










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias vol.42 no.3 Lisboa set. 2019
https://doi.org/10.19084/RCA.17306
ARTIGO
Metabolismo fotossintético em mudas de Pereskia aculeata Plum. propagadas por estaquia sob diferentes disponibilidades luminosas
Photosynthetic metabolism in Pereskia aculeata Plum. seedlings propagated by cutting under different light availabilities
Cleberton Correia Santos*, Neder Martins Lima, Maria do Carmo Vieira, Néstor Antonio Heredia Zárate e Silvana de Paula Quintão Scalon
Faculdade de Ciências Agrárias, Universidade Federal da Grande Dourados - UFGD, Rodovia Itahum, km 12, Cx. Postal 533, 79804-970, Dourados, MS, Brasil
(*E-mail: cleber_frs@yahoo.com.br)
RESUMO
Pereskia aculeata Plum. (Ora pro nóbis, Cactaceae) é uma planta de interesse medicinal e de valor alimentício por apresentar elevadores teores de fibras e proteínas. Porém, são incipientes informações referentes aos aspectos fisiológicos na fase de crescimento inicial de mudas. Assim, objetivou-se com este trabalho conhecer o metabolismo fotossintético de mudas de P. aculeata propagadas por estaquia sob contrastes de irradiância. O experimento foi desenvolvido sob duas disponibilidades luminosas (Pleno sol e sombreamento de 50%), avaliando-se os tipos de estaca apical, mediana e basal. O arranjo experimental foi em esquema fatorial 2 x 3, no delineamento blocos casualizados, com quatro repetições. Foram quantificados os pigmentos fotossintéticos, emissão da fluorescência da clorofila a e trocas gasosas. Constatou-se que os maiores teores de clorofilas e eficiências fotoquímicas ocorreram nas mudas de estacas apicais cultivadas sob 50% de sombreamento (baixa irradiância). As maiores taxas de assimilação de CO2, eficiência da Rubisco e de uso da água foram constatadas quando utilizadas estacas medianas e basais sob pleno sol (alta irradiância). A propagação por estacas medianas mantidas em alta ou baixa irradiância e basais em alta irradiância propiciaram maior capacidade fotossintética das mudas e podem ser indicadas para a propagação dessa espécie.
Palavras-chave: Ora pro nóbis, compensação luminosa, irradiâncias, planta medicinal.
ABSTRACT
Pereskia aculeata Plum. (Ora pro nóbis, Cactaceae) is a plant of medicinal interest and alimentary value to present high contents of fibers and proteins. However, information regarding the physiological aspects in the initial growth stage of seedlings is incipient. Thus, the aim of this work was to know the photosynthetic metabolism of P. aculeata seedlings propagated by cuttings under irradiance contrasts. The experiment was conducted under two light conditions (full sun and 50% shading), evaluating the types of apical, median and basal cuttings. The experimental arrangement was in a 2 x 3 factorial scheme, in the randomized complete block design, with four replications. Photosynthetic pigments, chlorophyll-a fluorescence emission and gas exchange were quantified. Results showed that the highest levels of chlorophylls and photochemical efficiencies were observed in the apical cuttings cultivated under 50% shading (low irradiance). The highest rates of CO2 assimilation, Rubisco efficiency and water use were observed when using median and basal cuttings under full sun (high irradiance). The propagation by median cuttings maintained in high or low irradiance, and basal cuttings under high irradiance provided a greater photosynthetic capacity of the seedlings, and may be indicated for the propagation of this species.
Keywords: Ora pro nóbis, light compensation, irradiance, medicinal plant.
INTRODUÇÃO
Pereskia aculeata Plum. (Ora pro nóbis, Cactaceae) é uma espécie de interesse alimentício e medicinal, pertencente ao grupo das plantas alimentícias não convencionais. As plantas são distribuídas por diversos estados do Brasil, e vem ganhando espaço nas áreas urbanas e rurais (Souza et al., 2016), tornando-se uma alternativa de renda e fonte para biofortificação.
