










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias vol.41 no.1 Lisboa mar. 2018
https://doi.org/10.19084/RCA17100
ARTIGO
Regeneração natural em área sob domínio de bambu, no sul do Brasil
Natural regeneration in area under domain of bamboo, in the south of Brazil
Roselene Marostega Felker1,2*, Ana Paula Moreira Rovedder1,2, Solon Jonas Longhi2, Elias Frank Araujo3, Maureen de Moraes Stefanello1,2, Jose Carlos Corrêa da Silva Júnior1,2, Djoney Procknow1,2, Rafaela Badinelli Hummel1,2, Bruna Balestrin Piaia1,2, Betina Camargo1,2, Marcela Peuckert Kamphorst Leal da Silva1,2 e Lucas Donato Toso1,2
1Núcleo de Estudos e Pesquisas em Recuperação de Áreas Degradadas (NEPRADE). Universidade Federal de Santa Maria (UFSM), Santa Maria, Rio Grande do Sul, Brasil
2Universidade Federal de Santa Maria (UFSM), Santa Maria, Rio Grande do Sul, Brasil
3CMPC Celulose Riograndense, Guaíba, Rio Grande do Sul, Brasil
(*E-mail: marostegafelker@gmail.com)
RESUMO
Conhecer a regeneração florestal em áreas dominadas por bambu pode auxiliar no gerenciamento das espécies nestas áreas, assim como auxiliar em propostas de recuperação e restauração destes ambientes. Dessa forma, o presente trabalho objetivou conhecer a estrutura, composição florística, mecanismos de dispersão, tolerância à sombra e hábito de vida das espécies da regeneração natural presentes em área sob domínio da espécie exótica Bambusa tuldoides Munro. O estudo foi desenvolvido em área de floresta estacional, município de Eldorado do Sul, Rio Grande do Sul, Brasil. Foram demarcadas 30 parcelas de 12x12 m, totalizando 0,432 ha de amostragem. Foram medidos e identificados todos os indivíduos presentes na classe I: altura maior ou igual a 30 cm e menor que 130 cm (30≤ H<130 cm) e na classe II: circunferência à altura do peito menor ou igual a 5 cm (CAP≤5). Foram observadas baixa densidade de indivíduos e espécies, principalmente na Classe II. Ocotea puberula, Myrsine umbellata, Trichilia elegans e Cupania vernalis apresentaram maior distribuição e relação com a área de bambus. Quanto à ecologia das espécies da regeneração, a maiorias destas podem ser classificadas como tolerantes à sombra (53%), de dispersão zoocórica (87%) e hábito de vida arbóreo (54%). Conclui-se que na área de superabundância de Bambusa tuldoides analisada, a classe da regeneração apresenta baixa densidade de espécies, indivíduos de pequeno porte, tolerantes à sombra e de dispersão zoocórica. Estas informações, devem ser consideradas para futuras ações de manejo e recuperação no local de estudo.
Palavras-chave: Bambusa tuldoides; Mata atlântica; Restauração.
ABSTRACT
Understand the native forest regeneration process in areas dominated by bamboo can be a support to the management of these species, as well as assist in proposals for recovery and restoration of these environments. Therefore, this study aimed to evaluate concomitantly the structure, floristic composition, mechanisms of dispersion, shade tolerance and lifeforms of the natural regenerating species present in the area under dominance of exotic species Bambusa tuldoides Munro. The study was carried out in seasonal forest, in the municipality of Eldorado do Sul, state of Rio Grande do Sul, Brazil. Thirty plots of 12x12 m (0.432 ha) were sampled and the plants divided into two classes. In Class I were measured and identified all the individuals with height greater or equal to 30 cm and less than 130 cm (30 ≤ H <130 cm) and in the Class II all the individuals with a circumference at breast height less than or equal to 5 cm (CBH≤5). Low density of individuals and species were observed, mainly in Class II. Ocotea puberula, Myrsine umbellate, Trichilia elegans and Cupania vernalis showed a greater distribution and relation with the bamboos area. About ecology, the majority of regenerating species can be classified as tolerant to shade (53%), zoocoric dispersion (87%) and arboreal lifeform (54%). It is concluded that in the area of superabundance of Bambusa tuldoides, the regeneration class (Class I) presents low density of species, small individuals, tolerant to shade and with zoocoric dispersion. It information should be considered for future management and recovery actions at the site.
Kewords: Bambusa tuldoides; Atlantic forest; Restauration.
