










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias vol.40 no.4 Lisboa set. 2017
https://doi.org/10.19084/RCA17035
ARTIGO
Development of Passiflora cincinnata Mast. submitted to different levels of nitrogen and potassium
Desenvolvimento de plantas de Passiflora cincinnata Mast. submetida a diferentes níveis de nitrogênio e potássio
Jerffson L. Santos1,*, Sylvana N. Matsumoto1, Perla N. de Oliveira2, Lucialdo O. D'Arêde1, Carmem L. L. Brito1 and Anselmo E. S. Viana1
1Departamento de Fitotecnia e Zootecnia, UESB, Campus de Vitória da Conquista, Estrada do Bem Querer, km 04, Caixa Postal 95, 45083-900, Vitória da Conquista-BA, Brasil
2 Departamento de Produção Vegetal, ESALQ, Caixa Postal 09, 13418-900, Piracicaba-SP, Brasil
(*E-mail: je.lucas@hotmail.com)
ABSTRACT
The present study aimed to evaluate the morphological characteristics of passion fruit (Passiflora cincinnata Mast.) under different levels of nitrogen and potassium, and interactions between these nutrients. The experimental design was randomized blocks, with a 4 x 4 factorial arrangement of treatments, three replications per treatment and 48 experimental plots and the experimental unit was composed of one plant. Factors were four doses of N (0, 75, 150 and 300 mg of N dm-3 of soil, applied as urea, 45 % N), and four doses of K (0, 150, 300 and 600 mg of K dm-3 of soil, applied as potassium chloride, 60 % K2O). After 60 days of transplanting the seedlings to pots, stem diameter, leaf number, SPAD (Soil Plant Analysis Development) index, leaf area, dry weight of shoot and root were evaluated. The interaction effect between levels of N and K was observed when the dry weight of shoot, stem diameter and SPAD index was assessed. The greatest growth rate of Passiflora cincinnata Mast. was obtained at doses of 180 to 300 mg of N dm-3 of soil. Potassium rates used in this study reduced the potential for accumulation of dry mass of the shoots.
Keywords: Passion fruit, ferlitization, SPAD index, dry matter.
RESUMO
O presente estudo teve como objetivo avaliar as características morfológicas de plantas de maracujá-do-mato (Passiflora cincinnata Mast.) submetidas a diferentes doses de nitrogênio e potássio, verificando as possíveis interações entre estes nutrientes. O delineamento experimental adotado foi em blocos casualizados, em esquema fatorial 4 x 4, com 16 tratamentos e três repetições, perfazendo 48 parcelas experimentais. Foram definidas quatro doses de N (0, 75, 150 e 300 mg de N dm-3 de solo) aplicadas na forma de ureia (45% N), e quatro doses de K (0, 150, 300 e 600 mg de K dm-3 de solo) utilizando o cloreto de potássio (60% K2O). Após 60 dias do transplante das mudas para os vasos, foram avaliados diâmetro do caule, número de folhas, índice SPAD, área foliar, massa seca da parte aérea e raiz. O efeito da interação entre níveis de N e K foi verificado quando foi avaliada a massa seca da parte aérea, diâmetro do caule e índice SPAD. O maior crescimento de Passiflora cincinnata Mast. foi obtido nas doses de 180 a 300 mg N dm-3 de solo. As doses de K utilizadas no presente estudo reduziram o potencial de acumulação de massa seca da parte aérea das plantas.
Palavras-chave: Maracujá-do-mato, adubação, índice SPAD, massa seca.
INTRODUCTION
The species P. cincinnata Mast., popularly known in Brazil as maracujá-do-mato, maracujá-mochila or maracujá-tubarão, is a widely distributed species in South America, and is registered from Eastern Brazil to Western Bolivia, in rupestrian field, caatinga, seasonal forest and savannah (Nunes and Queiroz, 2006). For the fact of being a perennial and resistant species to drought, it develops in a variety of semiarid region soils under unirrigated conditions (Kiill et al., 2010). Its fruits are sold in small cities in Brazil, without pesticides and with pleasant taste juicy pulp; it is particularly used in small home factories, since it is a differentiated and tasty product (Kiill et al., 2010; Santos et al., 2012).
In order to expand and diversify passiflora market, the interest of knowing the potential of other species belonging to this genus has been grown in the country (Wondracek et al., 2012). Among the wild native species in Brazil, P. cincinnata, P. setaceae, P. nitida are among the best known ones, showing the potential for economic use (Oliveira and Ruggiero, 2005), being resistant and tolerant to pests and diseases, and having good productivity with flowering during off-season periods (Junqueira et al., 2005). Several factors, such as the inadequate practice of liming and fertilizing, condition the low national average productivity of Passifloraceae. Factors such as the type, the doses, the seasons and ways to use fertilizers, combined with the ignorance of the physical and chemical characteristics of the cultivated soil, and especially the plant nutritional requirement, condition the execution of inappropriate management practices, affecting the growth and the productivity of passion fruit vine (Lima, 2005).
