










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias vol.41 no.1 Lisboa mar. 2018
https://doi.org/10.19084/RCA17152
ARTIGO
Agronomic characteristics of sunflower genotypes according to plant population
Características agronômicas de genótipos de girassol segundo a população de plantas
Diogo Stasiak1, Flávio Carlos Dalchiavon1*, Marcos Birck1, Rosivaldo Hiolanda1, Andréia Fernanda Silva Iocca1, Admar Junior Coletti2 and Claudio Guilherme Portela Carvalho3
1Instituto Federal de Educação, Ciência e Tecnologia de Mato Grosso, Departamento de Agronomia, Rodovia MT 235, km 12, Zona Rural, Caixa Postal 100, CEP 78360-000, Campo Novo do Parecis, MT, Brasil
2Universidade Federal de Mato Grosso, Sinop,MT, Brasil
3Embrapa Soja, Caixa Postal 231, CEP 86001-970, Londrina, PR, Brasil
*(E-mail: flavio.dalchiavon@cnp.ifmt.edu.br)
ABSTRACT
The agronomic characteristics of sunflower genotypes were evaluated under different plant populations. The experiment was performed at the IFMT experimental field, campus Campo Novo do Parecis, Brazil, between February and June 2015. The experimental design comprised randomized blocks, in a factorial 5 x 5 arrangement, with five sunflower genotypes (AGUARÁ 04, GNZ NEON, HÉLIO 251, SYN 045 and SYN 3950HO) and five plant populations (30,000; 37,500; 45,000; 52,500; 60,000 plants ha-1), with 3 replications. Plant height, stem diameter, capitulum height, capitulum size, achene mass per capitulum, mass of one thousand achenes and achene productivity were assessed. Increase in plant and capitulum height, reduction of stem diameter, capitulum size, achene mass per capitulum and mass of one thousand achenes occurred with increase of plant population. Productivity is correlated with the mass of achene per capitulum and with mass of one thousand achenes. The population of 47,150 plants ha-1 provided the highest achene productivity.
Keywords: performance of cultivars, Helianthus annuus L., seeding density, agronomic performance, productivity.
RESUMO
Objetivou-se avaliar características agronômicas de genótipos de girassol sob diferentes populações de plantas. O experimento foi realizado no campo experimental do IFMT, Campus Campo Novo do Parecis - MT, entre os meses de fevereiro e junho de 2015. O delineamento experimental foi o de blocos casualizados, em esquema fatorial 5 x 5, sendo cinco genótipos de girassol (AGUARÁ 04, GNZ NEON, HÉLIO 251, SYN 045 e SYN 3950HO) e cinco populações de plantas (30.000, 37.500, 45.000, 52.500 e 60.000 plantas ha-1), com 3 repetições. Avaliaram-se as características altura de planta, diâmetro da haste, altura do capítulo, tamanho do capítulo, massa de aquênios por capítulo, massa de mil aquênios e produtividade de aquênios. Há aumento da altura de planta e da altura de capítulo, redução do diâmetro da haste, do tamanho do capítulo, da massa de aquênios por capítulo e da massa de mil aquênios com o aumento da população de plantas. A produtividade está correlacionada com a massa de aquênios por capítulo e com a massa de mil aquênios. A população de 47.150 plantas ha-1 proporciona a maior produtividade de aquênios.
Palavras-chave: desempenho de cultivares, Helianthus annuus, densidade de semeadura, desempenho agronômico, produtividade.
INTRODUCTION
The production of sunflower (Helianthus annuus L.) in the state of Mato Grosso, Brazil, is 77.4% of total Brazilian production The region has the best soil-climate conditions for the development of the sunflower crop (Pérez et al., 1989). Cultivated area in Mato Grosso for the 2013-14 harvest was 126,200 hectares, with a production of 203,300 tons, whereas the cultivated area for the 2014-15 harvest was 86,400 hectares, with a production of 116,500 tons, or rather, a 31.5% decrease in cultivated area and 42.7% and 16.3% respectively for production and productivity when compared to the previous harvest. Low technology and cultivation treatments different from traditional ones may have been the main reduction cause (CONAB, 2015).
