










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias vol.39 no.3 Lisboa set. 2016
https://doi.org/10.19084/RCA15117
ARTIGO
Influência das condições microclimáticas no crescimento do milho BR 106, cultivado sob sementeira direta
Influence of microclimatic conditions in growth analysis of maize BR 106, cultivated under no-tillage
Marcos A. V. Silva1*, Williams M. P. Ferreira2, Vanda M. S. de Andrade3 e José M. N. da Costa4
1Universidade do Estado da Bahia, Departamento de Ciências Humanas Campus 9. BR 242, km 04, Loteamento Flamengo. CEP 47800000 - Barreiras, BA – Brasil. *E-mail: maavsilva@uneb.br
2Empresa Brasileira de Pesquisa Agropecuária, Centro Nacional de Pesquisa do Café. Vila Giannetti, casa 46, Campus UFV, Centro. CEP 36570000 - Viçosa, MG – Brasil;
3Universidade Federal Rural da Amazônia, Colegiados. Acesso do Tucunduba. Terra Firme. CEP 66077665 - Belém, PA – Brasil;
4Universidade Federal de Viçosa, Centro de Ciências Agrárias, Departamento de Engenharia Agrícola. Av. P. H. Rolfs s/n, Campus Universitário. CEP 36571000 - Viçosa, MG – Brasil.
RESUMO
Foi desenvolvida uma pesquisa no CNPMS (Embrapa) objetivando quantificar a influência das variáveis micrometeorológicas no crescimento e desenvolvimento do milho BR 106 cultivado via sementeira direta e condições de rega. Foi utilizado um anemómetro sónico tridimensional para a medição dos fluxos de CO2 acima da canópia. O crescimento do BR 106 foi afetado pelas variações do microclima da canópia, com grandes períodos de nebulosidade, diminuição do potencial de vapor na atmosfera e maior conteúdo de energia, principalmente luz e temperatura do ar, contribuindo para os valores máximos dos atributos de crescimento no período P2 (Florescimento). Contudo, a senescência das folhas do terço médio inferior foi mais expressiva na transição do P2 para o P3 (Embonecamento), houve aumento da razão da área foliar e da área foliar específica, além de redução da razão de massa foliar, que induziram a queda de 13% na assimilação do CO2 no período reprodutivo. Esse comportamento não afetou o rendimento final da variedade que apresentou um valor próximo do potencial máximo.
Palavras-chaves: Índice de área foliar, micrometeorologia, taxa de crescimento da cultura.
ABSTRACT
Research was conducted in CNPMS (Embrapa), Sete Lagoas-MG, to quantify the influence of micrometeorological variables on growth and development in the canopy of a maize crop variety BR 106 under no-tillage in irrigated conditions. The flow of CO2 was measured above the canopy using a system that comprises a three-dimensional sonic anemometer. The growth of BR 106 was affected by changing in the canopys microclimate, with extended periods of cloudiness, reducing the vapor potential in the atmosphere and higher content of energy, especially light and air temperature, contributing to the maximum values of the attributes of growth present in P2 (12 leaves to flowering). However, the senescence of lower leaves of canopy, in the transition from P2 to P3 (beginning of grain formation), increased the RLA (relation leaf area) and SLA (specific leaf area), and reduce the RLM (rate leaf matter), dependent on LAI (leaf area index) and NAR (net assimilation rate), which led to a decrease of 13% the assimilation of CO2 over the reproductive period. This behavior did not affect the final yield of the variety that showed a value close to potential.
Keywords: Crop growth rate, leaf area index, micrometeorology.
Introdução
Em função do valor nutritivo, o milho (Zea mays L.) é um dos cereais mais difundidos no mundo, assumindo grande importância social e económica. No Brasil, com cerca de 15,7 milhões de hectares, é a cultura mais largamente semeada, representando 29% da área agrícola do país (Conab, 2015). Segundo Fancelli (2003) o milho é considerado uma espécie com grande eficiência na conversão da energia radiante e, consequentemente na produção de biomassa, já que a semente contém, em média 0,260 g, após um período de tempo próximo a 135 dias, resulta em cerca de 0,8 a 1,25 kg de biomassa por planta e 180 a 250 g de grãos por planta, multiplicando, assim, mil vezes o peso da semente que a originou. Essa capacidade de desenvolvimento e produção é devida principalmente, à elevada acumulação de fotoassimilados que o milho apresenta, em função do seu mecanismo fotossintético tipo C4. Porém, a fotossíntese e os demais processos fisiológicos e bioquímicos responsáveis pelo desempenho da planta estão intimamente ligados aos fatores abióticos água, luz, temperatura e nutrientes, que devem, a priori, se apresentar em quantidades e valores adequados durante todo o ciclo da cultura, para que a planta expresse o seu máximo potencial produtivo.
