










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias vol.38 no.3 Lisboa set. 2015
ARTIGO
Óxido nítrico na germinação de sementes de baixo vigor de Dalbergia nigra
Nitric oxide on the germination of Dalbergia nigra seeds with low vigor
Glauciana M. Ataíde1*, Eduardo E.L. Borges2, Andressa V. Flores3 e Renato V. O. Castro4
1 Universidade Federal Rural do Rio de Janeiro, Instituto de Florestas, BR 465, km 07, CEP 23890-000, Seropédica, RJ, Brasil. E-mail: *glaucianadamata@yahoo.com.br, author for correspondence.
2 Universidade Federal de Viçosa, Departamento de Engenharia Florestal, Avenida pH Rolfs, s/n, CEP 36570-000, Viçosa, MG, Brasil.
3 Universidade Federal de Santa Catarina, Departamento de Engenharia Florestal, Rodovia Ulisses Gaboardi, Km 3, CEP 89520-000, Curitibanos, SC, Brasil.
4 Universidade Federal de São João Del-Rei, Departamento de Ciências Agrárias, Rodovia MG 424, km 47, CEP 35701-970, Sete Lagoas, MG, Brasil.
RESUMO
O óxido nítrico é um composto que participa de inúmeros processos fisiológicos vegetais, dentre os quais atua estimulando a germinação tanto sob condições normais quanto sob estresse. O presente trabalho teve como objetivo investigar a eficiência de nitrato de potássio (KNO3) e de nitroprussiato de sódio (SNP), formadores de óxido nítrico, na germinação de sementes de baixo vigor de Dalbergia nigra, de modo a determinar as melhores condições de condicionamento das sementes da espécie. As sementes foram pré-embebidas em água ou nas soluções de KNO3 ou SNP nas seguintes concentrações: 0,01; 0,1; 1 e 10 mmol L-1, por períodos de tempo de seis, 12, 24 e 48-h. Em seguida, foram conduzidos testes de germinação em câmaras de germinação a 25 °C durante 12 dias, estimando-se a porcentagem de germinação pela contagem diária das sementes que emitiram radícula e o índice de velocidade de germinação (IVG). Nas sementes pré-embebidas em soluções de SNP e KNO3, as percentagens de germinação nas concentrações de 0,01 e 0,1 mmol L-1foram significativamente superiores às outras concentrações e à testemunha, que apresentou germinação média de 48%. Conclui-se que a aplicação de KNO3 e SNP aumentou a germinação e o vigor das sementes de Dalbergia nigra de baixo vigor.
Palavras-chave: jacarandá-da-Bahia, nitrato de potássio, nitroprussiato de sódio, sementes florestais
ABSTRACT
Nitric oxide is a compound that participates in many plant physiological processes, among which works by stimulating the germination both under normal and under stress conditions. This study aimed to investigate the effectiveness of potassium nitrate (KNO3) and sodium nitroprusside (SNP), forming nitric oxide, on the germination of seeds of low vigor of Dalbergia nigra, to determine the best conditions for the conditioning of the seeds of the species in these solutions. The seeds were pre-soaked in water or in solutions of KNO3 or SNP at the following concentrations: 0.01, 0.1, 1 and 10 mmol L-1, for periods of six, 12, 24 and 48 hours. Then, tests were conducted in germination chamber at 25 °C for 12 days, calculating the percentage of germination of seeds by counting daily that issued radicle and germination speed index (GSI). In pre-soaked seeds in solutions of KNO3 and SNP, the seed germination at concentrations of 0.1 and 1 mmol L-1was higher than the others, and also higher than the control, that had averaged 48% seed. It is concluded that the application of KNO3 and SNP increased seed germination and seed vigor of Dalbergia nigra seeds.
Keywords: forest seeds, jacaranda-da-bahia, potassium nitrate, sodium nitroprusside
Introdução
A conservação da biodiversidade dos ecossistemas florestais tropicais apresenta-se como uma das principais preocupações da humanidade nas últimas décadas. Em decorrência da devastação florestal para a extração de madeira, tem sido observado uma regressão do potencial genético de muitas espécies, levando inclusivé à extinção de diversas espécies florestais nativas e colocando em risco a sobrevivência dos ecossistemas (Kaviani, 2011).