INTRODUÇÃO
Recentemente, a superabundância de bambus em áreas florestais têm sido motivo de preocupação. No sudeste brasileiro, mapeamentos revelaram que partes consideráveis da Mata Atlântica estão dominadas por maciços de bambus lenhosos (Araujo, 2008; Lima et al., 2012). Situação análoga pode ser observada na Amazônia (Carvalho et al., 2013) e em florestas no continente asiático (Kudo et al., 2011; Fukushima et al., 2015; Shinohara e Otsuki, 2015).
Diante da fragmentação atual das florestas, a superabundância do bambu pode ser considerada como grande ameaça à manutenção do equilíbrio ecológico. A forma de reprodução vegetativa por meio de rizomas, o rápido crescimento e a elevada densidade de colmos por área tornam os bambus altamente competitivos, alterando a estrutura dos ecossistemas naturais (Santos et al., 2012). Quando não manejados, as espécies de bambus alastrantes ou invasoras, devido a seus aglomerados estruturais, podem impedir o desenvolvimento da regeneração e comprometer a sucessão florestal (Griscom e Ashton, 2006).
As principais causas da ocorrência de espécies superabundantes estão ligadas à fragmentação florestal, desmatamentos e incêndios florestais (Lima et al., 2012). Ambientes perturbados são ambientes ideais para o crescimento e proliferação de espécies pioneiras agressivas, como o bambu (Griscom e Ashton, 2006). Soma-se a isso, a introdução para fins econômicos (Lima et al., 2007) e o abandono do cultivo sem adequado monitoramento, caso da espécie exótica Bambusa tuldoides Munro (Poaceae: Bambusoideae) no sul do Brasil. B. tuldoides, oriunda do continente asiático, foi introduzida no Rio Grande do Sul para abastecimento da indústria de papel e celulose no final do século XVIII (ENE, 2016). Com o término da atividade produtiva, as áreas foram abandonadas e atualmente compreendem extensas áreas dominadas por essa espécie de bambu.
Diante desta problemática, o objetivo da pesquisa foi analisar a diversidade e estrutura da regeneração natural existente em áreas sob domínio de B. tuldoides, assim como conhecer os mecanismos de dispersão, tolerância à sombra e hábito de vida das espécies diretamente influenciadas pela superabundância deste bambu, a fim de contribuir com o conhecimento da área para futuras estratégias de manejo e recuperação.
MATERIAIS E MÉTODOS
Área de estudo
O estudo foi desenvolvido em áreas sob domínio da espécie B. tuldoides decorrentes de plantios comerciais de meados do século XVIII, na zona rural do município de Eldorado do Sul, sul do Brasil. B. tuldoides é uma espécie exótica que não apresenta características de invasora alastrante (Shirasuna et al., 2013). No entanto, são entouceirantes e formadoras de adensamentos, o que pode interferir no estabelecimento e dinâmica da vegetação (Felker et al., 2017).
A área está localizada em zona ciliar, e inserida em uma matriz produtiva de Eucalyptus sp. com zonas de amortecimento de matas nativas. A área localiza-se na Depressão Central do Rio Grande do Sul, Bioma Mata Atlântica, onde a formação florestal característica é a Floresta Estacional (IBGE, 2012). Os solos são classificados como Cambissolos háplicos, de origem granítica, com presença de material de origem nos horizontes A e B. O clima é definido como subtropical úmido (ST UM), com temperatura média anual de 19,2ºC, precipitação pluvial anual de 1708 mm e deficiência hídrica anual de 11 mm (Maluf, 2000).
Amostragem dos dados
Foram demarcadas 30 parcelas de 12x12 m, totalizando 0,432 ha, em área de domínio de B. tuldoides. Para estudo da regeneração natural, foram medidos e identificados indivíduos presentes em duas classes de tamanho. Na classe I, foram identificados indivíduos com altura maior ou igual a 30 cm e menor que 130 cm (30≤ H<130 cm) e medidos o diâmetro à altura do solo (DAS) e altura. Na classe II, foram identificados todos os indivíduos com circunferência à altura do peito menor ou igual a 5 cm (CAP≤5) e medida a circunferência e altura de cada indivíduo.
A nomenclatura botânica utilizada foi baseada na Lista de Espécies da Flora do Brasil (JBRJ, 2012) e a delimitação das famílias através do sistema de classificação APG IV (2016). Espécies não identificadas in loco foram encaminhadas ao Herbário do Departamento de Ciências Florestais (HDCF) da Universidade Federal de Santa Maria (UFSM) para posterior identificação.