In order to reach the establishment of a production chain of this species, many management practices need to be defined. Despite the efforts made to build the knowledge base on P. cincinnata, up to this day there is little information on the phenotypic plasticity, genetic improvement, mineral nutrition and physiological aspects of the plant, thereby limiting the conception of management strategies for this species.
Nitrogen is an element present in several compounds which are essential for the growth and development of the plant and its deficiency causes generalized chlorosis and induction of leaves and fruits abscission (Freitas et al., 2011). Potassium, is essential for the maintenance of the hydric status of the plant and for the process of opening and closing of the stomata, which regulate the assimilation of CO2 and the production of photoassimilates (Rosolem and Steiner, 2014).
Although nitrogen (N) and potassium (K) are the most required nutrients by the cultures, the frequent plant response to fertilization is more dependent on the interaction between these elements than on the isolated nutrient (Malavolta et al., 1997). Even if N has proven a major effect on mass accumulation, many times its contribution is limited by inadequate K supply (Megda and Monteiro, 2010). Potassium is absorbed only in the form of monovalent cations and, although not incorporated into the organic compounds, it plays an important role for water transport, fruit formation, and as an activator and enzyme cofactor (Pettigrew, 2008). Apart from N, which through symbiotic relationships between plants and bacteria can be incorporated from the atmospheric air to the soil, the only source of K is the soil solution, whose availability is linked to the dynamics of the nutrient content and total K.
Thus, the objective of this study was to evaluate the development of Passiflora cincinnata Mast. plants under different doses of nitrogen and potassium, as well as the interactions between these nutrients.
MATERIALS AND METHODS
The experiment was carried out in the agricultural field at the State University of Southwest Bahia (UESB), Vitória da Conquista Campus, located at 14° 53' 23" S and 40° 48' 02" W, at 876 m of altitude. The annual average temperature varies from 19.5 to 20.5º C and the annual average air relative humidity varies from 70 to 85%.
Seedlings from light-green-colored seeds of maracujá-do-mato fruit, collected in the agricultural field of UESB and native areas in Vitória da Conquista, Bahia, were used. In order to break dormancy factor, the seeds were heated in a water bath for 5 minutes at 50°C during the planting, then they were sowed. Plastic bags, size 11x18 cm, were used, with a substrate consisting of agricultural land and aged corral manure (200L m-3). They were conducted under a netting tray with transparent plastic cover and black lateral webs, sombrite type (50% luminous restriction) and manually watered every day. After 75 days from the date of sowing, seedlings with four pairs of leaves were transplanted, keeping one plant per pot. They were irrigated daily to keep the moisture in the pot capacity.
The transplanting of seedlings to pots was made, and a moderate typical horizon A of dystrophic Yellow Latosols (Oxisols) was used as substrate (0 to 0.30 m deep). After drying the ground in open air, it was sifted through a 6 mm-network for clodding. By means of chemical analysis, mainly for soil fertility purposes, the following characteristics were observed (Table 1).

In the planting of seedlings, each pot received P (450 mg dm-3), B (0.5 mg dm-3) and Zn (5 mg dm-3). As P, B and Zn sources, single superphosphate (18% of P2O5), boric acid (17% of B) and zinc sulfate (22% of Zn) were used. The nitrogen (N) and potassium (K) doses were divided in two applying dates (50% during planting and 50% applied 30 days after planting).
The experimental design was arranged in blocks at random 4 x 4 factorial, with 16 treatments (4 doses of N and 4 doses of K) and 3 repetitions, totalizing 48 experimental units. Considering the quantitative factors, the relations between dependent variables and N and K dose were quantified by regression analysis. Urea (45% N), was used as N source (zero, 75, 150 and 300 mg of N dm-3 of soil) and potassium chloride (60% K2O) as K source (zero, 150, 300 and 600 mg of K dm-3 of soil).
The gradient of the nutrients in this study was determined based on the findings of several studies carried out on other species belonging to the genus Passiflora. This gradient was established in order to explore effects of higher nutrient doses on the growth maximizing point (Almeida et al., 2006). The experimental unit consisted of a plant placed in a pot containing 15L (14.33 dm3) of soil, arranged in a spacing of 2.0 × 2.5 m, and conducted on vertical trellis system with a wire kept at 1.80 m high from the ground. At the end of 60 days after transplanting the seedlings to pots, evaluations of the morphological and physiological characteristics such as stem diameter, number of leaves and the SPAD index (index of relative chlorophyll content), obtained by portable chlorophylometer SPAD 502, Minolta, Japan. Three readings were made at a third fully expanded leaf in a basipetal direction. In order to obtain the leaf area a leaf area integrator was used (LI-3100, LI-COR, USA). To determine the dry mass of the aerial part (DMAP) and root (MSR) of the plants they were placed in paper bags, and identified in accordance with the plot and treatment. Afterwards they were sent to the Plant Physiology Laboratory, drier in a forced air oven at 65°C for 48 h, until constant weight.