Since vegetative and reproductive characteristics of sunflower crops vary according to genotype, it is important to select, prior to implantation, the genotype with the due agronomic features. In Papanduva SC Brazil, Vogt et al. (2012) assessed 17 sunflower genotypes and reported a difference of 2,081.0 kg ha-1 between the genotype with the highest (Dow M734; 2,632.0 kg ha-1) and lowest (Hélio 358; 551.0 kg ha-1) productivity rate. In an assay in Campo Novo do Parecis MT Brazil, there was a difference of 1,069.0 kg ha-1 between genotypes with the highest (SYN 045; 2,674.0 kg ha-1) and lowest (BRS 323; 1,605.0 kg ha-1) productivity rates during the 2013 interim harvest (Carvalho et al., 2014).
In studies undertaken in Eldorado do Sul RS Brazil, with four plant populations (30,000; 50,000; 70,000 and 90,000 plants ha-1) in the 1989-90 and 1990-91 harvests, Rizzardi and Silva (1993) reported that sunflower genotypes in different plant populations enhanced productivity. There was no difference in productivity in the short cycle and low size genotype for the 1989-1990 harvest (Contisol 711) in different plant populations, although long cycle and medium and high size genotypes (DK 180 and GR 10, respectively) had the greatest productivity rates with a population of 30,000 plants ha-1. In the 1990-91 harvest, short cycle and low size genotype (GR 16) revealed highest productivity in the population of 56,279 plants ha-1. In their studies on different sunflower plant populations, Castro et al. (2011) reported that productivities around 2,500.0 kg ha-1 were obtained with populations between 31,000 and 45,000 plants ha-1. On the other hand, Castro et al. (1997) insisted that seeding density, which causes highest productivity rates, lay between 40,000 and 45,000 plants ha-1. No population size is established since it varies according to the region.
Since scanty information is available on sunflower cultivation and its importance in Brazilian economy, studies are highly relevant to increase productivity and provide agronomic information to producers. Cultivation practices will be underscored and future research work may be monitored. Current assay evaluates the agronomic characteristics of sunflower genotypes under different plant populations.
MATERIALS AND METHODS
Current assay was conducted in the experimental field of the Federal Institute of Education, Science and Technology of Mato Grosso, campus Campo Novo do Parecis, during the second harvest of the 2014/2015 agricultural year, at 13º4037 S and 57º4730 W, altitude 574 m.
Soil is Dystrophic Red Latosol (Dalchiavon et al., 2015). Prior to the preparation of the area with soil disking, soil samples at 0 – 0.2 m depth was removed to analyze their chemical properties. Mean data were pH (H2O) = 5.7; P = 6.1 mg dm-3; K = 0.24 cmolc dm-3; Ca = 2.5 cmolc dm-3; Mg = 1.0 cmolc dm-3; Al = 0; H = 4.6 cmolc dm-3; B = 0.36 mg dm-3; organic matter = 35.8 g dm-3 and V% = 44.8%.
Following Köppen´s classification, the local climate is Aw, tropical climate with a well-defined dry season between May and September. Figure 1 shows the meteorological data on air temperature and accumulated rainfall on the experimental area during the experiment
Seed was sowed on the 2nd February 2015, by hand, at a depth of 0.04 m, with three seeds per hole; thinning occurred after 16 days of seeding (DAS), with one plant per hole. Fertilization occurred in the hole with 40 kg ha-1 N, 83 kg ha-1 P2O5; 32 kg ha-1 K2O and 0.4 kg ha-1 B, source MAP (8% N, 51% P2O5), formulated N-P2O5-K2O (16-16-16) and ESPHERIC (2% N, 4.82% S, 10% B, 5% Zn), plus one application of 1 L ha-1 Vitalsolo Boro (10% de B; D=1.3 g L-1). Cover fertilization occurred on the 27th DAS with 19 kg ha-1 N, 7 kg ha-1 S, 1.52 kg ha-1 B, source N-P2O5-K2O (33-00-00 + 11% S) and ESPHERIC, plus application on leaf with 1 L ha-1 Vitalboro Polyol (10% B; D=1.3 g L-1).