Segundo Kunz et al. (2007), a radiação solar vem-se destacando nas pesquisas dos últimos anos, que buscam explorar o rendimento potencial das culturas. Embora a temperatura e o fotoperíodo sejam os principais fatores que atuam sobre o desenvolvimento vegetal, do ponto de vista quantitativo e qualitativo, a radiação solar é fundamental para o desenvolvimento e o crescimento vegetal, realizado pelos processos fotomorfogenéticos e fotossintéticos (Stewart et al., 2003). Assim, Müller e Bergamachi (2005) citam que a maneira como a radiação fotossinteticamente ativa é intercetada pelo dossel das plantas é fundamental para a fotossíntese e para a produção da cultura.
Na análise de crescimento são utilizadas diferentes equações matemáticas para avaliar índices de crescimento das plantas, muitos deles relacionados com a atividade fotossintética. Nos estudos ecofisiológicos das plantas, não se pode prescindir da análise de crescimento, pois os fatores ambientais e a disponibilidade de água e nutrientes, próprios de cada local, afetam sensivelmente a taxa assimilatória líquida e de crescimento relativo, a razão de área foliar além de outras variáveis das plantas. Por meio do estudo das interações dessas variáveis com os fatores ambientais, e em particular, com o estádio de desenvolvimento da planta, torna-se possível a identificação da eficiência de crescimento e a habilidade de adaptação às condições ambientais em que as plantas se desenvolvem.
As plantas apresentam resposta estomática diretamente influenciada pelas condições ambientais modificadas pelo sistema de rega, sendo essa resposta também condicionada pela variação dos elementos micrometeorológicos como: radiação fotossinteticamente ativa, vento, concentração de CO2 no ambiente e na câmara estomática, o cultivo irrigado tende a apresentar valores diferenciados em termos de interação gasosa, variáveis micrometeorológicas e indicadores de crescimento. Assim, a caracterização micrometeorológica de uma comunidade vegetal envolve todos os processos de troca de energia e de massa, sendo fundamental o conhecimento do fluxo de CO2, visto ser este o principal substrato para a síntese dos fotoassimilados utilizados no crescimento vegetal (Brunini et al., 1999). Alguns trabalhos reportando a análise de crescimento vegetal para a cultura do milho já foram desenvolvidos (Silva, 1995; Barros, 1998; Maggio, 2006; Moura et al., 2006; Garcia et al., 2007), porém, a maioria desses estudos evoca aspetos puramente morfológicos e fisiológicos, do ponto de vista agronómico, sem, contudo, investigar uma relação mais contundente dos elementos meteorológicos restritivos e a produção potencial da cultura, verificando as respostas das taxas, oriundas da análise de crescimento, aos fatores abióticos envolvidos no microclima da canópia.
O Milho BR 106, objeto do presente estudo, através de pesquisas, vem sendo melhorado geneticamente a cada ano, proporcionando ganhos em uniformização de plantas, empalhamento de espigas e, principalmente produtividade de grãos (Noce, 2004) e é referência em pesquisas de avaliação de novos materiais (Vilarinho et al., 2008; Costa et al., 2015). Assim, com o presente trabalho teve-se por objetivo quantificar a influência das variáveis micrometeorológicas no crescimento e desenvolvimento, em dossel de milho cultivado sob sementeira direta.
Material e Métodos
O ensaio foi conduzido em condições de campo, sob pivô central, na área de experimentação da EMBRAPA Milho e Sorgo, situada no Município de Sete Lagoas, MG, geograficamente localizado em -19º 45 S e 44º 25 W, à altitude de 762 m. O clima da região é classificado, de acordo com a classificação de Köppen, como Aw, ou seja, típico de savana, com inverno seco e temperatura média do ar superior a 18oC no mês mais frio. O solo do local é Latossolo Vermelho hálico argiloso e o relevo suavemente ondulado, fase cerrado.
O genótipo de milho utilizado foi a variedade BR 106 de ciclo semiprecoce, cujas características são: rusticidade, menor custo de sementes, boa estabilidade de produção e adaptabilidade a todas as regiões brasileiras, resistência ao acamamento e ao ataque das principais pragas comuns à cultura. É um milho ao alcance de todos os produtores brasileiros, independente de seu nível tecnológico, destacando-se como um dos mais plantados no Brasil.