The data were submitted to homogeneity of variance tests (Bartlett and Cochran's test) and normality test (Lilliefors). The results obtained were subjected to analysis of variance using the Statistical and Genetic Analyses System software, SAEG® version 9.1. When the effects were determined to be significant, regression equations were calculated; the linear and quadratic models were tested using the F test. The models that had a probability of error of less than 5% (p < 0.05), coefficient of determination higher than 50% and biological significance were chosen.
RESULTS AND DISCUSSION
The interaction between nitrogen (N) and potassium (K) was found for the stem diameter characteristics, SPAD index and dry matter accumulation of the aerial part were determined, and also, the single effect of N to the leaf area, number of leaves and dry weight of roots (Table 2).
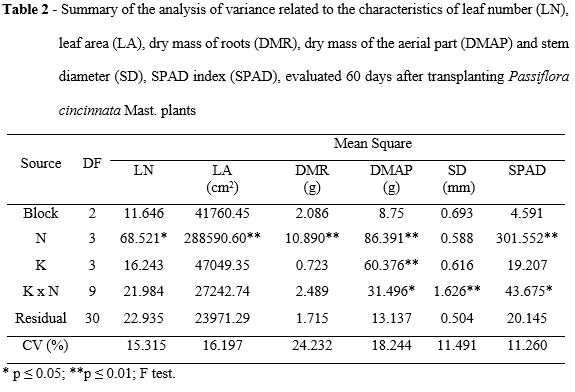
The number of leaves and the dry mass of the roots followed a quadratic behavior, in response to the nitrogen fertilization, reaching up to 32.96 leaves at the dose of 222.22 mg N dm-3, and 5.95 g of dry mass of roots at 180 mg N dm-3, respectively (Figure 1A and 1C). After the raise in the values of the number of leaves and root dry matter, there was a downward trend in those doses which were close to the maximum limit of N. The increase in the N availability in the substrate, usually results in positive effects on the carbon assimilation rate, because this nutrient is one of the main components of the photosynthetic system (Correia et al., 2005). However, positive effects of N up to the limit of its maximization for number of leaves and biomass accumulation occur within a great range of concentrations of this nutrient.
Opposite results were verified by Mendonça et al. (2007) in yellow passion fruit, showing linear behavior for the number of leaves and dry mass of the root when exposed to much higher nitrogen doses to the already evaluated results in this study. Plant biomass accumulation was restricted, either at lower or higher N doses than the defined nutrient gradient. This effect was characterized in studies done by Menegazzo et al. (2011) on papaya seedlings, and the quadratic models for the relations between nitrogen fertilization and root dry mass accumulation, aerial part and total biomass were defined. Souza et al. (2007) determined the quadratic model for the relation between N and mass accumulation of the aerial part of sweet passion fruit tree, however, the reducing effect of N was express at doses higher than 1600 mg dm-3 of substrate; in a present study, the when N was up to 300 mg dm-3 of substrate. This difference might be associated to a higher susceptibility of Passiflora cincinnata to N. According to Bredemeier and Mundstock (2000), the amount of N absorbed varies along the plant development cycle, due to the amount of roots and the root absorption rate per weight unit of the root. Almeida et al. (2006) observed the positive effect on the dry mass accumulation of the root with nitrogen fertilization for yellow passion fruit.
For the ratio between the total leaf-area of the plant (LA) and the nitrogen dose, a linear model was adjusted (Figure 1B). The largest LA (1140.57 cm-2) was measured at a maximum dose of 300 mg N dm-3. It should be noticed that, although there was a reduction in the number of leaves in the N levels above 222.22 mg N dm-3, the increasing co-relation between the leaf area and the levels of N remained. The larger leaf-area of each individual leaf in plants at higher N availability was the key factor for this behavior.
Santos et al. (2011) in a trial about different sources of N in the initial growth of yellow passion fruit tree, observed that the highest average values per leaf area, occurred in the treatment using urea, when compared with other sources of nitrogen. According to Kerpel et al. (2006), considering the P. suberosa, the additions of N until 300 mg dm-3 were associated with increases in leaf area, similarly to the one observed in the present study.
The use of indirect measurements to determine the nutritional status of plants is registered in several studies (Guimarães et al., 1999; Pôrto et al., 2011). Follett et al. (1992) studied the relation between productivity and nitrogen concentration in wheat leaves, using the SPAD chlorophyll measurer, and they observed that it is possible to calibrate the chlorophyll measurer to evaluate or predict the grain production, nitrogen concentration in leaves and NH4+ + NO3- availability in the soil.
When the K doses were fixed, because of the N doses, the maximum SPAD value reached 47.82, 45.53, 48.07 and 48.04 respectively (Figure 2A). Only a small variation of 2.45 among the values mentioned above was observed, indicating an upper limit of SPAD index, similar to all K doses which were analyzed as a function of the N gradient.