Experimental design comprised randomized blocks, with a 5 x 5 factorial scheme of five genotypes (AGUARÁ 04, GNZ NEON, HÉLIO 251, SYN 045, SYN 3950HO) and five plant populations (30,000; 37,500; 45,000; 52,500; 60,000 plants ha-1), with 3 replications, totaling 75 experimental plots. Each plot had an area of 15.75 m2, with 7 sowing rows, distant 0.45 m apart, and 5.0 m long. The plot´s usable area and the third and fourth row were taken into account, with the removal of one plant at each extremity.
When full florescence (stage R5.5) occurred in five plants of the plot´s usable area, plant height (PH; cm) was evaluated by measuring tape, from the ground level till the insertion of the capitulum; similarly, stem diameter (SD; mm), measured by digital caliper at 0.05 m from ground level. Capitulum height (CH; cm) was determined by evaluating five plants of the usable area, measuring (tape) the distance between ground level to the base of the capitulum at the start of maturation (R9). Capitulum size (CS; cm) was measured (tape) from one bract to another; mass of achenes per capitulum (MAC; g) was measured by randomly collecting five capitula from the usable area of the plot (R9). Mean rate of the above five characteristics was thus provided.
Mass of one thousand achenes (MTA; g) was calculated by randomly collecting and weighing of a sample of one thousand achenes. Their productivity (PR; kg ha-1) was determined after a manual harvest of the capitula of the usable area (R9; 103 DAS); humidity was corrected for 11% (bu) and calculated by Equation 1, as suggested by Dalchiavon et al. (2011):
PR = P.[(100-Uob) / (100 – Ud)] .............................................................................Eq. (1)
where: PR is the corrected mass of achene (kg ha-1); P is the field mass (not corrected) of the achenes (kg ha-1); Uob is the humidity of each plot (%) and Ud is the standard humidity (11%).
Data underwent analysis of variance. When F was significant (p<0.05), Tukey´s test was applied for qualitative factors (genotypes) and regression analysis was applied for quantitative factors (plant populations) with SISVAR (Ferreira, 2011). Co-relation matrix was prepared for simple linear co-relationships with regard to two-by-two combinations among the attributes under analysis.
RESULTS AND DISCUSSION
Table 1 summarizes the analyses of variance for plant height (PH), stem diameter (SD), capitulum height (CH), capitulum size (CS), mass of achenes per capitulum (MAC), mass of one thousand achenes (MTA) and productivity of achenes (PR). There was a significant effect for all isolated factors (p<0.05) for all the characteristics studied for genotypes and for plant populations. There was no interaction effect among these factors for any of the characteristics under analysis.
Genotype GNZ NEON showed the highest PH (205.0 cm), whilst AGUARÁ 04 had the lowest, with 170.3 cm (Table 2). There was no difference among the other genotypes (HÉLIO 251, SYN 045 and SYN 3950HO). Similar PH rates were presented in an assay in Campo Novo do Parecis MT Brazil, in the 2013 interim harvest, featuring GNZ NEON AP with 214.0 cm and AGUARÁ 04 with 179.0 cm (Carvalho et al., 2014). Lowest growth rate of the genotypes may be positive, or rather, the capitulum of the plants would be well-sustained and plants would not fall because of bad weather.