Desta forma, a produção de sementes de espécies florestais ganhou grande importância em programas de reposição florestal, reflorestamento, recuperação de áreas degradadas, arborização urbana, entre outras atividades. A qualidade fisiológica, genética e física das sementes é fator preponderante para o sucesso dos programas de produção das mudas a serem utilizadas para os diversos fins.
O vigor das sementes é definido como o conjunto de características que determinam o seu potencial fisiológico, ou seja, conferem à semente o potencial para germinar, emergir e resultar rapidamente em plântulas normais, quando expostas a diferentes condições ambientais (Marcos Filho, 2005). Sementes de baixo vigor são caracterizadas por apresentarem menor velocidade nos processos metabólicos, propiciando emissão mais lenta e desigual da raiz primária no processo de germinação, menores taxas de crescimento e plântulas com menor tamanho inicial (Schuch et al., 1999).
O condicionamento osmótico consiste na pré-embebição das sementes em soluções com potencial osmótico conhecido, permitindo assim, a ocorrência das fases iniciais do processo de germinação (fases I e II), sem, contudo, atingir o estado de emergência da raiz primária (fase III) (Bewley et al., 2013). Dessa forma, o condicionamento osmótico permite às sementes se hidratarem lentamente, resultando em acréscimos na taxa e uniformidade de germinação.
O óxido nítrico (NO) é um radical livre, incolor, inorgânico, sintetizado a partir da L-arginina por ação da óxido nítrico sintetase (NOS) (Beligni e Lamattina, 1999; Leite e Sarni, 2003), sendo a sua participação evidenciada em inúmeros processos fisiológicos vegetais, entre os quais: função protetora contra o estresse oxidativo (Wink et al., 1993), antioxidante por impedir reações de peroxidação de lipídios decorrentes de déficit hídrico (Cheng et al., 2002) e estresse salino (Uchida et al., 2002), e regulação nas respostas de defesa da planta à infecção por patógenos (Noritake et al., 1996).
Em sementes, o NO estimula a germinação tanto sob condições normais quanto sob estresse (Kopyra e Gwóźdź, 2003; Souza, 2007; Zheng et al., 2009), favorece a quebra de dormência de sementes de algumas espécies (Bethke et al., 2006; Renata e Agnieszka, 2006) e atua promovendo o alongamento e formação das raízes adventícias (Beligni e Lamattina, 2001).
Os estudos sobre os efeitos do NO na fisiologia das plantas, apesar de recentes, têm-se concentrado principalmente em definir a aplicação exógena de doadores do NO numa ampla gama de concentrações e tempos (Beligni e Lamatina, 2001), sendo as substâncias mais utilizadas o nitroprussiato de sódio (SNP), o nitrato de potássio (KNO3), o S-nitroso-Nacetilpenicilamino (SNAP) e o S-Nitrosoglutationa (GSNO). Assim, sua influência nas alterações fisiológicas e bioquímicas decorrentes do envelhecimento das sementes, assim como possíveis meios para reverter as baixas taxas de germinação das sementes deterioradas são um novo e estimulante campo de estudos da biologia vegetal.
A espécie jacarandá-da-Bahia (Dalbergia nigra (Vell.) Allemão ex Benth.) pertence à família Fabaceae (Caesalpinoidea) e ocorre nos biomas Mata Atlântica e no cerrado brasileiros, especialmente nos Estados de Minas Gerais, São Paulo e Mato Grosso do Sul. É uma espécie tardia a clímax, recomendada para arborização de pastos ou para arborização de culturas, e a sua madeira é muito usada em de móveis de luxo (Lorenzi, 2009). Atualmente, encontra-se na lista das espécies da flora brasileira ameaçadas de extinção, na categoria vulnerável (MMA, 2008).