Análise dos dados
Os estimadores da estrutura horizontal da floresta foram analisados de acordo com Mueller-Dombois e Ellenberg (1974), a diversidade através dos índices de Shannon (H´), e Equabilidade de Pielou (J´) (Magurran, 1988) e a dominância através do índice de Simpson (C) (Brower e Zarr, 1984), calculados com o software PAST, versão 1.79 (Hammer et al., 2004). Na determinação dos grupos florísticos, foi utilizado o método Twinspan (Hill, 1980), através de matriz com dados de presença e ausência. A análise foi realizada no programa PC-ORD para Windows versão 4.14 (McCune e Mefford, 1999).
A distribuição diamétrica e de altura foi determinada segundo Felfili e Rezende (2003). A classificação ecológica das espécies foi realizada através de mecanismos de dispersão (zoocóricas, anemocóricas e autocóricas) (Van der Pijl, 1982); da estratificação (sub-bosque, dossel e emergentes) (Oliveira-Filho et al., 1994); necessidades de luz (tolerantes à sombra, dependentes de luz e pioneiras (Swaine e Whitmore, 1988) e hábito de vida (arbusto, arvoreta e árvore).
RESULTADOS
Na classe I foram amostrados 233 indivíduos (539,3/ha) pertencentes a 39 espécies e 20 famílias botânicas; na classe II foram amostrados 56 indivíduos (129,6/ha) de 19 espécies e 15 famílias botânicas (Quadro 1).
Das 20 famílias botânicas amostradas na classe de regeneração I, Myrtaceae foi a mais representativa, com 9 espécies, seguida de Lauraceae, Rubiaceae e Meliaceae (três espécies cada), Cardiopteridaceae, Rutaceae, Sapindaceae, Celastraceae e Primulaceae (duas espécies cada). As demais famílias apresentaram apenas uma espécie. As espécies mais representativas nesta classe, observadas com maior densidade e frequência, foram Ocotea puberula, Myrsine umbellata e Trichilia elegans (Quadro 1).
Para a classe II, observou-se diminuição no número total de indivíduos e espécies em relação à classe I. As famílias Myrtaceae, Meliaceae, Sapindaceae e Celastraceae foram as mais representativas, com duas espécies cada. As demais 11 famílias apresentaram uma única espécie. A espécie com maior ocorrência foi Myrsine umbellata, seguida de Cabralea canjerana, Cupania vernalis e Trichilia elegans.
Ocorreram 15 espécies comuns em ambas as classes de regeneração. São elas, pela ordem de densidade: Ocotea puberula, Mysine umbellata, Trichilia elegans, Cupania vernalis, Cabralea canjerana, Maytenus sp., Myrcia sp., Maytenus dasyclada, Gymnanthes klotzschiana, Matayba elaeagnoides, Erythroxylum argentinum, Rubiaceae sp., Cytrus sp., Chrysophylum marginatum e Eugenia hiemalis. Por outro lado, 24 espécies somente ocorreram com indivíduos na classe I e quatro espécies somente na classe II (Quadro 1).
Na análise de agrupamento, foram formados dois grupos na classe I (Figura 1). O primeiro grupo (G1) foi originado na primeira divisão e apresentou autovalor de 0,5106. O segundo grupo (G2) foi originado da segunda divisão e apresentou autovalor de 0,3925. O G1 apresenta como espécie indicadora Trichilia elegans e como preferenciais Sorocea bonplandii e Trichilia elegans. O G2 traz como espécies indicadoras Ocotea puberula e Myrsine umbellata e como preferenciais Maytenus sp., Zanthoxylum rhoifolium, Cupania vernalis, Trichilia elegans, Nectandra megapotamica, Tibouchina sp., Myrcia sp., e Ocotea puberula.
Para a classe II, dois grupos florísticos foram separados, originários da primeira divisão, com autovalor de 0,8479 (Figura 2). O G1 apresentou Myrsine umbellata como espécie indicadora e como preferenciais Trichilia elegans e Myrsine umbellata. O G2 tem como espécies indicadoras Cupania vernalis e Strychnos brasiliensis e como preferenciais Cupania vernalis e Strychnos brasiliensis.
O índice de Shannon (H) e a equabilidade de Pielou apresentaram valores de diversidade próximos, assim como o índice de dominância de Simpson que ficou próximo a 1 nas duas classes de amostragem (Quadro 2).

Na distribuição diamétrica das espécies da classe I (Figura 3a) verifica-se uma maior concentração de indivíduos nas menores classes de diâmetro acima do solo (0,3-0,5 cm de DAS), diminuindo para as classes de maior tamanho (>0,81 cm de DAS). Já, para a classe II (Figura 2b) observa-se irregularidade na distribuição diamétrica, com maior número de indivíduos no intervalo de 1,7 a 3,4 cm de CAP e posterior decréscimo nos CAP superiores à 3,4 cm.