For plants that did not receive potassium fertilization, the linear model, characterized by raisings on the SPAD index values with increasing N, was delineated. The relation between SPAD reading and N content is attributed mainly to the fact that from 50% to 70% of the total N of the leaves is directed to synthesize compounds which are associated with chloroplasts and chlorophyll content of the leaves (Prado and Vale, 2008).
However, as the potassium fertilization doses were increased, the maximum SPAD value could be determined by N successively smaller doses, as it can be seen through the defined second order models (Figure 2A). According to Medeiros et al. (2009) the excess of mineral fertilization might increase the soil salinity resulting in reduced availability of water for the plants, changing their physiological and metabolic processes. Ashraf and Harris (2013) indicated that the reduction of the photosynthetic pigment contents in plants under salinity and water restriction are affected by synthesis restriction factors and acceleration in degradation processes.
Another indirect effect of salinity that is commonly reported is the reduction in the photosynthetic capacity, which promotes the degradation of chlorophyll pigments and the degradation of the thylakoids membranes (Ashraf and Harris, 2013). Shabala and Munns (2012) verified that in salinity conditions, plants reduce the opening of the stomata. The stomatal closing, induced by the salinity, causes an imbalance between the photochemical stages and the carbon reduction (Silva et al., 2010), increasing the ratio NADPH/NADP+ in the stroma due to a smaller activation of the Calvin Cycle (Silveira et al., 2010). In conditions of excessive reducing power, the thylakoids can be structurally unsettled due to the peroxidation processes in the lipid portion of the membranes, resulting in degradation of the chlorophyll.
The SPAD index reductions occurred can be related to the chlorosis in leaves, because of the toxic effect caused by the NH4+ excess (Britto and Kronzucker, 2002). According to Silva et al. (2011) studying the SPAD index in different times and locations in potato leaflets under nitrogen fertilization, a quadratic increase on the readings of the SPAD index was observed, determined in the proximal region of the leaf. When analyzing the increasing linear model of the plants that received 75 mg N dm-3 due to the increase of potassium fertilization, the positive interaction between K and N is confirmed ( Figure 2B). For the lowest N dose, the increasing of the SPAD index, due to the K gradient was related to the favorable nutritional balance for the chlorophyll formation. According to Porto et al. (2013), the increase in the SPAD readings in rocket leaves depends on the interaction between N and K. The same authors found maximum values of chlorophyll content in the 156.45 mg dm-3, K dose, and the value of 55.82 as SPAD reading. Marques et al. (2011) noticed that high doses from different sources of potassium (K2SO4 and KCl) led to reduced dry matter production of eggplant roots and shoots to 120 DAT (days after transplanting). This mass production fall was related to raisings in the electrical conductivity rates of the soil solution.
Mascarenhas et al. (2000) said that the application of high doses of potassium fertilizer can cause nutritional imbalance in plants. This fact demonstrated the antagonistic effect of K, regarding the Ca and Mg absorption, as they are all cationic nutrients that fight for the same sites of absorption. Chlorophylls are molecules formed by complexes of porphyrin derivatives, whose central atom is the Mg that is linked to 4 atoms of N (Streit et al., 2005).
The opposite behavior to the highest dose of N (300 mg dm-3) due to the increase of the K gradient, defined by the decreasing linear model can be related to a lesser absorption and availability of magnesium (Mg) in plants with increased K rates, resulting in a nutritional imbalance of the plant, because of the highest amount of N and the lowest availability of Mg, thereby interfering in the chlorophyll formation. This reduction in Mg due to the highest doses of K supplied to yellow passion fruit was noticed before by Natale et al. (2006) and Prado et al. (2004).
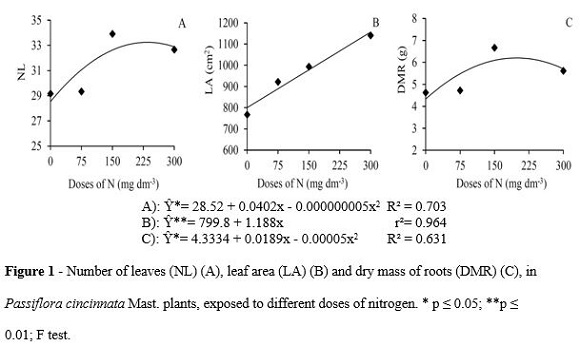
The interaction between N and K was observed when the stem diameter was quantified, making possible to establish the square root model for the doses of the evaluated elements ( Figure 3). For 0 and 300 mg of K dm-3 doses, the N gradient was characterized by diameter raises, showing a variation between 5.5 to 6.93 mm, thus defining higher values for the plants under a 0 mg K dm-3 dose (Figure 3A).