Vários trabalhos têm sido desenvolvidos com sementes da espécie, tais como a avaliação do seu comportamento germinativo durante o armazenamento (Aguiar et al., 2010), a definição da metodologia adequada para a condução do teste de condutividade elétrica (Marques et al., 2002), a cinética de embebição das sementes (Ataíde et al., 2014) e a mobilização de reservas e atividade enzimática durante a germinação (Ataíde et al., 2013a,b).
Face à relevância de tais estudos para a germinação de sementes da espécie e à escassez de mais informações com sementes de espécies florestais, este trabalho teve como objetivo avaliar a eficiência de nitrato de potássio (KNO3) e de nitroprussiato de sódio (SNP), formadores de óxido nítrico, na germinação de sementes de baixo vigor de Dalbergia nigra, com a finalidade de determinar as melhores condições para o condicionamento osmótico da espécie nestas soluções.
Materiais e métodos
O presente trabalho foi conduzido no Laboratório de Sementes Florestais do Departamento de Engenharia Florestal da Universidade Federal de Viçosa (UFV). Foram utilizadas sementes de Dalbergia nigra colhidas em várias árvores na região de Viçosa, Minas Gerais. Durante a limpeza foram eliminadas as sementes imaturas, deterioradas ou danificadas. As sementes selecionadas foram acondicionadas em tambores de fibra e armazenadas em câmara fria a 5 °C e 60% humidade relativa do ar, durante duas semanas, até a realização dos testes.
De seguida, as sementes de D. nigra foram colocadas sobre duas folhas de papel germtest em placas de vidro e pré-embebidas em água ou em 14 mL de soluções de nitrato de potássio (KNO3) ou nitropussiato de sódio (SNP) nas concentrações de 0,01, 0,1, 1 e 10 mmol L-1, por seis, 12, 24 e 48-h à temperatura de 25 °C, sob luz constante. As placas foram vedadas com fita crepe para evitar a evaporação das soluções.
Após cada tempo de pré-embebição, as sementes foram retiradas das soluções e colocadas para germinar em placas de Petri com 9 cm de diâmetro, forradas duplamente com papel de filtro do tipo germtest umedecido com água destilada. Essas placas foram mantidas em câmara de germinação a 25°C e sob luz constante durante 12 dias. As sementes pré-embebidas em água foram empregadas como testemunha.
Foram utilizadas 5 repetições de 20 sementes, sendo a germinação determinada pela contagem diária das sementes que emitiram radícula (> 2 mm), e os resultados expressos em porcentagem média. Para o cálculo do Índice de Velocidade de Germinação (IVG), foi utilizada a fórmula proposta por Maguire (1962):
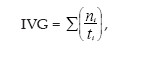
onde, ni é o número de sementes por dia e ti é o tempo (dias).
O delineamento experimental utilizado foi inteiramente casualizado (DIC). Os resultados foram submetidos à análise de variância (ANOVA), sendo a comparação entre os diferentes tratamentos feita pelo teste de Tukey a 5% de significância. As médias dos tratamentos de pré-embebição nas soluções de SNP e KNO3 foram comparadas às médias da testemunha (sementes pré-embebidas em água) pelo teste de Dunett, também a 5% de significância. O software utilizado foi o Statistica 8.0 (Statsoft, 2008).
Resultados e discussão
As sementes de Dalbergia nigra colocadas a germinar em água (testemunha) apresentaram porcentagem de germinação média de 48% e IVG médio de 1,30, valores considerados como de baixo vigor numa avaliação de qualidade de sementes (Bonner, 1998), considerando que as sementes foram colhidas pouco antes da condução do teste de germinação.
