brasileira, Brasília, vol. 41, n. 2, p. 267-273. [ Links ]
Kennedy, G.G.; Yamamoto, R.T.; Dimock, M.B.; Williams, W.G. e Bordner, J. (1981) - Effect of daylength and light intensity on 2-tridecanone levels and resistance levels in Lycopersicon hirsutum f. glabratum to Manduca sexta. Journal of Chemical Ecology, vol. 7, n. 4, p. 707-716. [ Links ]
Lara, F.M. (1991) - Princípios de resistência de plantas a insetos. 2a. ed. São Paulo, Ícone. 336p. [ Links ]
Leite, G.L.D. (2004) - Resistência de tomates a pragas. Unimontes Científica, vol. 6, n. 2, p. 129-140. [ Links ]
Leite, G.L.D.; Costa, C.A.; Almeida, C.I.M. e Picanço, M. (2003) - Efeito da adubação sobre a incidência de traça-do-tomateiro e alternaria em plantas de tomate. Horticultura Brasileira, vol. 21, n.3, p. 448-451. [ Links ]
Leite, G.L.D.; Picanço, M.; Azevedo, A.A. e Gonring, A.H.R. (1999) - Efeito de tricomas, aleloquímicos e nutrientes na resistência de Lycopersicon hirsutum à traça-do-tomateiro. Pesquisa Agropecuária Brasileira, vol. 34, n.11, p. 2059-2064. [ Links ]
Lin, S.Y.H.; Trumble, J.T. e Kumamoto, J. (1987) - Activity of volatile compounds in glandular trichomes of Lycopersicon species against two insect herbivores. Journal of Chemical Ecology, vol. 13, n. 4 p. 837-850. [ Links ]
Linden, A. (1996) - Control of carterpillars in integrated pest management. IOBC/WRPS Bulletin, vol. 19, n. 1, p. 91-94. [ Links ]
Lourenção, A.L.; Nagai, H. e Zuloo, M.A.T. (1984) - Fontes de resistência a scrobipalpula absoluta (Meyrick, 1917) em tomateiro. Bragantia, vol. 43, n. 2, p. 569-577. [ Links ]
Miranda, M.M.M.; Picanço, M.C.; Zanuncio, J.C. e Guedes, R.N.C. (1998) - Ecological life table of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Biocontrol Science and Technology, vol. 8, n.4, p. 597-606. [ Links ]
Pereira, N.E.; Leal, N.R. e Pereira, M.G. (2000) - Controle genético da concentração de 2-tridecanona e de 2-undecanona em cruzamentos interespecíficos de tomateiro. Bragantia, vol. 59, n.2, p. 165-172. [ Links ]
Rodríguez-Lópes, M. J.; Garzo, E.; Bonani, J.P.; Fereres, A.; Fernández-Muñoz, R. e Moriones, E. (2011) - Whitefly resistance traits derived from the wild tomato Solanum pimpinellifolium affect the spread of tomato yellow leaf curl vírus. Phytopathology, vol. 101, n. 10, p. 1191-1201. [ Links ]
Silva, A.C. e Carvalho, G.A. (2004) - Manejo Integrado de Pragas. In: AVARENGA, M.A.R. (Ed.) - Tomate: produção em campo, em casa-de-vegetação e em hidroponia. Lavras, UFLA, p. 309-366. [ Links ]
Siqueira, H.A.A.; Guedes, R.N.C.; Fragoso, D.B. e Magalhães, L.C. (2001) - Abamectin resistance and synergism in Brazilian populations of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). International Journal of Pest Management, vol. 47, n. 4, p. 247-251. [ Links ]
Statistica Statsoft. (2004) - Data Analysis Software System and User's Manual. Version 7. StatSoft Inc. [ Links ]
Suinaga, F.A.; Picanço, M.C.; Moreira, M.D.; Semeão, A.A. e Magalhães, S.T.V. (2004) - Resistência por antibiose de Lycopersicon peruvianum à traça do tomateiro. Horticultura Brasileira, vol. 22, n. 2, p. 281-285. [ Links ]
Thomazini, A.P.B.W.; Vendramim, J.D.; Brunherotto, R. e Lopes, M T.R. (2001) - Efeito de genótipos de tomateiro sobre a biologia e oviposição de Tuta absoluta (Meyrick) (Lep.: Gelechiidae). Neotropical Entomology, vol.30, n.2, p. 283-288. [ Links ]
Recebido/Received: 2014.08.16
Aceite/Accepted: 2015.02.27