The model definition for plants under 0 mg of N dm-3 (Figure 3B) was only possible when the relation between stem diameter and K doses was analyzed. According to Kanai et al. (2011), there was a close and positive relation between the expansion of the stem diameter, aquaporin activity and conveyors of K channels in the roots of tomato trees. For the present study, such a behavior was observed in plants under low N availability until the supplying limit of 139.04 mg of K dm-3. Higher doses of K resulted in reduced stem diameter, indicating greater intensity effect of N gradient for such a feature. Almeida et al. (2006) observed quadratic behavior because of the fertilizers which were applied to the stem diameter of yellow passion fruit plants, reaching the maximum development in approximate doses of 342 mg of N and 207 mg of K dm-3.
Due to the high value of the upper limit of the potassium fertilization gradient, the possibility of induction of the water availability restriction by K, through the soil water potential reduction should also be considered. According to Silva et al. (2001), the application of high doses of fertilizers may result in a salinity raise, as for example, potassium chloride. The KCl affects the growth and distribution of the roots as well as the absorption of water and nutrients, decreasing the osmotic potential near the rhizosphere, making difficult the absorption of ions by roots.
By analyzing the interaction between N and K, for 0 and 300 mg dm-3, doses of K, dry mass accumulation of the shoot due to the increasing doses of N supplied to the plant was noticed (Figure 4A). The high magnitude of the effect of N gradient for plants without potassium fertilization was expressed by the biggest angle coefficient of the defined model for the dry mass of the aerial parts of these plants when compared to the dose of 300 mg K dm-3.
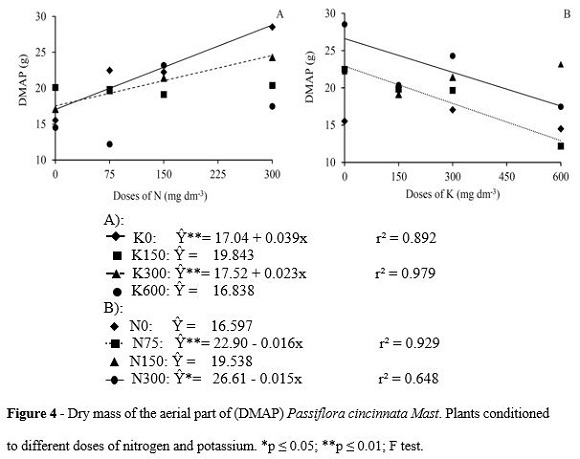
The greatest expression of N gradient to the DMAP accumulation was observed for plants that did not receive potassium fertilization in relation to those under 300 mg K dm-3 doses. The addition from 0 to 300 mg N dm-3 resulted in a differential of 11.7 g of the shoot dry mass for plants without potassium fertilization and 6.9 g for plants under 300 mg K dm-3. Although the mass accumulation is directly related to the availability of N for the plant, in the presence of K, the effect of nitrogen fertilization has become less pronounced.
This effect was related to the decrease of the plant development when using high doses of KCl, and it may increase the chloride content in the plant and cause toxicity (Silva et al., 2001; Prado et al. 2004). Another important fact to be considered was the greater availability of photoassimilates directed to the aerial part of the plant, resulting from the growth restriction of the root system at the upper limit of N doses of this study.
Almeida et al. (2006) evaluated the production of dry matter of shoots in yellow passion fruit seedlings. Considering the N and K fertilization, there was a maximum response of 16 g pot-1 production in a dose of approximately 362 mg N dm-3 and 14 g pot-1 for 300 mg K dm-3.
For doses of 75 and 300 mg N dm-3 of soil a different behavior to the relation between dry mass weight of shoot and doses of K was verified (Figure 4B). For any K gradient, the highest mass values to the highest dose of N were maintained. However, the parity between the patterns of decreases in K values remained, and can be seen by the similarity between the angular coefficients of the defined models for 75 and 300 mg N dm-3. The reductions which were seen in this study were related to the high K levels supplied, associated with an actual availability in the soil, thus interfering negatively with the accumulation of dry mass of the aerial part of the plant. According to Marschner (1995), over-fertilization of KCl can increase the salinity of the soil, making difficult the growth of the plant. This effect was shown by Nascimento et al. (2012) in cowpea (feijão-de-corda), a reduction in the production of dry biomass of the aerial part, in response to the salinity increase.
According to Prado et al. (2004) from doses of 225 and 300 mg of K dm-3 in yellow passion fruit tree, there was a reduction in the growth and production of dry matter of the plants, which has been linked to the depressive effects caused by the chloride and its antagonistic effect concerning the nitrate. However, the application of high doses of K associated with high levels of nitrogen fertilization increased both the grain production and the dry matter accumulation of corn plants.
For the present study, contrasting Viana and Kiehl (2010) even for the highest dose of N, potassium gradient was related to systematic dry matter decreases of the aerial part of the passion fruit plants. Supporting this difference, as described above, the most remarkable effect of N for dry matter accumulation of the aerial part was observed for plants that did not receive potassium fertilization.
CONCLUSION
Nitrogen was the main factor that influenced the Passiflora cincinnata Mast. plants morphology, and the largest growth was obtained with 180 to 300 mg N dm-3.