No limiar entre os quadrantes Nordeste e Sudeste do pivot foi instalada uma torre micrometeorológica equipada com uma Estação Meteorológica Automática (EMA), cuja finalidade foi a obtenção dos dados meteorológicos, os quais contribuíram para a caracterização do microclima no dossel do milho, ao longo do período experimental. Os dados dos elementos climáticos foram registrados a cada segundo, com valores médios calculados a cada 30 minutos, em um Datalogger. A referida estação foi composta dos sensores de temperatura e humidade relativa do ar, velocidade do vento, radiação fotossinteticamente ativa, saldo de radiação, fluxo de calor no solo, temperatura e humidade do solo, e chuva, cujas características se encontram no Quadro 1. Dados complementares de insolação e radiação solar incidente, para o mesmo período, foram disponibilizados pela EMBRAPA Milho e Sorgo por meio da Estação Climatológica Principal (ECP), bem como pela Estação Meteorológica Automática da EMBRAPA (EMAEMBRAPA), as quais ficavam localizadas, respetivamente, a 3 km e 300 m distantes do local do ensaio.
Foi instalado na torre micrometeorológica um sistema composto por um anemómetro sónico tridimensional com frequência de 21 Hz, um analisador de gás a infravermelho com microcomputador. O software utilizado foi o EddySol, de modo caminho fechado, desenvolvido na University of Edinburgh, por John Moncrieff, que calcula os fluxos de CO2, vapor de água e energia acima da canópia, em tempo real. Uma descrição completa do equipamento e software EddySol é dada por Grace (1995) e Moncrieff et al. (1997). Os sensores utilizados para estimativa desses fluxos, mostrados no Quadro 1, foram instalados na parte superior da torre, a quatro metros acima do solo, com orientação Norte (Carswell et al., 2002).
Para a análise de crescimento, foi necessário a amostragem de valores primários por meio de colheitas destrutivas, representativas do conjunto de plantas cultivadas no pivot. Assim, durante o ciclo da cultura foram realizadas avaliações para caracterização dos estádios vegetativos, reprodutivo e de formação de grãos baseado na escala fenológica para cultura do milho, adaptado de Magalhães et al. (2002) e Magalhães e Durães (2006). Numa área de controlo dentro do pivot com dimensões de 90 m x 90 m, em intervalos de oito dias, a partir da emergência, foram colhidas 20 plantas para determinar a altura da planta (AP), matéria seca aérea (MSa) e área foliar (AF). Em cada colheita a amostra de uma planta foi separada por partes (folha, colmo, pendão, espiga e grãos) para a determinação da massa seca e da área foliar. Nessas colheitas, os valores de MSa e AF foram obtidos a partir da média dos valores encontrados nas quatro linhas.
As massas secas obtidas das partes das plantas e a massa seca total foram convertidas em biomassa por metro quadrado (m2) (produtividade biológica). Para a estimativa da massa de matéria seca aérea produzida, relativa ao período de crescimento, foi empregada a função sigmoidal, utilizando-se um programa interativo para minimizar a variância residual.
A determinação da área foliar (m2) foi medida com o planímetro ótico modelo LI 3000, Licor, Lincoln, EUA.O Índice de Área Foliar (IAF) de cada amostra foi determinado como a relação entre a área foliar e a área do terreno ocupada pela amostra (m2). O modelo para estimativa do IAF foi ajustado a partir dos dados colhidos, sendo o melhor ajuste feito com o pico de Gaussian (3 parâmetros).
A eficiência das folhas na produção de novos órgãos foi calculada pela Taxa de Assimilação Líquida (TAL):
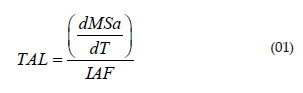
Além das taxas de crescimento foram determinadas as razões de crescimento com a finalidade de comparar o crescimento das diferentes partes da planta com outras partes bem como com a planta inteira.
Os Graus-dia Acumulados (GDA), da emergência até ao final do ciclo, foram determinados a partir da diferença entre a temperatura média diária e a temperatura basal inferior da cultura, que foi considerada como 10ºC (Cardoso et al., 1997). Salienta-se que a utilização deste modelo se dá em função de a temperatura média (Tm) no período estudado não ter superado a temperatura basal superior para o milho que é de 30ºC, conforme proposto por Villa Nova (1972). Os valores diários de GDA foram utilizados no ajuste de modelos para biomassa e índice de área foliar.