Na Figura 1 são apresentadas as médias de germinação das sementes colocadas em embebição nas soluções de SNP. Os maiores valores de germinação foram observados nas concentrações de 0,01 e 0,1 e mmol L-1, que não diferiram entre si, mas foram estatisticamente superiores às outras concentrações, com exceção no tempo de 48-h, no qual estas foram iguais estatisticamente à concentração 1 mmol L-1. Dentro de cada concentração utilizada, somente foram observadas médias significativamente diferentes entre os tempos de embebição na concentração de 1 mmol L-1, onde a porcentagem de germinação após embebição por 48-h foi semelhante à embebição por 24-h e superior aos tempos de 6 e 12-h. Em relação à testemunha, valores significativamente superiores foram observados para as concentrações 0,01 e 0,1 mmol L-1nos tempos de 12, 24 e 48-h de embebição e na concentração 1 mmol L-1em 48-h de embebição. As maiores percentagens alcançadas corresponderam a 73 e 74 % de germinação após embebição por 48-h nas concentrações 0,01 e 0,1 mmol L-1, respectivamente. Tais resultados representam acréscimos de 25 e 26% na germinação inicial. Dessa forma, a pré-embebição das sementes de D. nigra em soluções de SNP durante maiores períodos de tempo alterou positivamente a capacidade germinativa das sementes da espécie.
Segundo Grubíšic et al. (1992), o nitroprussiato de sódio (SNP) e outros nitratos na sua forma orgânica e inorgânica foram capazes de estimular a germinação de sementes em concentrações ótimas variando entre 1 e 10 mM. Estes autores sugerem que os efeitos positivos estão relacionados à redução do SNP para ON. Apesar dos trabalhos recentes com SNP abrangerem apenas algumas espécies, tem sido observado que esse doador atua tanto na redução da dormência das sementes, quanto no aumento dos efeitos da luz sobre a germinação ou na porcentagem final de germinação de sementes não dormentes (Bethke et al., 2007).
Concentrações de SNP de 0,1 e 1,0 mmol L-1 também estimularam a germinação de sementes de Vigna unguiculata (L.) Walp e Phaseolus vulgaris L. submetidas ao estresse hídrico, as quais apresentaram acréscimos entre 26 e 36% na germinação final (Zimmer-Prados, 2007). Resultados semelhantes foram encontrados por Gniazdowska et al. (2010), onde a porcentagem de germinação de Malus domestica Borkh.,inicialmente de 5% sem aplicação do tratamento com SNP, atingiu 64% após a imersão dos embriões na solução por 3-h, na concentração de 5 mM.
As sementes submetidas a pré-embebição nas soluções de KNO3 apresentaram comportamento semelhante às sementes embebidas na solução de SNP, de forma que as concentrações 0,01 e 0,1 mmol L-1 proporcionaram maiores percentagens de germinação (Figura 2). Para as demais concentrações de KNO3, os tempos avaliados não diferiram entre si, apesar de as maiores médias de germinação ocorrerem nos tempos de 24 e 48-h. Após 48-h de pré-embebição, as sementes apresentaram germinação de 83 e 85% nas concentrações 0,01 e 0,1 mmol L-1, respectivamente. Estes valores, significativamente superiores à testemunha, implicam incrementos de 35 e 37% nas percentagens iniciais de germinação.
A eficácia do nitrato de potássio em promover a germinação é exercida através da sua redução a nitrito, hidroxilamina ou óxido nítrico (NO) (Hendricks e Taylorson, 1974). Segundo Nonogaki et al. (2010), o NO é reconhecido como um componente importante de transdução de sinal, e pode acelerar o fluxo metabólico através da via das pentoses fosfatadas, aumentando indiretamente a oxidação de NADPH, embora possa estimular a germinação por outras vias, tais como aquelas envolvidas no metabolismo hormonal.
Em Arabidopsis, o NO pode atuar promovendo a biossíntese de giberelina (GA) (Bethke et al., 2007), ou por meio da desativação do áciso abscísico (ABA) (Matakiadis et al., 2009). Sirovà et al. (2011) destacam ainda que a aplicação de nitrato e SNP afeta significativamente o fitocromo a específico de germinação, enquanto o fitocromo b é pouco influenciado.
Os Quadros 1 e 2 apresentam as médias de IVG para as sementes embebidas nas soluções de SNP e KNO3, respectivamente.
Na embebição das sementes com SNP foi observada uma tendência de acréscimo nos valores de IVG à medida em que se aumentou o tempo de exposição das sementes à solução proposta. Dessa forma, sementes embebidas durante 48-h apresentaram médias superiores de IVG em todas as concentrações de SNP avaliadas.