Genotypes with the highest and lowest PH also had the greatest and smallest SD (GNZ NEON = 27.6 mm; AGUARÁ 04 = 23.4 mm), as Table 2 shows. In their 2007 assay on sunflowers in Papanduva SC Brazil, Backes et al. (2008) reported PH = 211.0 cm and SD = 25.3 mm in genotype AGUARÁ, whereas PH and SD were 201.0 cm and 28.2 mm respectively for genotype HÉLIO 251, or rather, higher than those in current assay. In studies in Campinas SP Brazil, with 14 sunflower genotypes to estimate correlation rates between agronomic characteristics, Amorim et al. (2008) reported a positive co-relationship between PH and SD (r = 0.84**), corroborated by current assay (r = 0.57**) (Table 3). The above co-relationship is important. In fact, the greatest the PH, the greatest should be its support base (SD) to decrease the possibility of breaking or falling down of the plants.
AGUARÁ 04 (156.4 cm) had the greatest CH whilst HELIO 251 (130.8 cm) had the smallest (Table 2). Since there was a negative co-relationship between CH and SD (r = - 0.46**), MAC (r = - 0.50**) and MTA (r = - 0.38**), CH was affected by its mass (Table 3). On the other hand, Amorim et al. (2008) reported a positive co-relationship between CH and PH (r = 0.99**) and between CH and SD (r = 0.82**), different from rates in current assay.
Table 3 shows a positive co-relationship between CS and MAC (r = 0.57**). However, SYN 045 (15.9 cm) and AGUARÁ 04 (17.8 cm) had the smallest and the largest CS respectively. Genotype SYN 45 had the greatest MAC (42.7 g) and AGUARÁ 04 had the smallest MAC (32.9 g) (Table 2). Co-relationship was positive due to the fact that the genotype AGUARÁ 04 did not differ from GNZ NEON, HÉLIO 251 and SYN 3950HO with regard to CS, and SYN 045 did not differ from GNZ NEON and HÉLIO 251 with regard to MAC.
Genotype SYN 45 had the greatest MAC and MTA (44.0 g), whereas AGUARÁ 04 had the lowest MAC and MTA (31.4 g) (Table 2), confirming co-relation (r = 0.67**).
Lowest CS and greatest MAC and MTA of the genotype SYN 045 may compensate this characteristic by producing achenes with a greater mass, confirmed in current analysis by a greater PR in SYN 045 (1,625.5 kg ha-1) and a lower one in AGUARÁ 04 (1,299.0 kg ha-1). The above suggests that PR is more related to MAC and MTA than to CS, and thus corroborates co-relationships for MAC and MTA with regard to PR (r = 0.34** and r = 0.33**, respectively) (Table 3). An assay conducted in Campo Novo do Parecis MT Brazil, in the 2013 interim harvest, showed that genotype AGUARÁ 04 produced 2,271.0 kg ha-1, with 17.0 cm for CS, whereas genotype SYN 45 had a 15.1% more production, even with CS smaller by 1 cm than in AGUARÁ 04 (Carvalho et al., 2014). Amorim et al. (2008) also reported positive co-relationship between PR and MTA (r = 0.55*).
There was a direct and linear increase in PH with increase in plant populations (Figure 2A), corroborating reports by Castro et al. (2011). Similarly, there was a 0.29 cm increase in PH for each increase of 1,000 plants ha-1. Consequently, PH increased from 181.0 to 189.8 cm when plant population rose from 30,000 to 60,000 plants ha-1. The above fact may be related to inter-species competition for light, causing plant blanching. Competition of light has been reported by Silva and Almeida (1994) in Eldorado do Sul RS Brazil, when they reported PH quadratic increase in four populations (30,000; 45,000; 60,000; 75,000 plants ha-1) due to increase in plant population when measurement occurred in R1 and in R6. On the other hand, in his studies on four populations (27,000; 35,000; 43,000; 51,000 plants ha-1) and four spacings (0.90; 0.75; 0.60; 0.45 m), Orlando (2008) reported that there was a decrease of PH with plant population increase in 0.45 m spacing. The author concluded that other factors, such as water and nutrients, may have had more limiting effects than solar radiation in greater populations.