O potencial de vapor de água foi determinado por meio da equação a seguir (Reichardt, 1987):
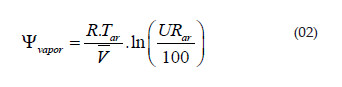
em que: Yvapor é o potencial do vapor de água no ar (MPa); R é a constante dos gases (0,0083141 LMpa.mol-1.K-1); Tar é a temperatura absoluta do ar (K); V é o volume parcial molar da água (0,018048 L.mol-1) e URar é a humidade relativa do ar (%).
Resultados e Discussão
No Quadro 2 está apresentado o resumo dos carateres morfoagronómicos do BR 106 medidos por ocasião da colheita. Verifica-se que esses tiveram valores condizentes com as médias obtidas nos programas de melhoramento (Noce, 2004) e acima de pesquisas que utilizaram o BR 106 como referência (Vilarinho et al., 2008 e Costa et al., 2015). Este resultado confirma as condições ótimas, em termos de manejo da cultura, a que o ensaio foi submetido. Desta forma a análise em relação à variação dos fatores ambientais ao longo do período do ensaio, seguiu a linha de discussão na qual a grandeza das mudanças desses fatores serviu de base para essa variedade, no que diz respeito ao potencial produtivo dentro das condições de sementeira direta com rega.
O acompanhamento do ciclo da cultura do milho compreendeu 10 estádios que foram identificados desde a emergência até ao grão fisiologicamente maduro (Quadro 3). Para nortear a discussão, objetivando correlacionar o desempenho das plantas cultivadas com os fatores ambientais, o ciclo foi dividido em períodos (Pi) importantes do ponto de vista agronómico, a saber: P1 (VE-V12: Definiu o potencial de produção e o número de fileiras nas espigas); P2 (V12- R1: Definiu o tamanho da espiga e o máximo acúmulo de matéria seca pela planta); P3 (R1-R3: Correlacionado com o período anterior, definiu o número de grãos, bem como iniciou o acúmulo de amido no endosperma dos grãos) e P4 (R3-R6: Definiu a densidade do grão).
O número de horas de brilho solar (n) e o comprimento astronómico do dia (H) em cada estádio estão apresentados na Figura 1a. A razão de insolação, obtida a partir da divisão do brilho solar pelo comprimento do dia, foi em média 0,47 para todo o ciclo, indicando um grande percentual de nebulosidade. Essa razão foi de 0,56 e 0,20, respetivamente, para P1 e P3. Mesmo com o valor de H maior, em torno de 13, o P3 teve dias proporcionalmente menos ensolarados, com brilho solar médio de 2,75. Esse dado aumenta em torno de 150% quando comparado com o P1, apesar da variação do brilho solar não apresentar tanta amplitude para ambos os períodos. Tal comportamento pode ser confirmado pelo potencial de vapor na atmosfera, Yvapor (Figura 1b). Potencial esse que é obtido pela diferença de energia entre um determinado estado e o estado padrão. Como o vapor de água em ar não saturado apresenta menor energia do que no ar saturado, o potencial do vapor de água na atmosfera é geralmente negativo (Reichardt, 1987). Assim, quanto mais negativo o valor do Yvapor, menor é a possibilidade de saturação do ar e, por consequência, menor a presença de nuvens. Nota-se que a variação extrema dessa amplitude é representada pelos valores médios do Yvapor para os períodos P1 (-68,57 atm) e P3 (-21,21 atm).
O volume de precipitação pluviométrica acumulada durante o cultivo foi de 925 mm. Mesmo assim, a fim de garantir a manutenção da humidade do solo, em condições ótimas, foi utilizada a rega suplementar, sendo aplicado o total de 86 mm, do qual 65% ocorreu antes da emissão da oitava folha.
O resumo dos valores médios da radiação incidente (global e fotossinteticamente ativa), para os períodos de P1 a P4, está apresentado no Quadro 4. Apesar do registro médio de brilho solar apresentar-se equivalente entre os períodos P1 (7,08), P2 (6,44) e P4 (6,26), a maior disponibilidade da RI e RFA ocorreu no P1, com 21,92 MJ.m-2, possivelmente por este apresentar maior número de dias com brilho solar igual ou acima de 8. No P3, devido à maior ocorrência de precipitação média do período experimental, houve maior nebulosidade promovendo redução em torno de 57% de brilho solar em relação aos demais períodos.