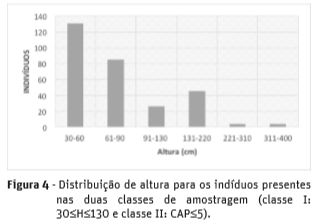
Com relação a altura verifica-se que indivíduos presentes no intervalos de 30 à 60 cm são dominantes na área, enquanto àqueles com altura superior a 220 cm são escassos (Figura 4).
Quanto à classificação ecológica (Quadro 3), 53% das espécies presentes na área, são tolerantes à sombra na fase inicial de seu desenvolvimento; 27% são pioneiras típicas e 20% dependentes de luz em alguma fase de seu desenvolvimento.
Para os mecanismos de dispersão, 87% das espécies presentes são zoocóricas, 8% anemocóricas, 3% autocóricas e 2% seriam autocóricas ou anemocóricas. Quanto à estrutura vertical dos regenerantes, verificou-se que 63% dos indivíduos presentes na amostragem são espécies pertencentes ao sub-bosque florestal, 32% frequentes do dossel e 5% emergentes. Com relação ao hábito de vida, 54% foram classificados como árvore, 23% como arvoreta e 23% como arbusto.
DISCUSSÃO
Observou-se uma diminuição de indivíduos e espécies da classe I para a classe II. Das 39 espécies encontradas na classe I, menos da metade (15 espécies), foram representadas na classe II. Dentre estas, destacam-se Ocotea puberula, Mysine umbellata, Trichilia elegans, Cupania vernalis e Cabralea canjerana, observadas em grande densidade e frequência na classe I. A maioria das espécies com densidade média e baixa representatividade (em termos de frequência) na classe I não avançaram para a classe II (Quadro 1). Segundo Chazdon (2008), o tipo de manejo e a colonização por espécies agressivas pode alterar a trajetória sucessional de uma área. Se uma espécie estiver representada em apenas uma classe de regeneração, existe o risco de ocorrer extinção local (Elias et al., 2017). Silvério et al (2010) e Silveira (2005) destacam que a presença do bambu pode causar diminuição na densidade de indivíduos de classes maiores por competir por recursos, assim como causar danos mecânicos influenciando o crescimento dos regenerantes (Griscom e Ashton, 2006).
Com relação aos grupos florísticos existentes na área, observou-se a formação de dois grupos para a classe I (Figura 1). No grupo 1 (G1) Trichilia elegans foi a espécie indicadora. Ecologicamente, Trichilia elegans é classificada como secundária tardia, típica de subbosque (Venzke, 2012; Bündchen et al., 2015). No segundo grupo formado (G2), Ocotea puberula e Myrsine umbellata foram indicadoras. Estas espécies podem ser vistas frequentemente colonizando áreas alteradas como o sub-bosques de Eucalyptus sp. e Pinus sp. (Carvalho, 2002). Segundo Howe (1993), Ocotea puberula produz grandes quantidades de sementes como estratégia de colonização e estas germinam facilmente em qualquer tipo de solo (Backes e Irgang, 2002), mesmo em ambientes degradados (Souza e Moscheta, 1991). Os dois grupos apresentam espécies, em sua maioria, características de ambientes sombreados, o que explica sua amostragem em ambientes com presença de bambu. Este padrão de regenerantes tolerantes à sombra, em áreas dominadas por bambus, também foi observado em trabalhos de Giehl et al. (2007) e Santos et al. (2012).
Para a classe II, Myrsine umbellata é indicadora no G1 e Cupania vernalis e Strychnus brasiliensis no G2 (Figura 2). Myrsine umbellata é uma espécie agressiva podendo ocorrer em matas primárias até capoeirões (BACKES e IRGANG, 2002). Esta espécie foi observada em alta densidade na Floresta Ombrófila sob domínio de bambu no Paraná (Sanquetta, 2007). No G2, Cupania vernalis e Strychnus brasiliensis são espécies mais dependentes de luz. S. brasiliensis pode ser classificada como espécie pertencente ao dossel florestal (Leyser et al., 2012); C. vernalis é considerada espécie de luz difusa atingindo estágios avançados rapidamente na sucessão (Marchiori, 1997). A espécie responde a diferentes intensidades de luz durante o primeiro ano de crescimento (Siminski e Paulilo, 2004). Cupania vernalis e Trichilia elegans também foram registradas em grande densidade e frequência em área sob influência de Merostachys multiramea Hack., em zona de transição entre Floresta Ombrófila Mista e Floresta Estacional no Sul do Brasil (Santos et al., 2012). Estas espécies, verificadas com maior representatividade, são de grande importância, uma vez que podem ser espécies-chave no processo de restabelecimento da vegetação, assim como podem servir como fontes de entrada de propágulo, alimento para frugívoros e poleiros. Em áreas alteradas, pequenos núcleos de espécies podem contribuir no processo de recuperação destes ambientes (Rother et al., 2009).