As folhas de P. aculeata são carnudas e macias, destacando-se na alimentação humana devido ao elevado teor de fibras e proteínas (Almeida & Corrêa, 2012; Kinupp & Lorenzi, 2014) e por ser rica em minerais (ferro, cálcio, zinco e magnésio) (Souza et al., 2015). Além disso, as folhas apresentam-se como fonte de compostos antioxidantes (Souza et al., 2014), fazendo-se necessário seu cultivo ex situ.
A aquisição e padronização de sementes da espécie é difícil, o que inviabiliza a propagação sexuada. Assim, é imprescindível o estudo da propagação vegetativa por estaquia, considerando que é uma técnica de fácil execução, baixo custo e que reduz o período de juvenilidade (Gomes et al., 2015), propiciando mudas vigorosas.
Todavia, salienta-se que são incipientes as informações quanto ao tipo de estaca a ser utilizado, fazendo-se necessários estudos nesta área. Existe grande variação de eficiência de propagação quanto ao tipo de estaca que está associada ao grau de lignificação e reservas presentes nas diferentes partes das estacas caulinares (Santos et al., 2016), podendo afetar a capacidade fotossintética das mudas provenientes das diferentes porções do ramo.
O padrão de qualidade das mudas está interligado à capacidade fotossintética da espécie em relação às condições de cultivo devido ao incremento de fotoassimilados. Dentre os fatores ambientais que podem influenciar as trocas gasosas, pode-se citar a irradiância. O conhecimento da exigência luminosa das plantas é necessário (Lenhard et al., 2013), pois a intensidade pode promover alterações no metabolismo fotossintético e, consequentemente, no desenvolvimento da planta (Yan et al., 2013), que podem apresentar mecanismos de adaptação aos ambientes contrastantes (Veloso et al., 2017).
Portanto, estudos ecofisiológicos auxiliam na compreensão da flexibilidade das respostas bem como na capacidade de aclimatação de espécies vegetais às variantes ambientais (Silva et al., 2011), almejando o aproveitamento da luz, favorecendo a atividade fotossintética e pegamento das mudas.
Entretanto, para a Pereskia aculeata, são incipientes os trabalhos quanto à associação do tipo de estaca e irradiâncias nos aspectos fisiológicos. Desta forma, objetivou-se com este trabalho conhecer as respostas do metabolismo fotossintético em mudas de P. aculeata propagadas por estaquia sob diferentes disponibilidades luminosas.
MATERIAL E MÉTODOS
Desenvolveu-se o experimento no período de maio à agosto de 2017, em ambientes protegidos no Horto de Plantas Medicinais – HPM (22º11'43.7"S e 54º56'08.5"W, 452 m), da Universidade Federal da Grande Dourados, Dourados – MS, Brasil.
As características do metabolismo fotossintético das plantas de P. aculeata foram avaliadas em mudas propagadas por estacas de diferentes posições no ramo, sendo elas do tipo apical, mediana e basal. As mudas foram produzidas sob duas disponibilidades luminosas: pleno sol e sombreamento (cobertura plástica + tela de polipropileno preto de 50% de retenção do fluxo de radiação solar), cujas características são encontradas no Quadro 1. Os dados de déficit de pressão de vapor de água foram calculados considerando a temperatura e humidade relativa de cada ambiente, segundo proposta de Sandler & Evans (1989).
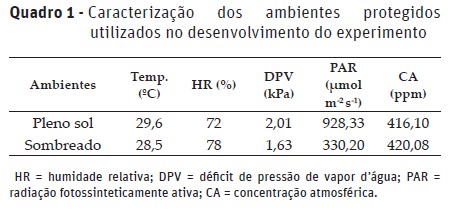
O arranjo experimental foi em esquema fatorial 3 x 2, no delineamento experimental blocos casualizados, com quatro repetições. A unidade experimental constituiu-se de seis recipientes, com uma estaca cada.