There was a linear decrease in SD as plant population increased (Figure 2B) from 26.4 to 23.4 mm when plant population rose from 30,000 to 60,000 plants ha-1, with a reduction of 0.1 mm for every increase of 1,000 plants ha-1. Bezerra et al. (2014) reported that the highest inter-species competition, caused by the highest plant population, decreased SD at 42 DAS at the rate of 0.04 and 0.05 mm for each increase of 1,000 plants ha-1, in assays in Fortaleza CE Brazil, and Pentecoste CE Brazil, respectively, with a 0.06 mm decrease at 70 DAS, regardless of place.
Although highest plant population rates were generally related to lodging through PH increase and SD reduction, Silva and Almeida (1994) reported an increase in PH and a decrease in SD when plant population rose, with no lodging or break of plants in the populations studied (30,000; 45,000; 60,000; 75,000 plants ha-1) for genotype Contisol 711. However, Rezende et al. (2003) evaluated two plant populations (40,000 and 60,000 plants ha-1) and six sunflower genotypes and reported differences in the number of lodged plants when there was an increase in plant populations in two genotypes (V 2000 and DK 180), probably attributed to their low tolerance for diseases, enhancing increase in lodged plants. The two studies showed that lodging and plant breaking actually depended on genotypes.
CH increased linearly from 132.7 to 153.3 cm (Figure 2C) when plant population increased. Seeding density increased from 30,000 to 60,000 plants ha-1, with a 0.7 cm increase in CH for every increase of 1,000 plants ha-1.
CS, MAC and MTA (Figures 2D-F) decreased linearly when plant population increased. CS decreased from 18.4 to 15.6 cm when plant population rose from 30,000 to 60,000 plants ha-1, with a 0.1 cm reduction for every increase of 1,000 plants ha-1 (Figure 2D). A similar decrease was reported by Orlando (2008): diameter of the capitulum decreased 0.1 cm for every increase of 1,000 plants ha-1, from 17.9 to 14.5 cm when density oscillated from 27,000 to 51,000 plants ha-1. Silva and Almeida (1994) reported CS decrease according to increase in plant population and concluded that since the capitulum had the smallest size in the highest densities, correlated to stem diameter (r = 0.89), it counterbalanced possible effects of the plant´s greater size and the stem´s smaller diameter. Table 3 also shows that there was a positive co-relationship between CS and SD (r=0.43**).
MAC reached 45.4 g for a population of 30,000 plants ha-1, or rather, 48.3% higher than MAC with 60,000 plants ha-1, and a decrease of 0.49 g for every increase of 1,000 plants ha-1 (Figure 2E). Orlando (2008) also reported a linear reduction in MAC, oscillating between 69.3 and 28.5 g, when plant population rose from 27,000 to 51,000 plants ha-1. In other words, increase in plant population caused a greater inter-species competition and formed achenes with smaller mass. The author verified high co-relationship between MAC and MTA (r = 0.99**), corroborated by current study (r = 0.66**; Table 3).
MMA decreased from 39.7 to 34.3 g when plant population increased from 30,000 to 60,000 plants ha-1, or rather, a reduction of 0.2 g for every increase of 1.000 plants ha-1 (Figure 2F). Castro et al. (2011) also reported a reduction ranging between 58.3 and 43.3 g for densities oscillating between 30,000 and 60,000 plants ha-1. Further, Rizzardi and Silva (1993) in their studies in Eldorado do Sul RS Brazil during the 1989-90 harvest, with four plant populations (30,000; 50,000; 70,000; 90,000 plants ha-1) reported that MMA decreased quadratically as plant population increased, with a 31% reduction when plant population increased from 30,000 to 90,000 plants ha-1, with a minimum point of 94,000 plants ha-1, when MMA reached 42.0 g.
Corroborating data in current study and those by Rizzardi and Silva (1993), Orlando (2008) also registered that MMA responded linearly and inversely to increase in plant population. In the latter´s study, MMA decreased from 59.6 to 49.1 g in populations with 27,000 and 51,000 plants ha-1, respectively. The above revealed a negative effect in the intra-species competition for production and partition of photo-assimilates.