A equabilidade de Pielou (J) indica que poucas espécies estão bem representadas na floresta, indicado também pela alta dominância de espécies em ambas as classes (Quadro 2). Essa dominância se refere as espécies Ocotea puberula, Myrsine umbellata, Trichilia elegans e Cupania vernalis. Pela alta expressividade e formação de grupos florísticos, estas espécies podem ser as mais adaptadas às condições locais.
Silvério et al. (2010) sugere que a ocupação do espaço e a redução da incidência luminosa causada pelas touceiras do bambu dificultam a germinação das sementes e o estabelecimento das plântulas, selecionando as espécies mais tolerantes à sombra e assim modificando a composição florística e a estrutura da vegetação. Tais características podem estar favorecendo o estabelecimento dessas espécies, o que reforça a necessidade do desenvolvimento de novos estudos que avaliem o potencial de espécies com comportamento similar às supracitadas, para o plantio em áreas ocupadas por bambu.
Quanto à distribuição diamétrica (Figura 3a, Figura 3b), observou-se uma maior concentração de indivíduos nas classes menores, diminuindo para as maiores. Florestas nativas geralmente caracterizam-se por apresentar maior quantidade de indivíduos nas classes de tamanhos menores, diminuindo com o aumento das classes (Lima e Leão, 2013). No entanto, na classe II, onde foi avaliado CAP, se observa um padrão diamétrico irregular. Cordeiro et al. (2007) em estudo em área de bambus, encontrou esse mesmo padrão, com a maioria da vegetação (cerca de 83%) concentrada nas menores classes de tamanho. Para Marimon et al. (2001), este tipo de distribuição pode indicar falhas no recrutamento, pois as espécies regenerantes não estão conseguindo se estabelecer e assim alcançar classes superiores.
Com relação a altura (Figura 4) pode-se observar que a maioria dos indivíduos avaliados se encontra na faixa de 30 a 90 cm de altura. Indivíduos com alturas superiores à 220 cm equivalem à apenas 2,7% da população amostrada (Figura 3). Ziller e Galvão (2002) comentam que dentre os impactos que as espécies mais agressivas causam, uma destas é causar interferência na densidade e porte da vegetação. Estas irregularidades na distribuição diamétrica e na altura podem estar relacionados à desestruturação do fragmento, decorrente da pressão exercida pelo bambu. As interferências podem estar relacionadas à estresse físico ou fisiológico (Lima et al., 2012).
Quanto às síndromes de dispersão, existiu a predominância de espécies zoocóricas, tolerantes à sombra e pertencentes ao sub-bosque (Quadro 3). Santos et al., (2012), em área de influência de Merosthachys multiramea, também observou predominância de espécies com dispersão zoocórica.
Verificou-se que 53% das espécies presentes na área são tolerantes à sombra na fase inicial de seu desenvolvimento, 27% são pioneiras típicas e 20% dependentes de luz em alguma fase de seu desenvolvimento. Silvério et al. (2010) sugere que a ocupação do espaço e a redução da incidência luminosa causada pelas touceiras do bambu pode dificultar a germinação das sementes e o estabelecimento das plântulas, selecionando as espécies mais tolerantes à sombra e assim modificando a composição florística e a estrutura da vegetação.
CONCLUSÃO
A classe da regeneração encontra-se restrita a indivíduos de pequeno porte, tolerantes à sombra e com dispersão zoocórica. Ocotea puberula, Myrsine umbellata, Trichilia elegans e Cupania vernalis, por sua ecologia e representatividade, devem ser consideradas para futuras ações de manejo e recuperação de área dominadas por bambus na região.