A espécie foi identificada, e uma exsicata está depositada no herbário DDMS, sob nº 5226. As estacas foram coletadas no período matutino de plantas matrizes do HPM da UFGD. As estacas foram padronizadas com comprimento de 20 cm, diâmetro médio de 2,27; 3,45 e 4,51 mm e 6, 2, e 2 folhas, para estacas apicais, medianas e basais, respectivamente. A base das estacas foi imersa em recipiente com água para evitar a desidratação do tecido vegetal e/ou oxidação do nó basal.
O substrato base foi composto de mistura de Latossolo Vermelho distroférrico + Tropstrato® (1:1, v/v), apresentando os seguintes atributos químicos após a mistura (Silva, 2009): pH CaCl2 = 5,94; P = 42,65 mg dm3; K = 1,99 cmolc dm3; Ca = 13,28 cmolc dm3; Mg = 19,00 cmolc dm3; H + Al = 2,37 cmolc dm3; V (%) = 62,88, e os atributos físicos: porosidade total = 61,0% e densidade real = 0,94 g cm3.
Realizou-se o enterrio de 1/3 das estacas em sacos de polietileno de 500 mL preenchidos com o substrato base. Os tratos culturais estabelecidos durante o período experimental, constituíram-se de irrigações diárias visando manter a capacidade de campo do substrato base em 70%, e arranquio das plantas espontâneas quando necessário; não ocorreu ataque de pragas e incidência de doenças.
Aos 80 dias após o enterrio (DAE) a estaquia, quando a porcentagem de sobrevivência das estacas estabilizou-se, nas folhas totalmente expandidas localizadas no terço médio das brotações, foram quantificadas as trocas gasosas das mudas, no período da manhã (08h às 10h), utilizando-se de um sistema portátil de analisador de gás por infravermelho (IRGA). Os parâmetros avaliados foram a taxa de assimilação de CO2 (A), concentração interna de CO2 (Ci), condutância estomática (gs) e transpiração (E). A partir dos dados obtidos foram calculados a eficiência do uso da água – (EUA = A/E), de carboxilação da Rubisco – (EiC = A/Ci) e razão Ci/Ca (concentração de CO2 atmosférico/CO2 intercelular).
Para a determinação da emissão da fluorescência da clorofila a, as folhas utilizadas para quantificação das trocas gasosas foram submetidas à condição de escuro por 30 minutos, utilizando-se clipes foliares. Após esse período, sob flash de 1.500 µmol m-2 s-1, com fluorômetro portátil modelo OS 30P, foram mensuradas as fluorescências inicial, variável e máxima da clorofila a. Calcularam-se a eficiência quântica fotoquímica potencial do fotossistema II (FS II) (Fv/Fm), da conversão de energia absorvida (Fv/F0), rendimento máximo não fotoquímico (F0/Fv) e taxa de transporte de elétrons (ETR) (Baker, 2008).
A quantificação dos pigmentos fotossintéticos foi realizada aos 81 DAE, coletando-se limbos foliares totalmente expandidos das brotações por planta em função de cada tipo de estaca e ambiente luminoso, considerando-se as repetições; logo após, foi macerado 1 g de folha em 8 mL de acetona (80%), e utilizando-se microcentrífuga à velocidade de 1.500 rpm, durante 10 minutos, obteve-se as soluções. A leitura da absorbância foi realizada nos comprimentos de onda de 470, 645 e 663 nm, utilizando-se espectrofotômetro e os teores clorofilas a, b, total e carotenoides foram calculados segundo Lichtenthaler (1987).
Os dados foram submetidos à análise de variância, e quando significativos pelo teste F, as médias foram comparadas pelo teste t de Student, para disponibilidades luminosas, e Tukey para tipos de estacas (p≤0,05), utilizando-se o software SISVAR.