Pereira et al. (2010) avaliando a influência do óxido nítrico na germinação de sementes de Plathymenia reticulata Benth. com baixo vigor, constataram que a partir de 12-h de embebição em soluções de SNP e KNO3, todos os tratamentos diferiram significativamente da testemunha.
No presente trabalho, as médias de IVG após pré-embebição em SNP por 24-h na concentração 0,01 mmol L-1 e por 48-h em todas as concentrações testadas foram estatisticamente superiores à testemunha.
Semelhante à pré-embebição em SNP, a embebição das sementes em solução de KNO3 por períodos de 48-h foi a que apresentou maiores valores de IVG, em todas as concentrações. Nas concentrações 0,1 e 1 mmol L-1, as médias de IVG em 24-h não diferiram das médias de IVG após embebição durante períodos de 48-h, indicando que nestas concentrações a embebição das sementes em KNO3 a partir de 24-h já é significativamente benéfica para estimular a germinação das sementes de Dalbergia nigra.
Quando aplicado exogenamente, o NO estimulou a germinação de sementes de alface (Lactuca sativa L.) no escuro (Beligni e Lamattina, 2001), a germinação de sementes de Paulownia tomentosa (Thunb.) Steud.,(Giba et al., 1998) e a germinação e crescimento radicular de tremoço (Lupinus luteus L.) (Kopyra e Gwóźdź, 2003) e induziu a quebra de dormência em sementes de Arabidopsis e cevada (Bethke et al., 2006).
Conclusões
As aplicações de diferentes concentrações e tempos de SNP e KNO3 estimularam a germinação e o IVG de sementes de Dalbergia nigra, porém resultados mais expressivos foram observados nas concentrações de 0,01 e 0,1 mmol L-1 de ambas as soluções, especialmente em períodos de embebição de 24 e 48 horas.
Agradecimentos
Os autores agradecem a Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG) pelo apoio financeiro.
Referências bibliográficas
Aguiar, F.F.A.; Tavares, A.R.; Kanashiro, S.; Luz, P.B. e Santos Júnior, N.A. (2010) - Germinação de sementes de Dalbergia nigra (Vell.) Allemao ex Benth. (Fabaceae-Papilionoideae) no armazenamento. Ciência e Agrotecnologia, vol. 34, Edição Especial, p. 1624-1629. [ Links ]
Ataíde, G.M.; Borges, E.E.L.; Flores, A.V. e Castro, R.V.O. (2014) - Avaliação preliminar da embebição de sementes de jacarandá-da-bahia. Pesquisa Florestal Brasileira, vol. 34, n. 78, p. 133-139. [ Links ]
Ataíde, G.M.; Borges, E.E.L.; Goncalves, J.F.C.; Guimaraes, V.M.; Bicalho, E.M. e Flores, A.V. (2013a) - Activities of α-galactosidase and polygalacturonase during hydratation of Dalbergia nigra ((Vell.) FR ALL. ex Benth.) seed at different temperatures. Journal of Seed Science, vol. 35, n. 1, p. 92-98.