Referências bibliográficas
APG. (2016) - An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants IV. Botanical Journal of the Linnaean Society, vol. 181, n. 1, p. 1-20. http://dx.doi.org/10.1111/boj.12385 [ Links ]
Araujo, L.S. (2008) - Padrões e condicionantes da dinâmica da paisagem na floresta com bambus do Parque Estadual Intervales, SP. Tese de doutoramento. São Paulo, Escola Superior de Agricultura Luiz de Queiroz. 126 p. [ Links ]
Backes, P. & Irgang, B. (2002) - Árvores do sul: guia de identificação e interesse ecológico. Santa Cruz do Sul: Clube da Árvore. 326 p. [ Links ]
Brower, J.E. & Zar, J.H. (1984) - Field & laboratory methods for general ecology. 2. ed. Dubuque, Iowa: Wm. C. Brown Publishers, 226 p. [ Links ]
Bündchen, M.; Boeger, M.T.B. & Reissmann, C.B. (2015) - Estrutura foliar de espécies lenhosas de dossel e sub-bosque em uma floresta subtropical do sul do Brasil. Iheringia, vol. 70, n. 1, p. 105-114. [ Links ]
Carvalho, A.L. de; Bruce W.; Nelson, B.W.; Bianchini, M.C.; Plagnol, D.; Kuplich, T.M. & Daly, D.C. (2013) - Bamboo-Dominated Forests of the Southwest Amazon: Detection, Spatial Extent, Life Cycle Length and Flowering Waves. Plos One, vol. 8, n. 1, e54852. https://doi.org/10.1371/journal.pone.0054852 [ Links ]
Carvalho, P.E. (2002) - Canela guaicá. Colombo, PR: EMBRAPA, circular técnica, 62, 11 p. [ Links ]
Chazdon, R.L. (2008) - Chance and determinism in tropical forest succession. In: Carson, W. & Schnitzer, S.A. (Eds.) - Tropical forest community ecology. Wiley-Blackwell Publishing, Oxford, p. 384-408. [ Links ]
Chazdon, R. (2012) - Regeneração de florestas tropicais. Boletim do Museu Paraense Emílio Goeldi, Ciências Naturais, vol. 7, n. 3, p. 195-218. [ Links ]
Cordeiro, A.O.O.; Soares, S.M.P.; Morais, L.E.; Ribeiro, J.H.C.; Garcia, P.O. & Lobo Faria, P.C. (2007) - Fragmento florestal dominado por Phyllostachys aurea Carrière ex Rivière & C. Rivière (Poaceae), no campus da Universidade Federal de Juiz de Fora, MG. In: Congresso de ecologia do Brasil, Caxambu, MG, Sociedade de Ecologia do Brasil, p. 1-2. [ Links ]
Elias, F.; Marimon, B.S.; Neves, E.C.; Morandi, P.S., Reis, S.M.; Mews, H.A. & Marimon-júnior, B. (2015) - Regeneração de espécies lenhosas sob a influência do bambu Actinocladum verticillatum (Nees) McClure ex Soderstr. (Poaceae) em cerradão e cerrado típico na transição Cerrado-Amazônia. Revista Brasileira de Biociências, vol. 13, n. 2, p. 68-78. [ Links ]
Ene, J. (2016) - Antiga fábrica de papel de bambu de bom retiro de Guaíba. [cit. 2017-03-01]. <http://agrobambu.blogspot.com.br/> [ Links ]
Felfili, J.M. & Rezende, R.P. (2003) - Conceitos e métodos em fitossociologia. Comunicações Técnicas Florestais. Universidade de Brasília, Departamento de Engenharia Florestal, Brasília, vol. 5, n. 1. [ Links ]
Felker, R.M.; Rovedder, A.P.; Longhi, S.J.; Frank, E.A.; Stefanello, M.M. & Peccatti, A. (2017) - Impacto de Bambusa tuldoides Munro (Poaceae) na regeneração florestal. Cerne, vol. 23, n. 2, p. 275-282. http://dx.doi.org/10.1590/01047760201723022297 [ Links ]
Fukushima, K.; Usui, N.; Ogawa, R. & Tokuchi, N. (2015) - Impacts of moso bamboo (Phyllostachys pubescens) invasion on dry matter and carbon and nitrogen stocks in a broad-leaved secondary forest located in Kyoto, western Japan. Plant Species Biology, vol. 30, n. 2, p. 81–95. http://dx.doi.org/10.1111/1442-1984.12066 [ Links ]
Giehl, E.L.H.; Athayde, E.A.; Budke, J.C.; Gesing, J.P.A.; Eisinger, S.M. & Canto-Dorow, T.S. (2007) - Espectro e distribuição vertical das estratégias de dispersão de diásporos do componente arbóreo em uma floresta estacional no sul do Brasil. Acta Botanica Brasilica, vol. 21, n. 1, p. 137-145. http://dx.doi.org/10.1590/S0102-33062007000100013 [ Links ]
Griscom, B.P. & Ashton, P.M.S. (2006) - A self-perpetuating bamboo disturbance cycle in a neotropical forest. Journal of Tropical Ecology, vol. 22, p. 587–597. https://doi.org/10.1017/S0266467406003361 [ Links ]
Hammer, O.; Harper, D.A. & Ryan, P.D. (2004) - AST -Paleontological Statistics. Versão 1.12. <http://www.folk.uio.no/ohammer/past> [ Links ].