RESULTADOS E DISCUSSÃO
A concentração interna de CO2, transpiração e condutância estomática foi influenciada apenas pelo tipo de estaca, independentemente do ambiente luminoso (Quadro 2). A maior Ci e razão Ci/Ca ocorreram nas mudas provenientes de estacas apicais, as quais na fase inicial de implantação do experimento, já apresentavam maiores quantidades de folhas, o que favoreceu maior entrada de CO2.
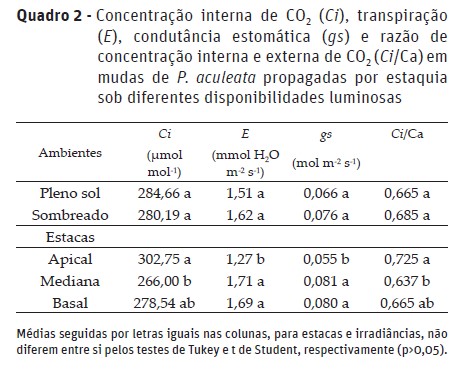
A menor E e gs ocorreu nas mudas provenientes de estacas apicais possivelmente devido a menor turgescência hídrica, comparada às demais estacas em detrimento aos maiores diâmetros das estacas, ou seja, maior quantidade de reservas possivelmente. Esta observação pode ser justificada no facto de as estacas de ponteiro terem apresentado menor grau de lignificação e reservas, desidratando-se facilmente (Rosa et al., 2017). Assim, as folhas da maioria das plantas fecham os estômatos para evitar perda excessiva de água por transpiração (Bertolli et al., 2015); redirecionando a água para manutenção da turgescência da estaca. Com a desidratação das células foliares o aparelho estomático tende a reduzir a condutância, o que minimiza a perda de água por transpiração, mas em consequência, pode reduzir também a entrada de CO2.
A menor eficiência de carboxilação da Rubisco leva à menor assimilação de CO2 representada pela elevação do Carbono interno (Ci) (Quadro 2), reduzindo a capacidade fotossintética (Quadro 3) das mudas de P. aculeata. Assim, observa-se que as estacas medianas apresentam maior eficiência de carboxilação (EiC) e capacidade fotossintética (A) tanto no cultivo a pleno sol quanto sob sombreamento, com valores que não variaram significativamente das estacas basais nos dois regimes de luz. Ressalta-se que maior capacidade fotossintética é uma característica desejável uma vez que a maior produção de fotoassimilados garante maior crescimento e desenvolvimento das mudas.
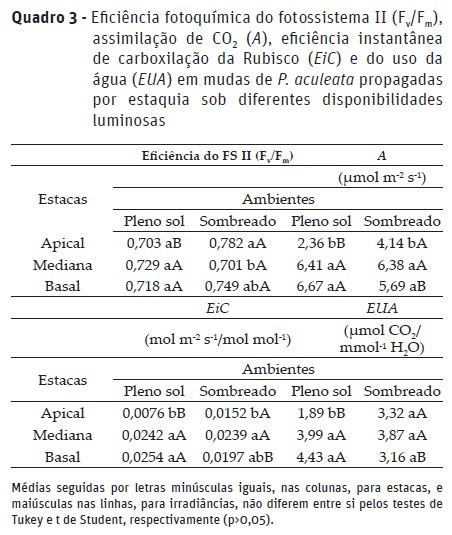
Em plantas de Solanum melongena L. (berinjela), estudando-se lâminas de irrigação, também foi observado que a menor disponibilidade hídrica para as plantas (33% da evapotranspiração), reduziu a condutância estomática e a eficiência de carboxilação (Silva et al., 2015). De maneira semelhante, em mudas de Campomanesia adamantium O. Berg. (Guavira), constatou-se que a menor condutância estomática e redução da eficiência da Rubisco limitaram a capacidade fotossintética (Junglos et al., 2016).
A eficiência do uso da água foi menor nas folhas das mudas provenientes de estacas apicais expostas a alta irradiância (Quadro 3), o que pode ser atribuído à maior desidratação do tecido vegetal em função do menor diâmetro da estaca, isto é, de material de reserva para manutenção da turgescência.