Ataíde, G.M.; Borges, E.E.L.; Goncalves, J.F.C.; Guimaraes, V.M.; Flores, A.V. e Bicalho, E.M. (2013b) - Alterations in seed reserves of Dalbergia nigra ((Vell.) Fr All. ex Benth.) during hydration. Journal of Seed Science, vol. 35, n. 1, p. 56-63. [ Links ]
Beligni, M.V. e Lamattina, L. (1999) - Nitric oxide protects against cellular damage produced by methylviologen herbicides in potato plants. Planta, vol. 3, p. 199-208. [ Links ]
Beligni, M.V. e Lamattina, L. (2001) - Nitric oxide in plants: the history is just beginning. Plant, Cell and Environment, vol. 24, p. 267-278. [ Links ]
Bethke, P.C.; Libourel, I.G.L. e Jones, R.L. (2006) - Nitric oxide reduces seed dormancy in Arabidopsis. Journal of Experimental Botany, vol. 57, n. 3, p. 517-526. [ Links ]
Bethke, P.C.; Libourel, I.G.L.; Aoyama, N.; Chung, Y.; Still, D.W e Jones, R.L. (2007) - The Arabidopsis aleurone layer responds to nitric oxide, gibberellin and abscisic acid and is sufficient and necessary for seed dormancy. Plant Physiology, vol. 143, n. 3, p. 1173-1188. [ Links ]
Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M. e Nonogaki, H. (2013) - Seeds: physiology of development, germination and dormancy. Nova York: Springer, 392 p. [ Links ]
Bonner, F.T. (1998) - Testing tree seeds for vigor: a review. Seed Technology, vol. 20, n. 1, p. 5-17. [ Links ]
Cheng, F.; Hsu, S. e Kao, C.H. (2002) - Nitric oxide counteracts the senescence of detached rice leaves induced by dehydration and polyethylene glycol but not by sorbitol. Plant Growth Regulation, vol. 38, n. 3, p. 265-272. [ Links ]
Giba, Z.; Grubisic, D.; Todorovic, S.; Sajc, L.; Stojakovic, D.; Konjevic, T.(1998) - Effect of nitric oxide-releasing compounds on phytochrome-controlled germination of Empress tree seeds. Plant Growth Regulation, vol. 26, n. 3, p. 175-181. [ Links ]
Gniazdowska, A.; Krasuska, U. e Bogatek, R. (2010) - Dormancy removal in apple embryos by nitric oxide or cyanide involves modifications in ethylene biosynthetic pathway. Planta, vol. 232, n. 6, p. 1397-1407. [ Links ]
Grubíšic, D.; Giba, Z. e Konjevic, R. (1992) - The effect of organic nitrates in phytochrome-controlled germination of Paulownia tormentosa seeds. Photochemistry and Photobiology, vol. 56, n. 5, p. 629-632.
Hendricks, S.B. e Taylorson, R.B. (1974) - Promotion of seed germination by nitrate, nitrite, hydroxylamine, and ammonium salts. Plant Physiology, vol. 54, n. 3, p. 304-309. [ Links ]
Kaviani, B. (2011) - Conservation of plant genetic resources by cryopreservation. Australian Journal of Crop Science, vol. 5, n. 6, p. 778-800. [ Links ]
Kopyra, M. e Gwóźdź, E.A. (2003) - Nitric oxide stimulates seed germination and counteracts the inhibitory effect of heavy metals and salinity on root growth of Lupinus luteus.Plant Physiology and Biochemistry, vol. 41, n. 11-12, p. 1011-1017. [ Links ]
Leite, H.P. e Sarni, R.S. (2003) - Radicais livres, antioxidantes e nutrição. Revista Brasileira de Nutrição Clínica, vol. 18, n. 2, p. 87-94. [ Links ]
Lorenzi, H. (2009) - Árvores brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. Nova Odessa: Plantarum. [ Links ]
Maguire, J.D. (1962) - Speed of germination: aid in selection and evaluating or seedling emergence and vigour. Crop Science, vol. 2, n. 2, p. 176-177. [ Links ]
Marcos Filho, J.M. (2005) - Fisiologia de sementes de plantas cultivadas. Piracicaba: FEALQ, 495 p. [ Links ]
Marques, M.A.; Paula, R.C. e Rodrigues, T.J.D. (2002) - Adequação do teste de condutividade elétrica para determinar a qualidade fisiológica de sementes de jacarandá-da-bahia (Dalbergia nigra (Vell.) Fr. All. ex Benth.). Revista Brasileira de Sementes, vol. 24, n. 1, p. 271-278. [ Links ]
Matakiadis, T.; Alboresi, A.; Jikumaru, Y.; Tatematsu, K.; Pichon, O. e Renou J.P. (2009) - The Arabidopsis abscisic acid catabolic gene CYP707A2 plays a key role in nitrate control of seed dormancy. Plant. Physiology, vol. 149, n. 2, p. 949-960. [ Links ]
Recebido/Received: 2014:12:03
Aceite/Accepted: 2015.04.30