The interaction between nitrogen and potassium doses resulted in the definition of different models associated to morphological traits.
The potassium gradient induced salinity effects as noticed in this study, thus reducing the accumulation of dry mass in the aerial part of passion fruit (P. cincinata).
REFERENCES
Almeida, E.V.; Natale, W.; Prado R. de M. & Barbosa, J.C. (2006) - Adubação nitrogenada e potássica no desenvolvimento de mudas de maracujazeiro. Ciência Rural, vol. 36, n. 4, p. 1138-1142. http://dx.doi.org/10.1590/S0103-84782006000400015 [ Links ]
Ashraf, M. & Harris, P.J.C. (2013) - Photosynthesis under stressful environments: An overview. Photosynthetica, vol. 51, n. 2, p. 163-190. http://dx.doi.org/10.1007/s11099-013-0021-6 [ Links ]
Bredemeier, C. & Mundstock, C.M. (2000) - Regulação da absorção e assimilação do nitrogênio nas plantas. Ciência Rural, vol. 30, n. 2, p. 365-372. http://dx.doi.org/10.1590/S0103-84782000000200029 [ Links ]
Britto, D.T. & Kronzucker, H.J. (2002) - NH4+ toxicity in higher plants: a critical review. Journal of Plant Physiology, vol. 159, n. 6, p. 567-584. http://dx.doi.org/10.1078/0176-1617-0774 [ Links ]
Correia, C.M.; Pereira, J.M.M.; Coutinho, J.F.; Björn, L.O. & Pereira, J.M.G.T. (2005) - Ultraviolet-B radiation and nitrogen affect the photosynthesis of maize: a Mediterranean field study. European Journal of Agronomy, vol. 22, n. 3, p. 337-347. http://dx.doi.org/10.1016/j.eja.2004.05.002 [ Links ]
Follet, R.H.; Follet, R.F. & Halvarson, A.D. (1992) - Use a chlorophyll meter to evaluate the nitrogen status of dryland winter wheat. Communications in Soil Science and Plant Analysis. vol. 23, n. 7/8, p. 517-697. http://dx.doi.org/10.1080/00103629209368619 [ Links ]
Freitas, M.S.M.; Monnerat, P.H.; Carvalho, A.J.C. & Vasconcellos, M.A.S. (2011) - Sintomas visuais de deficiência de macronutrientes e boro em maracujazeiro-doce. Revista Brasileira de Fruticultura, vol. 33, n. 4, p. 1329-1341. http://dx.doi.org/10.1590/S0100-29452011000400034 [ Links ]
Guimarães, T.G.; Fontes, P.C.R.; Pereira, P.R.G.; Alvarez, V.V.H. & Monnerat, P.H. (1999) - Teores de clorofila determinados por medidor portátil e sua relação com formas de nitrogênio em folhas de tomateiro cultivados em dois tipos de solo. Bragantia, vol. 58, n. 1, p. 209-216. http://dx.doi.org/10.1590/S0006-87051999000100020 [ Links ]
Junqueira, N.T.V.; Braga, M.F.; Faleiro, F.G.; Peixoto, J.R. & Bernacci, L.C. (2005) - Potencial de espécies silvestres de maracujazeiro como fonte de resistência a doenças. In: Faleiro, F.G.; Junqueira, N.T.V. & Braga, M.F. (Eds.) - Maracujá: germoplasma e melhoramento genético, Planaltina, DF: Embrapa Cerrados. p. 81-107. [ Links ]
Kanai, S.; Moghaieb, R.E.; El-Shemy, H.A.; Painigrahi, R.; To, J.; Nguyen N.T.; Saneoka, H. & Fujita, K. (2011) - Potassium deficiency affects water status and photosynthetic rate of the vegetative sink in green house tomato prior to its effects on source activity. Plant Science, vol. 180, n. 2, p. 369-374. http://dx.doi.org/10.1016/j.plantsci.2010.10.011 [ Links ]
Kerpel, S.M., Soprano, E. & Moreira, G.R.P. (2006) - Effect of nitrogen on Passiflora suberosa L. (Passifloraceae) and consequences for larval performance and oviposition in Heliconius eratophyllis (Fabricius) (Lepidoptera: Nymphalidae). Neotropical Entomology, vol. 35, n. 2, p. 192-200. http://dx.doi.org/10.1590/S1519-566X2006000200006 [ Links ]
Kiill, L.H.P.; Siqueira, K.M.M.; Araújo, F.P.; Trigo, S.P.M.; Feitoza, E.A. & Lemos, I.B. (2010) - Biologia reprodutiva de Passiflora cincinnata Mast. (Passifloraceae) na região de Petrolina (Pernambuco, Brazil). Oecologia Australis, vol. 14, n. 1, p. 115-127. http://dx.doi.org/10.4257/oeco.2010.1401.05 [ Links ]