Os parâmetros da fluorescência da clorofila a e eficiência fotoquímica, exceto a fluorescência inicial, foram influenciadas pela interação entre os fatores em estudo. A fluorescência máxima (Fm) da clorofila a foi menor nas mudas de estacas apicais sob alta irradiância, quando comparado as sob ambiente sombreado utilizando-se esse mesmo tipo de estaca. Esse resultado deve-se ao excesso de energia luminosa, ou seja, atingindo o ponto de saturação (Quadro 4), fazendo com que as folhas sob condições de pleno sol apresentam maior quenching não fotoquímico (Casierra-Posada & Ávila-León, 2015) e redução do teor de clorofila (Naschitz et al., 2015), o que pode comprometer as atividades no fotossistema II, tal como observado.
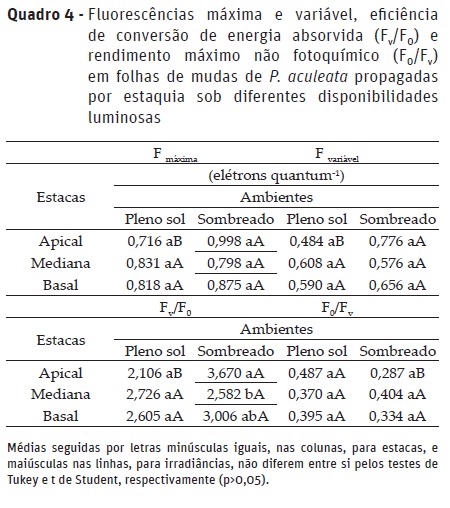
A redução da Fm aponta debilidade no FS II, pois essa característica representa a intensidade da fluorescência quando toda a quinona é reduzida e os centros de reação atingem a capacidade máxima das reações fotoquímicas (Suassuna et al., 2010). No que concerne à alta irradiância, esse resultado pode ser explicado em função de que a planta apresenta um limite de capacidade de absorção de energia luminosa (Gonçalves et al., 2012), isto é, ocorre saturação da maquinaria fotossintética, causando fotoinibição dinâmica.
Quanto à fluorescência variável, eficiência efetiva de conversão de energia absorvida (Fv/F0) e quântica potencial fotoquímica do FS II (Fv/Fm), verificou-se menores valores nas mudas de estacas apicais sob alta irradiância (Quadro 5), diferindo-se estatisticamente das mudas sombreadas, utilizando-se o mesmo tipo de estaca. Os maiores valores em ambiente sombreado devem-se a maior quantidade de clorofilas, isto é, ocorre transporte de forma sincronizada e eficiente, considerando-se como mecanismo de estratégia para evitar valores muito baixos de fotossíntese diária por área foliar (Díez et al., 2017).
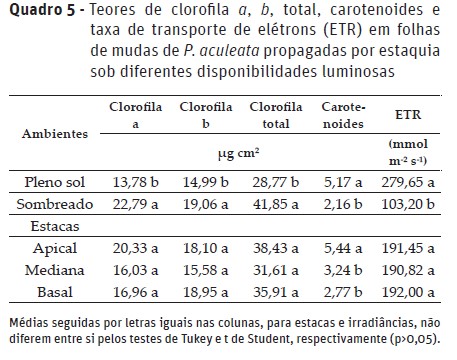
Algumas espécies vegetais realizam adaptações visando otimizar o aproveitamento da luz solar. Essas adaptações sob diferentes condições de irradiâncias estão relacionadas com a eficácia e a rapidez com que os padrões de alocação de biomassa e comportamento fisiológico são ajustados (Mota et al., 2012).
Os teores de pigmentos fotossintéticos foram influenciados apenas pelos contrastes de irradiância. Independente do tipo de estaca constatou-se maiores teores de clorofila a, b e total nas mudas sombreadas (Quadro 5), e os carotenoides e a taxa de transporte de elétrons apresentaram-se em maior concentração nas folhas das plantas cultivadas a pleno sol.