Figure 3 shows PR regression due to plant population by which plant populations was calculated (47,150 plants ha-1), with maximum PR (1,552.6 kg ha-1). Population with 47,150 plants ha-1 is close to that given by Castro et al. (1997), with 45,000 plants ha-1. Quadratic regressions for PR were also obtained in other studies: in their studies on genotype AGUARÁ 04 and 0.45 m spacing, Castro et al. (2011) registered that a population of 38,731 plants ha-1 provided the highest PR (2,312.1 kg ha-1); in their studies in Eldorado do Sul RS Brazil, for the agricultural year 1990-91, on genotype GR 16, short cycle and small size, Rizzardi and Silva (1993) underscored that maximum PR (1,609.5 kg ha-1) was obtained with a population of 56,279 plants ha-1; in Passo Fundo RS Brazil, Rizzardi and Küffel (1993) assessed four plant populations (30,000; 50,000; 70,000; 90,000 plants ha-1) for genotype G18 (short cycle and low stature) and obtained maximum PR (2,370.01 kg ha-1) with a population of 60,838 plants ha-1. It may be perceived that, in every agricultural year and for each genotype cultivated in a different place, a different response was provided. In fact, a standard population that would result in a higher productivity is impossible. A population proper to place and circumstances is more feasible.
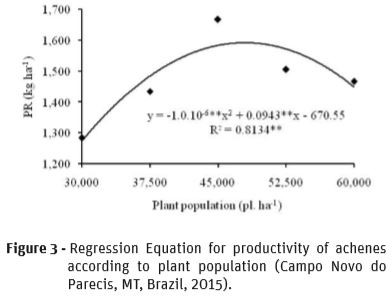
However, linear reduction in PR due to increase in plant population was reported by Rizzardi and Silva (1993). PR was 23 and 17% lower for genotypes DK 180 and GR 10, respectively, when population decreased from 90,000 to 30,000 plants ha-1, and by Orlando (2008) with decrease from 1,835.8 to 1,511.8 kg ha-1 when population increased from 27,000 to 51,000 plants ha-1.
When compared to PR rates described by Rizzardi and Küffel (1993) and by Castro et al. (2011), the low PR in current analysis may be due to accumulated rainfall (752 mm) and mean temperature (between 21.1 and 25.7 °C) during the experimental period (Figure 1) since total evapotranspiration of crops (ETc) lies between 442 and 522 mm (Silva et al., 2007; Cavalcante Júnior et al., 2013). Rainfall excess and temperature rates enhanced the development of the Alternaria spot. In fact, Leite and Amorim (2002) had underscored that temperature around 25 °C and non-continuous leaf wetness between 12 and 24 h triggered the development of the disease. Alternaria species may actually spread spores and cause infections during several short wet periods instead of a single long one.
According to Bauer et al. (2013), great Alternaria severity is proportional to increase in plant population due to low aeration and solar irradiation in the seedling rows. In their study with 0.5m in-between rows, leaf area affected rose from 15.0 to 29.0% when plant population increased from 74,000 to 132,000 plants ha-1, whereas with 1.0 m in-between spacing, leaf area affected increased from 10.1 to 26.4% when population rose from 37,000 to 66,000 plants ha-1. There is actually a strict relationship between disease severity and plant population due to the interference of solar radiation on the canopy and the establishment of a favorable microclimate.
As a rule and regardless of the genotype and density tested in current analysis, the productivity of achenes was higher than Brazilian (1,376.0 kg ha-1) and Mato Grosso (1,348.0 kg ha-1) averages for the 2014-15 (CONAB, 2015) and revealed the viability of the genotypes studied.
CONCLUSIONS
Increase in plant and capitulum height and decrease of stem diameter, capitulum size, mass of achenes per capitulum and mass of one thousand achenes occurred when plant population increased.
Productivity is co-related to mass of achenes per capitulum and to mass of one thousand achenes.
The population of 47,150 plants ha-1 provides greater productivity in achenes.