A radiação solar global foi em média 17,58 MJ.m-2 para todo o ciclo da cultura, com diferenças dos valores médios, entre os períodos, oscilando entre 20% e 34%. Porém, em se tratando de amplitude, foram registados valores absolutos de 4,47 MJ.m-2 (P2) e 32,26 MJ.m-2 (P3) ocorridos devido a alternância de dias com alta e baixa nebulosidade. Para a RFA, os valores representaram 45% da Radiação Solar Global (RI), e devido a alta nebulosidade os extremos variaram de 0 a 14,52 MJ.m-2, com médias que declinaram de 9,89 MJ.m-2 em P1 até 6,50 MJ.m-2 em P3. Ressalta-se, porém, que ao longo do período P3 ocorre o início da formação de grãos, concomitantemente com o começo da senescência foliar, e nessas condições o milho está a principiar a translocação das reservas acumuladas para o enchimento dos grãos, podendo diminuir a atividade fotossintética.
No Quadro 5 estão apresentados os valores médios das temperaturas médias, máximas e mínimas durante o ciclo da cultura. A amplitude térmica foi maior na fase inicial de crescimento vegetativo e na final, próximo da maturação fisiológica. A partir dessa condição e aliado ao baixo Yvapor, pode-se inferir que, em termos práticos, a cultura esteve sob atmosfera mais seca durante o período P1, mas para o P4, apesar das temperaturas extremas promoverem amplitude semelhante ao P1, a maior presença de chuva permitiu a ocorrência de um ambiente com baixo déficite de pressão de vapor (DPV) na atmosfera, para a qual tendeu a diminuição média ao longo de toda a fase reprodutiva. Nesses períodos não foram verificadas temperaturas acima de 22°C, nos horários entre 18h30min e 05h30min, as quais por serem consideradas elevadas, certamente iriam promover alto consumo energético, devido ao aumento da respiração celular, contribuindo para o menor saldo de fotoassimilados, com consequente queda no rendimento da cultura (Dourado Neto, 1999).
Constata-se também no Quadro 5 que as temperaturas mínimas foram predominantemente superiores a 10°C (temperatura-base adotada no cálculo de graus-dia) para todos os períodos. Em relação às temperaturas máximas, observa-se que nos períodos P1 e P4 elas se apresentaram levemente superiores a 30°C (temperatura basal superior do milho). Apesar dessa condição microclimática, crê-se que estas não foram limitantes, já que a influência da temperatura do ar sobre o crescimento do milho depende de outros fatores, como a duração dos seus valores extremos durante o dia. Dentro desse aspeto, mesmo com a diminuição da amplitude térmica de 15°C para 10°C entre os períodos P1 e P3, foi garantido o fornecimento de energia para o crescimento das plantas, com mais exposição da canópia ao brilho solar, por causa do aumento do fotoperíodo.
A duração do ciclo fenológico do BR 106 foi de 120 dias, apresentando o florescimento masculino em torno do 64° dia após a emergência (DAE) corroborando com a ocorrência da panícula terminal, semelhantes às médias dos ensaios nacionais (Quadro 3).
Segundo Barros (1998) a variação dos fatores ambientais, principalmente em relação à temperatura do ar, que influencia a distribuição de hidratos de carbono na planta, é estabelecida de acordo com os estádios de desenvolvimento da cultura, que é condicionado pelo somatório de graus-dia. Assim, foram feitas análises de regressão relacionando o estádio fenológico de desenvolvimento (EF) com a DAE e o GDA, para o BR 106, cujos resultados são apresentados no Quadro 6. Os valores dos coeficientes de determinação r2 foram elevados para ambas as equações, sendo ligeiramente superior na relação entre EF e GDA. Em termos de graus-dia, a taxa de desenvolvimento (coeficiente b1) foi de 0,0065. Em condições semelhantes, Silva (1995) analisou o crescimento de dois híbridos e encontrou taxas maiores entre 0,06965 e 0,07029. Essas taxas, que representam a unidade de estádio de desenvolvimento por graus-dia acumulados, foram superiores devido à condição de envelhecimento do híbrido, que tem um potencial de produção biológica superior à variedade BR 106.
As variações da produção biológica da MSa do BR 106 em função do DAE são apresentadas na Figura 2. A evolução da MSa apresentou três fases comportamentais distintas: a inicial, entre a emergência e 33 DAE, caracterizada por um acúmulo menor de biomassa; a intermediária, na qual ocorreu o acúmulo mais rápido entre 40 e 89 DAE, e a final, entre 97 e 120 DAE, na qual o acúmulo se manteve mais constante, com um pequeno decréscimo da MSa no final. Tal comportamento foi similar aos observados por Lima (1995), Silva (1995) e Barros (1998).