Lima, A. de A. (2005) - Aspectos fitotécnicos: desafios da pesquisa. In: Faleiro, F.G.; Junqueira, N.T.V. & Braga, M.F. (Eds.) - Maracujá: germoplasma e melhoramento genético, Planaltina, DF: Embrapa Cerrados. p. 643-677 [ Links ]
Malavolta, E.; Vitti, G.C. and Oliveira, S.A. (1997) - Avaliação do estado nutricional das plantas: princípios e aplicações. Piracicaba: Associação Brasileira da Potassa e do Fosfato. 319 p. [ Links ]
Marques, D.J.; Broetto, F.; Silva, E.C. & Carvalho J.G. (2011) - Dinâmica de cátions na raiz e folhas de berinjela cultivada sobre doses crescentes de potássio oriundas de duas fontes. Idesia, vol. 29, n. 2, p. 69-77. http://dx.doi.org/10.4067/S0718-34292011000200009 [ Links ]
Marschner, H. (1995) - Mineral nutrition of higher plants. London: Academic Press. 889 p. [ Links ]
Mascarenhas, H.A.A.; Tanaka, R.T.; Carmello, Q.A. de C.; Gallo, P.B. & Ambrosano, G.M.B. (2000) - Calcário e potássio para a cultura da soja. Scientia Agricola, vol. 57, n. 3, p. 445-449. http://dx.doi.org/10.1590/S0103-90162000000300012 [ Links ]
Medeiros, P.R.F. de; Duartes, S.N. & Dias, C.T.S. (2009) - Tolerância da cultura do pepino à salinidade em ambiente protegido. Revista Brasileira de Engenharia Agrícola e Ambiental, vol. 13, n. 4, p. 406-410. http://dx.doi.org/10.1590/S1415-43662009000400006 [ Links ]
Megda, M.M. & Monteiro, F.A. (2010) - Nitrogen and potassium supply and the morphogenic and productive characteristics of marandu palisadegrass. Revista Brasileira de Zootecnia, vol. 39, n. 8, p. 1666-1675. http://dx.doi.org/10.1590/S1516-35982010000800007 [ Links ]
Mendonça, V.; Ferreira, E.A.; Paula, Y.C.M.; Batista, T.M.V. & Ramos, J.D. (2007) - Crescimento de mudas de maracujazeiro-amarelo influenciado por doses de nitrogênio e de superfosfato simples. Revista Caatinga, vol. 20, n. 4, p. 137-143. [ Links ]
Menegazzo, M.L., Oliveira, A.C. de & Silva, E.A. da (2011) - Adubação nitrogenada na produção de mudas de mamoeiro. Revista Agrarian, vol.4, n. 13, p. 189-196. [ Links ]
Nascimento, R.; Nascimento, D.A.M.; Silva, D.A. & Alves, A.G. (2012) - Índice spad e partição de biomassa em plantas de feijão-caupi submetidas ao estresse salino. Revista Educação Agrícola Superior, vol. 27, n. 2, p. 128-132. http://dx.doi.org/10.12722/0101-756X.v27n02a09 [ Links ]
Natale, W.; Prado, R. de M.; Almeida, E.V. & Barbosa, J.C. (2006) - Adubação nitrogenada e potássica no estado nutricional de mudas de maracujazeiro-amarelo. Acta Scientiarum. Agronomy. vol. 28, n. 2, p. 187-192. http://dx.doi.org/10.4025/actasciagron.v28i2.1036 [ Links ]
Nunes, T. S. & Queiroz, L.P. (2006) - Flora da Bahia: Passifloraceae. Sitientibus série Ciências Biológicas, vol. 6, n. 3, p. 194-226. [ Links ]
Oliveira, J.C. de & Ruggiero, C. (2005) - Espécies de maracujá com potencial agronômico. In: Faleiro, F.G.; Junqueira, N.T.V. & Braga, M.F. (Eds.) - Maracujá: germoplasma e melhoramento genético, Planaltina, DF: Embrapa Cerrados. p. 143-158. [ Links ]
Pettigrew, W.T. (2008) - Potassium influences on yield and quality production for maize, wheat, soybean and cotton. Physiologia Plantarum, vol. 133, n. 4, p. 670-681. http://dx.doi.org/10.1111/j.1399-3054.2008.01073.x [ Links ]
Pôrto, M.L.; Puiatti, M.; Fontes, P.C.R.; Cecon, P.R.; Alves, J.C. & Arruda, J.A. (2011) - Índice SPAD para o diagnóstico do estado de nitrogênio na cultura da abobrinha. Horticultura Brasileira, vol. 29, n. 3, p. 311-315. http://dx.doi.org/10.1590/S0102-05362011000300009 [ Links ]
Porto, R.A.; Bonfim-Silva, E.M.; Souza, D.S.M.; Cordova, N.R.M.; Polyzel, A.C. & Silva, T.J.A. (2013) - Adubação potássica em plantas de rúcula: produção e eficiência no uso da água. Revista Agro@mbiente, vol. 7, n. 1, p. 28-35. http://dx.doi.org/10.18227/1982-8470ragro.v7i1.760 [ Links ]