References
Amorim, A.P.; Ramos, N.P.; Ungaro, M.R.G. and Kiihl, T.A.M. (2008) - Correlações e análise de trilha em girassol. Bragantia, vol. 67, n. 2, p. 307-316. http://dx.doi.org/10.1590/S0006-87052008000200006 [ Links ]
Backes, R.L.; Souza, A.M. de; Balbinot Junior, A.A.; Gallotti, G.J.M. and Bavaresco, A. (2008) - Desempenho de cultivares de girassol em duas épocas de plantio de safrinha no planalto norte catarinense. Scientia Agraria, vol. 9, n. 1, p. 41-48. http://dx.doi.org/10.5380/rsa.v9i1.10131 [ Links ]
Bauer, F.C.; Gonçalves, A.E. and Nagaoka, A.K. (2013) - Severidade de Alternaria helianthi na cultura do girassol em diferentes espaçamentos e população de plantas. Revista de Ciências Agroveterinárias, vol. 12, n. 1, p. 149-154. [ Links ]
Bezerra, F.T.C.; Dutra, A.S.; Bezerra, M.A.F.; Oliveira Filho, A.F. de and Barros, G. de L. (2014) - Comportamento vegetativo e produtividade de girassol em função do arranjo espacial das plantas. Revista Ciência Agronômica, vol. 45, n. 2, p. 335-343. [ Links ]
Castro, C. de; Castiglioni, V.B.R.; Balla, A.; Leite, R.M.V.B. de C.; Karam, D.; Mello, H.C.; Guedes, L. C.A. and Farias J.R.B. (1997) - A cultura do girassol. Londrina: EMBRAPA-CNPSo. 36 p. (EMBRAPA-CNPSo. Circular técnica, 13). [ Links ]
Castro, C. de; Júnior, A. de O.; Oliveira, F.A. de; Leite, R.M.V.B.C. and Rodak B.W. (2011) - Avaliação do arranjo de plantas de girassol. In: Reunião Nacional de Pesquisa De Girassol, 19., Simpósio Nacional Sobre a Cultura do Girassol, 7., 2011, Aracaju. Anais. Aracaju. p. 241-245. [ Links ]
Carvalho, C.G.P. de; Silva, M.F. da; Amabile, R F.; Godinho, V. de P.C.; Oliveira, A.C.B. de; Carvalho, H.W.L. de; Ramos N.P. and Ribeiro J.L. (2014) - Informes da avaliação de genótipos de girassol 2012/2013 e 2013. Londrina: Embrapa Soja. 105 p. (Embrapa Soja. Documentos, 355). [ Links ]
Cavalcante Júnior, E.G.; Medeiros, J.F. de; Mel, T.K. de; Sobrinho, J.E.; Bristot, G. and Almeida B.M. de (2013) - Necessidade hídrica da cultura do girassol irrigado na chapada do Apodi. Revista Brasileira de Engenharia Agrícola e Ambiental, vol. 17, n. 3, p. 261-267. [ Links ]
CONAB (2015) - Acompanhamento da safra brasileira de grãos – safra 2014/2015 - Décimo Segundo Levantamento Setembro/2015. Brasília: Conab - Companhia Nacional de Abastecimento. 139 p. [cit. 2015.09.17]. <http://www.conab.gov.br/OlalaCMS/uploads/arquivos/15_09_11_10_42_03_boletim_graos_setembro_2015.pdf> [ Links ]
Dalchiavon, F.C.; Carvalho, M.P.; Freddi, O.S.; Andreotti, M. and Montanari, R. (2011) - Variabilidade espacial da produtividade do feijoeiro correlacionada com atributos químicos de um Latossolo Vermelho Distroférrico sob sistema de semeadura direta. Bragantia, vol. 70, n. 4, p. 908-916. http://dx.doi.org/10.1590/S0006-87052011000400025 [ Links ]
Dalchiavon, F.C.; Montanari, R.; Andreotti, M.; Dallacort, R. and Souza, M.F.P. (2015) - Relationship between sunflower productivity and soil´s chemical properties by geo-statistical techniques. African Journal of Agricultural Research, vol. 10, n. 35, p. 3525-3532. http://dx.doi.org/10.5897/AJAR2014.9472 [ Links ]