A contínua acumulação de massa seca total na parte aérea da planta atingiu o máximo observado de 1,753 g.m-2, aos 104 DAE, no estádio de grão pastoso. A partir desse estádio, a redução da produção de biomassa da canópia resultou do processo de redefinição de drenos preferenciais. Na realidade, essa redefinição já se tinha pronunciado no R1, quando os grãos estavam no início do enchimento, tornando-se mais evidente no R4, pois a perda das folhas senescente apresentou-se no decréscimo da área foliar ativa em relação à MSa. No estádio R6 as plantas reduziram, de forma mais contundente, as taxas de acúmulo de matéria seca aérea, devido às modificações morfofisiológicas necessárias ao suprimento da necessidade em fotoassimilados pelos espigas e grãos, além do estado avançado de senescência, em função da perda das folhas do terço médio inferior. Com os dados da MSa foi feito um ajuste, descrito por meio de uma equação gaussiana com três parâmetros, cujo coeficiente a foi de 2006,35 g.m-2, o qual representa o crescimento máximo hipotético. Para os valores ajustados, a estimativa obtida pela equação resultou num valor máximo de MSa de 1,839 g.m-2 aos 119 DAE.
A evolução do IAF do BR 106 em relação ao DAE seguiu o padrão característico de plantas anuais (Figura 3a). A curva do IAF foi ajustada ao modelo pico gaussiano, com três parâmetros que apresentou limitações na estimativa do início do ciclo da cultura até 46 DAE, mas tornou-se eficiente para o resto do ciclo, inclusive para a rápida senescência foliar. O grau de dispersão dos valores em torno da média foi de 11,58%, indicando que cerca de 88,42% da estimativa do IAF ocorrera próximo aos dados observados em campo. O comportamento do IAF mostrou-se crescente até atingir o ponto de máximo valor decrescendo em função da senescência das folhas basais, sendo esse comportamento atribuído, possivelmente, ao autossombreamento.
Borrás et al. (2003) e Kunz et al. (2007) observaram tendências semelhantes em ensaio com a cultura do milho. Os primeiros investigadores estudaram diferentes populações, enquanto os segundos analisaram o IAF para dois tipos de preparação de solo, incluindo a sementeira direta, semelhante à modalidade de preparo de solo estudado na presente pesquisa com o BR 106. Ainda, no presente estudo, apesar da população final – de 55 000 plantas por hectare – encontrar-se acima do uso comum para o BR 106, observou-se que os valores de IAF alcançaram o patamar das médias encontradas pelos ensaios nacionais com esta variedade.
O IAF é uma informação importante para o processo fotossintético, já que 80% da radiação solar global, na faixa do espectro do visível, é intercetada quando esse índice alcança os valores entre 3 e 5, que é convencionalmente denominado de canópia fechada.
Assim, baseado na relevância da informação e visando a possibilidade da estimativa, considerando as condições da área de estudo (em termos de temperatura do ar), a evolução do IAF desde a emergência até o máximo (canópia fechada), sobre os GD acumulados, foi ajustada a um modelo sigmoidal com 3 parâmetros, o qual apresentou parâmetros significativos a 1% quando analisado pelo teste F, e o coeficiente de determinação de 0,98. O coeficiente a referente à estimativa máxima do IAF foi de 3,94 ficando próximo do valor medido em campo, que foi de 3,97.
A variação da Taxa de Assimilação Líquida (TAL) em função do DAE é apresentada na Figura 3b. A TAL depende da radiação fotossinteticamente ativa, das condições internas da planta, do IAF, das condições de humidade do solo, e representa o saldo entre o material produzido pela fotossíntese e aquele perdido pela respiração. Devido ao aumento recíproco do sombreamento foliar em face do IAF, a taxa de assimilação líquida relaciona-se negativamente com ele, e consequentemente, com os demais fatores que influenciam o seu aumento. Em virtude disso, a TAL normalmente decresce durante o crescimento e desenvolvimento de um conjunto de plantas. Assim, desde o valor máximo, no início do crescimento vegetativo, até o valor mínimo, na maturação fisiológica, a queda na TAL deve-se à competição por luz, ao avanço na idade média da área foliar da planta e ao incremento das necessidades respiratórias.