Hill, M.O. & Gauch J.R. (1980) - Detrended Correspondence analysis: an improved ordination technique. Vegetatio, vol. 42, n. 1-3, p. 47-58. [ Links ]
Holz, C.A. & Veblen, T.T. (2006) - Tree regeneration responses to Chusquea montana bamboo die-off in a subalpine Nothofagus forest in the southern Andes. Journal of Vegetation Science, vol. 17, n. 1, p. 19-28. http://dx.doi.org/10.1111/j.1654-1103.2006.tb02419.x [ Links ]
Howe, H.F. (1993) - Specialized and generalized dispersal systems: where does "the paradigm'' stand? Vegetation, vol. 107, n. 1, p. 3-13. http://dx.doi.org/10.1007/BF00052208 [ Links ]
IBGE (2012) - Manual técnico da vegetação brasileira. Rio de Janeiro, RJ – Brasil, Instituto Brasileiro de Geografia e Estatística. [cot. 2017.08.20]. <ftp://geoftp.ibge.gov.br/documentos/recursos_naturais/manuais_tecnicos/manual_tecnico_vegetacao_brasileira.pdf> [ Links ]
JBRJ (2012) - Lista de Espécies da flora do Brasil. Jardim Botânico do Rio de Janeiro. [cit. 2016-06-12].<www.floradobrasil.jbrj.gov.br/> [ Links ].
Kudo, G.; Amagai, Y.; Hoshino, B. & Kaneko, M. (2011) - Invasion of dwarf bamboo into alpine snow-meadows in northern Japan: pattern of expansion and impact on species diversity. Ecology and Evolution, vol. 1, n.1, p.85–96. https://dx.doi.org/10.1002/ece3.9 [ Links ]
Leyser, G.; Zanin, E.M.; Budke, J.C.; Mélo, M.A. de & Henke-Oliveira, C. (2012) - Regeneração de espécies arbóreas e relações com componente adulto em uma floresta estacional no vale do rio Uruguai, Brasil. Acta Botanica Brasilica, vol. 26, n. 1, p. 74-83. http://dx.doi.org/10.1590/S0102-33062012000100009 [ Links ]
Lima, J.P.da C. & Leão, J.R.A. (2013) - Dinâmica de crescimento e distribuição diamétrica de fragmentos de florestas nativa e plantada na Amazônia sul ocidental. Floresta Ambiente, vol. 20, n. 1, p. 70-79. http://dx.doi.org/10.4322/floram.2012.065 [ Links ]
Lima, G.R.A.F; Rother, D.C.; Araujo, L.S.; Gandolfi, S. & Rodrigues, R.R. (2007) - Bamboo dominated gaps in the Atlântica rain Forest: impacts on vegetation structure and species diversity. In: Congresso de ecologia do Brasil, Caxambu, MG. Sociedade de Ecologia do Brasil, 1285 p. [ Links ]
Lima, R.A.F.; Rother, D.C.; Muler, A.E.; Lepsch, I.F. & Rodrigues, R.R. (2012) -Bamboo overabundance alters forest structure and dynamics in the Atlantic Forest hotspot. Biological Conservation, vol. 147, n. 1, p.32–39. http://dx.doi.org/10.1016/j.biocon.2012.01.015 [ Links ]
Magurran, A.E. (1988) - Ecological diversity and its measurements. Princeton: Princeton University Press, 179 p. [ Links ]
Maluf, J.R.T. (2000) - Nova classificação climática do Estado do Rio Grande do Sul. Revista Brasileira de Agrometeorologia, vol. 8, n. 1, p. 141-150. [ Links ]
Marchiori, J.N.C. (1997) - Dendrologia das angiospermas: leguminosas. Santa Maria: UFSM, 200 p. [ Links ]
Marimon, B.S.; Felfili, J.M.; Lima, E.S. & Rodrigues, A.J. (2001) - Distribuição de circunferências e alturas em três porções da mata de galeria do Córrego Bacaba, Nova Xavantina-MT. Árvore, vol. 25, n. 3, p. 335-343. [ Links ]
McCune, B. & Mefford, M.J. (1999) - PC-ORD version 4.14: Multivariate analysis of ecological data. Oregon: Glaneden Beach: MjM Software Design, 237 p. [ Links ]
Mueller-Doimbois, D. & Ellenberg, H. (1974) - Aims and methods of vegetation ecology. New York: John Wiley & Sons. 547 p. [ Links ]