Ferreira, D.F. (2011) - Sisvar: a computer statistical analysis system. Ciência e Agrotecnologia, vol. 35, n. 6, p. 1039-1042. http://dx.doi.org/10.1590/S1413-70542011000600001 [ Links ]
Leite, R.M.V.B.C. and Amorim, L. (2002) - Influência da temperatura e do molhamento foliar no monociclo da mancha de alternaria em girassol. Fitopatologia Brasileira, vol. 27, n. 2, p. 193-200. http://dx.doi.org/10.1590/S0100-41582002000200012 [ Links ]
Orlando, A.F. (2008) - Cultivo de girassol na safrinha no oeste do Paraná: efeitos do espaçamento entre linhas e populações de plantas. Master´s Dissertation in Agronomy - Universidade Estadual do Oeste do Paraná, Marechal Cândido Rondon. 52 p. [ Links ]
Pérez, D.F.; Agudclo, O. and Peña, J. (1989) - Respuesta del girasol a diferentes profundidades de incorporacion del fertilizante. Acta Agronômica, vol. 39, n. 3/4, p. 82-92. [ Links ]
Rezende, A.V.; Evangelista, A.R.; Siqueira, G.R.; Barcelos, A.F.; Rocha, G.P. and Santos R.V. (2003) - Efeito da densidade de semeadura sobre a produtividade e composição bromatológica de silagens de girassol (Helianthus annuus L.). Ciência e Agrotecnologia, vol. 27, n. sp., p. 1672-1678. [ Links ]
Rizzardi, M.A. and Küffel A. (1993) - Influência do arranjo de plantas no rendimento de grãos e óleo e componentes do rendimento de girassol. Ciência Rural, vol. 23, n. 3, p. 287-290. https://doi.org/10.1590/S0103-84781993000300006 [ Links ]
Rizzardi, M.A. and Silva P.R.F. da (1993) - Resposta de cultivares de girassol à densidade de plantas em duas épocas de semeadura. Pesquisa Agropecuária Brasileira, vol. 28, n. 6, p. 675-687. [ Links ]
Silva, M. de L.O.; Faria, M.A. de; Morais, A.R. de; Andrade, G.P. and Lima E.M. de C. (2007) - Crescimento e produtividade do girassol cultivado na entressafra com diferentes lâminas de água. Revista Brasileira de Engenharia Agrícola e Ambiental, vol. 11, n. 5, p. 482-488. http://dx.doi.org/10.1590/S1415-43662007000500006 [ Links ]
Silva, P.R.F. da and Almeida, M.L. de (1994) - Resposta de girassol à densidade em duas épocas de semeadura e dois níveis de adubação. Pesquisa Agropecuária Brasileira, vol. 29, n. 9, p. 1365-1371. [ Links ]
Vogt, G.A.; Balbinot Júnior, A.A. and Souza A.M. de (2012) - Divergência fenotípica em genótipos de girassol. Revista de Ciências Agroveterinárias, vol. 11, n. 1, p. 26-34. [ Links ]
Acknowledgements
The authors would like to thank the Research Group in Phytothecny (GPF) of the Federal Institute of Education, Science and Technology of Mato Grosso (IFMT), campus Campo Novo do Parecis, for its collaboration in current assay. The authors are also grateful to the Brazilian Center for Scientific and Technological Development (CNPq - Process 402022 / 2014-9) for funding Initiation Scientific Research. To the Foundation for Research Support of the State of Mato Grosso (FAPEMAT - Process 460160/2014) by granting scientific training scholarship. Thanks are also due to Embrapa Soja for providing seeds and the required information for the development of current research.
Received/recebido: 2017.06.23
Received in revised form/recebido em versão revista: 2017.11.08
Accepted/aceite: 2017.11.08