Os dados médios das razões RAF (razão da área foliar, 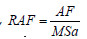 ), RMF (razão da massa foliar,
), RMF (razão da massa foliar, 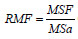 e a AFE (área foliar específica,
e a AFE (área foliar específica, 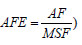 também foram calculados (Quadro 7). Os valores médios da AFE aumentaram a sua magnitude à medida que avança do P1 para o P3. Tal comportamento leva a crer que o BR 106, por meio da evolução da matéria seca das folhas ao longo de seu ciclo, registra um favorecimento da eficiência fotossintética das suas plantas, por ter folhas menos espessas, notadamente no P3 com 19,42 m2.g-1, permitindo uma diminuição das barreiras internas à entrada tanto da luz como do CO2. No P4, houve uma redução no valor da AFE, indicando que houve aumento da espessura das folhas quando a idade da canópia começou a entrar na maturação.
também foram calculados (Quadro 7). Os valores médios da AFE aumentaram a sua magnitude à medida que avança do P1 para o P3. Tal comportamento leva a crer que o BR 106, por meio da evolução da matéria seca das folhas ao longo de seu ciclo, registra um favorecimento da eficiência fotossintética das suas plantas, por ter folhas menos espessas, notadamente no P3 com 19,42 m2.g-1, permitindo uma diminuição das barreiras internas à entrada tanto da luz como do CO2. No P4, houve uma redução no valor da AFE, indicando que houve aumento da espessura das folhas quando a idade da canópia começou a entrar na maturação.
Quanto a RAF, os maiores valores foram encontrados nos P1 e P2, o que naturalmente era esperado, pois essa informação fornece o grau de produção de folhas novas, principalmente no P2, com 6,4 m2.g-1, quando ocorre o rápido crescimento vegetativo. Com o crescimento e desenvolvimento das plantas do BR 106, observa-se uma redução absoluta média de 2,84 m2.g-1 entre os períodos P2 e P4. Esse declínio de 44% justifica-se pelo aumento do autossombreamento das folhas inferiores, além da tendência de diminuição da área foliar a partir do P3 por indução à senescência dessas folhas (Benicasa, 2003). A pequena diferença da RAF de 4,8% entre P1 e P2 indica uma boa eficiência do BR 106 na captação de luz no período vegetativo dessa variedade. Os resultados da RMF mostram que no P1 o BR 106 obteve 0,50 g de matéria seca das folhas por grama de matéria seca total da parte aérea, ou seja, mais de 50% da sua biomassa seca aérea está na forma de folha, significando que a maioria dos seus fotoassimilados foram drenados para a formação de folhas.
Já que a fotossíntese fundamenta-se numa série de reações que resultam na produção de açúcares a partir do dióxido de carbono e da água, tendo como fonte de energia a radiação solar na faixa espectral do visível, e considerando ainda que é um processo físico-químico dependente da densidade de plantação e do arranjo das folhas, representado pelo IAF, foi feita uma análise confrontando o fluxo médio diário de assimilação do CO2 (Fco) com a evolução do IAF para os períodos P1 a P4 (Figura 4). Os dados do Fco foram selecionados no intervalo entre 08:00h e 18:00h, sem falhas, dentro dos períodos considerados [P1 (média de 16 dias); P2 (média de 19 dias); P3 (média de 10 dias) e P4 (média de 25 dias)]. No P1 a assimilação de CO2 e o IAF encontravam-se nos seus valores mínimos, pois neste estádio entre V4 e V8 a área foliar média era pequena, representando aproximadamente 14% da área foliar média máxima entre os períodos. Dentro dessa condição, o aparelho fotossintético da canópia do BR 106 proporcionava uma pequena assimilação do CO2 atmosférico, a saber, 0,79 µmol.m-2.s-1. Esses resultados enfatizam que o IAF expressa a disponibilidade de superfície assimiladora de CO2 e da radiação fotossinteticamente ativa da canópia de plantas (Müller e Bergamaschi, 2005).
Na transição do P1 para o período correspondente entre o pendoamento e o início do enchimento de grãos, o IAF aumentou 5 vezes, para um patamar de 3,17 m-2.m-2, e o dossel incrementou quase 8 vezes o poder assimilatório do CO2, alcançando a média diária em torno de 7,65 µmol.m-2.s-1, representando o seu valor máximo médio para todo o ciclo.