Prado, R.M.; Braghirolli, L.F.; Natale, W.; Corrêa, M.C.M. & Almeida, E.V. (2004) - Aplicação de potássio no estado nutricional e na produção de matéria seca de mudas de maracujazeiro-amarelo. Revista Brasileira de Fruticultura, vol. 26, n. 2, p. 295-299. http://dx.doi.org/10.1590/S0100-29452004000200028 [ Links ]
Prado, R. de M. & Vale, D.W. do (2008) - Nitrogênio, fósforo e potássio na leitura spad em porta-enxerto de limoeiro-cravo. Pesquisa Agropecuária Tropical, vol. 38, n. 4, p. 227-232. [ Links ]
Rosolem, C.A. and Steiner, F. (2014) - Adubação potássica para o crambe. Bioscience Journal, vol. 30, n. 3, p. 140-146. [ Links ]
Santos, T.C.; Júnior, J.E.N. & Prata, A.P.N. (2012) - Frutos da Caatinga de Sergipe utilizados na alimentação humana. Scientia Plena, vol. 8, n. 4, p. 1-7. [ Links ]
Santos, P.C. dos; Lopes, L.C.; Freitas, S. de J.; Sousa, L.B. de & Carvalho, A.J.C. de (2011) - Crescimento inicial e teor nutricional do maracujazeiro amarelo submetido à adubação com diferentes fontes nitrogenadas. Revista Brasileira de Fruticultura, vol. 33, n. Especial, p. 722-728. http://dx.doi.org/10.1590/S0100-29452011000500101 [ Links ]
Shabala, S. & Munns, R. (2012) - Salinity stress: physiological constraints and adaptive mechanisms. In: Shabala, S. (Ed.) - Plant Stress Physiology. Wallingford, UK: CAB International. p. 59-93. [ Links ]
Silva, E.N.; Ribeiro, R.V.; Ferreira-Silva, S.L.; Viégas, R.A. & Silveira, J.A.G. (2010) - Comparative effects of salinity and water stress on photosynthesis, water relations and growth of Jatropha curcas plants. Journal of Arid Environments, vol. 74, n. 10, p. 1130-1137. http://dx.doi.org/10.1016/j.jaridenv.2010.05.036 [ Links ]
Silva, M.A.G.; Boaretto, A.E.; Fernandes, H.G. & Scivittaro, W.B. (2001) - Efeito do cloreto de potássio na salinidade de um solo cultivado com pimentão, Capsicum annuum L., em ambiente protegido. Acta Scientiarum. Agronomy, vol. 23, n. 5, p. 1085-1089. http://dx.doi.org/10.4025/actasciagron.v23i0.2563 [ Links ]
Silva, M.C.C.; Coelho, F.S.; Braun, H. & Fontes, P.C.R. (2011) - Índice SPAD em função de diferentes horários e posições no folíolo da batata sob fertilização nitrogenada. Revista de Ciência Agronômica, vol. 42, n. 4, p. 971-977. http://dx.doi.org/10.1590/S1806-66902011000400020 [ Links ]
Silveira, J.A.G.; Silva, S.L.F.; Silva, E.N. & Viégas, R.A. (2010) - Mecanismos biomoleculares envolvidos com a resistência ao estresse salino em plantas. In: Gheyi, H.R.; Dias, N.S. & Lacerda, C.F. (Eds.) - Manejo da salinidade na agricultura irrigada: estudos básicos e aplicados. Fortaleza: INCTSal, p. 161-180. [ Links ]
Souza, H.A.; Mendonça, V.; Abreu, N.A.A.; Teixeira, G.A.; Gurgel, R.L.S. & Ramos, J.D. (2007) - Adubação nitrogenada e substratos na produção de mudas de maracujazeiro doce. Ciência e Agrotecnologia, vol. 31, n. 3, p. 599-604. http://dx.doi.org/10.1590/S1413-70542007000300001 [ Links ]
Streit, N.M.; Canterle, L.P.; Canto, M.W. do & Hecktheuer, L.H.H. (2005) - As Clorofilas. Ciência Rural, vol. 35, n. 3, p. 748-755. http://dx.doi.org/10.1590/S0103-84782005000300043 [ Links ]
Viana, E.M. & Kiehl, J.C. (2010) - Doses de nitrogênio e potássio no crescimento do trigo. Bragantia, vol. 69, n. 4, p. 975-982. http://dx.doi.org/10.1590/S0006-87052010000400024 [ Links ]
Wondracek, D.C.; Vieira, R.F.; Silva, D.B.; Agostini-Costa, T.S.; Sano, S.M. & Faleiro, F.G. (2012) - Influência da saponificação na determinação de carotenóides em maracujás do cerrado. Química Nova, vol. 35, n. 1, p. 180-184. http://dx.doi.org/10.1590/S0100-40422012000100031 [ Links ]
Received/recebido: 2017.02.19
Accepted/aceite: 2017.08.10