Oliveira-Filho, A.T.; Vilela, E.A.; Carvalho, D.A. & Gavilanes M.L. (1994) - Effects of soils and topography on the distribution of tree species in a tropical riverine forest in south-eastern Brazil. Journal of Tropical Ecology, vol. 10, n. 4, p. 483-508. https://doi.org/10.1017/S0266467400008178 [ Links ]
Rother, D.; Rodrigues, R.R. & Pizo, M.A. (2009) - Effects of bamboo stands on seed rain and seed limitation in a rainforest. Forest Ecology and Management, vol. 257, n. 3, p. 885-892. https://doi.org/10.1016/j.foreco.2008.10.022 [ Links ]
Sanquetta, C.R.; Dalla Corte, A.P.; Vulcanis, L. & Berni, D.M. (2005) - Sobrevivência de mudas de Araucaria angustifolia Bert. O. Ktze. plantadas em um experimento de controle de taquaras (Bambusoideae) no sul do Paraná, Brasil. Floresta, vol. 35, n. 1, p. 127-135. [ Links ]
Santos, S.C. dos; Budke, J.C. & Muller, A. (2012) - Regeneração de espécies arbóreas sob a influência de Merostachys multiramea Hack. (Poaceae) em uma floresta subtropical. Acta Botânica Brasilica, vol. 26, n. 1, p. 218-229. http://dx.doi.org/10.1590/S0102-33062012000100021 [ Links ]
Shinohara, Y. & Otsuki, K. (2015) - Comparisons of soil-water content between a Moso bamboo (Phyllostachys pubescens) forest and an evergreen broadleaved forest in western Japan. Plant Species Biology, vol. 30, n. 3, p. 96–103. http://dx.doi.org/10.1111/1442-1984.12076 [ Links ]
Shirasuna, R.T.; Filgueiras, T. de S. & Barbosa, L.M. (2013) - Poaceae do Rodoanel Mario Covas, Trecho Sul, São Paulo, SP, Brasil: florística e potencial de uso na restauração de áreas degradadas. Hoehnea, vol. 40, n. 3, p. 521-536. [ Links ]
Silveira, M. (2005) - A floresta aberta com bambu no sudoeste da Amazônia: padrões e processos em múltiplas escalas. Rio Branco: Edufac, 127 p. [ Links ]
Silvério, D.V.; Mews, H.A.; Lenza, E. & Marimon, B.S. (2010) - Impactos do agrupamento do bambu Actinocladum verticillatum (Nees) McClure ex Soderstr. (Poaceae) sobre a vegetação lenhosa de duas fitofisionomias de Cerrado na transição Cerrado-Floresta Amazônica. Acta Amazonica, vol. 40, n. 2, p. 347-356. http://dx.doi.org/10.1590/S0044-59672010000200013 [ Links ]
Siminski, A. & Paulilo, M.T.S. (2004) - Resposta de Cupania vernalis Cambess (Sapindaceae) a variação na intensidade de luz. Insula, vol. 33, p. 101-110. [ Links ]
Souza, L.A. & Moscheta, I.S. (1991) - Estrutura da flor e antese de Ocotea puberula (Rich.) Nees - Lauraceae. In: Congresso nacional de botânica, Goiânia: Sociedade Botânica do Brasil / Universidade Federal de Góias 359 p. [ Links ]
Swaine, M.D. & Whitmore, T.C. (1988) - On the definition of ecological species groups in tropical rain forests. Vegetatio, vol. 75, n. 1-2, p. 81-86. [ Links ]
Tabarelli, M. & Mantovani, W. (1999) - Clareiras naturais e a riqueza de espécies pioneiras em uma floresta Atlântica Montana. Revista Brasileira de Biologia, vol. 59, n. 2, p. 251-261. http://dx.doi.org/10.1590/S0034-71081999000200009 [ Links ]
Van der Pijl, L. (1982) - Principles of dispersal in higher plants. Berlin: Springer-Verlag, 1982. 162 p. [ Links ]
Venzke, T.S. (2012) - Florística de comunidades arbóreas no Município de Pelotas, Rio Grande do Sul. Revista Rodriguésia, vol. 63, n. 3, p. 571-578. http://dx.doi.org/10.1590/S2175-78602012000300008 [ Links ]
Ziller, S.R. & Galvão, F. (2002) - A degradação da Estepe Gramíneo-Lenhosa no Paraná por contaminação de Pinus elliottii e P. taeda. Revista Floresta, vol. 32, n. 1, p. 41-47. http://dx.doi.org/10.5380/rf.v32i1.2348 [ Links ]
Recebido/received: 2017.04.21
Recebido em versão revista/received in revised form: 2017.09.27
Aceite/accepted: 2017.09.28