Nos períodos P3 e P4, que representam a complementação da fase reprodutiva, observa-se um pequeno incremento do IAF, em torno de 12%, em P3, para posteriormente em P4 ocorrer o declínio de aproximadamente 50%. No comportamento do CO2 observa-se uma diminuição de seu fluxo de forma mais gradual até à maturação fisiológica: com 13% (6,61 µmol.m-2.s-1) entre P2 e P3, e 12% (5,81 µmol.m-2.s-1) entre P3 e P4. Como foi afirmado existe uma relação entre a área foliar e a utilização do CO2 pelas plantas, e, desse modo, mesmo que a tendência do processo assimilatório não tenha acompanhado o aumento do IAF entre P2 e P3, nota-se que ambos os parâmetros interagem entre si, e tendem a cair com a idade das plantas. A redução do IAF é explicada pela senescência das folhas, em consequência da degradação da clorofila, tornando-as amarelas, o que aumenta a resistência estomática e, em consequência, a resistência à penetração do CO2 através do mesófilo. Essa observação torna-se mais clara na Figura 4b, onde o fluxo encontra-se confrontado com a TAL, que mede o aumento líquido da MSa da planta em função da área foliar unitária (g.m-2), com a taxa fotossintética, representada pela assimilação de carbono.
Apesar de a maior eficiência fotossintética ser apresentada no P1, tem-se concomitantemente o menor fluxo assimilado de CO2, já que a TAL reflete a dimensão do sistema assimilador que está envolvido na produção de matéria seca, ou seja, a baixa demanda respiratória, em razão da pouca biomassa e o rápido surgimento de folhas novas com alta capacidade fotossintética explicam os maiores valores médios da TAL, em torno de 32 g.m-2. No entanto, sendo um parâmetro fisiológico, a TAL diminui em função do próprio ritmo de crescimento da planta, com taxas de aparecimento e alongamento foliares, proporcionando senescência das folhas primeiramente formadas, já que aumenta a competição por luz e assimilados, que está diretamente ligada à fixação do CO2 para a produção fotossintética.
Conclusões
O crescimento do BR 106, cultivado sob o sistema de sementeira direta com rega suplementar, foi afetado pelas variações do microclima da canópia, que foram caracterizadas por grandes períodos de nebulosidade, diminuição do potencial de vapor na atmosfera e oscilações no conteúdo de energia, principalmente luz e temperatura do ar, contribuindo para que os valores máximos dos principais atributos ecofisiológicos de crescimento fossem observados no P2. Contudo a senescência das folhas do terço médio inferior, mais expressiva na transição do P2 para o P3, aumentou a RAF e a AFE, além de reduzir a RMF, dependentes do IAF e da TAL, que induziram a uma queda de 13% na assimilação do CO2, ao longo do período reprodutivo, não afetando o rendimento final de grãos do BR 106 que apresentou um valor próximo do potencial.
Agradecimentos
EMBRAPA Milho e Sorgo, pela logística da execução do ensaio experimental.
Referências Bibliográficas
Barros, A.H.C. (1998) - Análise do crescimento, do desenvolvimento e da produtividade da cultura do milho (Zea Mays L.): Experimentos e modelos. Dissertação Mestrado, UFV, Viçosa. 85 p. [ Links ]
Benicasa, M.M.P. (2003) - Análise de crescimento das plantas (Noções Básicas). Jaboticabal, FCAVUNEP. 42 p. [ Links ]
Borrás, L.; Maddonni, G.A. e Otegui, M.E. (2003) - Leaf senescence in maize hybrids: plant population, row spacing and kernel set effects. Field Crops Research, vol. 82, n. 1, p. 13-26. http://dx.doi.org/10.1016/S0378-4290(03)00002-9 [ Links ]
Brunini, O.; Machado, E.C.; Sawazaki, E.; Miranda, L.T. de. (1999) - Fluxo de gás carbônico sobre as culturas de dois híbridos de milho. Revista Brasileira de Fisiologia Vegetal, vol. 11, n. 3, p. 161-168. [ Links ]
Cardoso, M.J.; Bastos, E.A.; Andrade, JR., A.S. e Rodrigues, B.H.N. (1997) - Uso da exigência térmica para determinação do ciclo dos cultivares de milho sob irrigação. In: X Congresso Brasileiro de Agrometeorologia, 10, Piracicaba, SP, Sociedade Brasileira de Agrometeorologia, p. 23-25. [ Links ]
Carswell, F.E.; Costa, A.C.L.; Palheta, M.; Malhi, Y.; Costa, J.P.R.; Leal, L.S.M.; Costa, J.M.N.; Clemente, R.J. e Grace, J. (2002) - Seasonality in CO2 and H2O flux at an eastern Amazonian rain forest. Journal Geophysical Research, vol.107, n. D20, art. 8076. http://dx.doi.org/10.1029/2000JD000284 [ Links ]
Recebido/received: 2015.09.08
Aceite/accepted: 2016.01.15