Os maiores teores de clorofilas nas folhas das mudas sombreadas é uma estratégia da planta a essa condição limitante, isto é, de menor radiação fotossinteticamente ativa incidente (Quadro 1). Isso porque, geralmente, plantas cultivadas sob baixa irradiância tendem a aumentar a quantidade de clorofilas como mecanismo de compensação à condição de menor energia no aparato fotossintético (Dutra et al., 2012), tal como observado nas mudas de P. aculeata.
A maior quantidade de clorofila b nos cloroplastos das mudas cultivadas sob baixa irradiância é desejável, tendo em vista que esse pigmento é considerado acessório, isto é, responsável pela captação da energia luminosa, além de ser convertido em clorofila a por meio da enzima clorofila a oxigenase (Xu et al., 2001; Taiz & Zeiger, 2013), fazendo com que haja sincronismo entre os aceptores na cadeia de transporte de elétrons dos fotossistemas. Além disso, a clorofila b realiza a captação da luz em outros comprimentos de onda, transferindo a energia para a clorofila a, favorecendo aproveitamento de luz mais eficiente em condições ambientais de baixa intensidade luminosa (Lima et al., 2011).
Resultados similares quanto à irradiância foram observados por Nery et al. (2016), em plantas jovens de Calophyllus brasiliens Cambess. (Guanandi) cultivadas sob níveis de irradiância (0, 30, 50 e 70%) que apresentaram menores teores de clorofila b nas folhas sob alta irradiância. Quanto aos carotenoides, pode-se destacar que os maiores valores nas mudas sob alta irradiância, devem-se à sua ação como agente mitigador da foto-oxidação das clorofilas (Wondracek et al., 2011).
A partir desses resultados, observou-se que mudas de P. aculeata apresentaram respostas diferenciadas quanto ao metabolismo fotossintético em função do tipo de estaca e irradiância incidente no ambiente de cultivo. Assim, as informações obtidas são relevantes no momento de coleta do material a ser propagado e aclimatização na fase inicial, ou seja, de obtenção de mudas, visando seu cultivo ex situ.
CONCLUSÃO
A propagação por estacas medianas e basais sob alta ou baixa irradiância propiciaram maior capacidade fotossintética das mudas e podem ser indicadas para a propagação vegetativa da Pereskia aculeata Plum.
CONFLITO DE INTERESSES
Não existe conflito de interesses.
REFERÊNCIAS BIBLIOGRÁFICAS
Almeida, M.E.F. & Corrêa, A. D. (2012) - Utilização de cactáceas do gênero Pereskia na alimentação humana em um município de Minas Gerais. Ciência Rural, vol. 42, n. 4, p. 751-756. https://doi.org/10.1590/S0103-84782012000400029 [ Links ]
Baker, B. (2008) - Chlorophyll Fluorescence: A Probe of Photosynthesis in Vivo. Annual Review of Plant Biology, vol. 59, p. 89-113. https://doi.org/10.1146/annurev.arplant.59.032607.092759 [ Links ]
Bertolli, S.C.; Souza, J. & Souza, G.M. (2015) - Caracterização fotossintética da espécie isohídrica pata-de-elefante em condições de deficiência hídrica. Revista Caatinga, vol. 28, n. 3, p. 196–205. https://doi.org/10.1590/1983-21252015v28n322rc
Casierra-Posada, F. & Ávila-León, O.F. (2015) – Shade tolerance of marigold plants (Calendula officinalis). Revista U.D.C.A. Actualidad & Divulgación Científica, vol. 18, n. 1, p. 119-126.
Díez, M.C.; Moreno, F. & Gantiva, E. (2017) – Effects of light intensity on the morphology and CAM photosynthesis of Vanilla planifolia Andrews. Revista Facultad Nacional de Agronomía, vol. 70, n. 1, p. 8023-8033. https://doi.org/10.15446/rfna.v70n1.61736
Dutra, T.R.; Massad, M.D. & Santa, R.C. (2012) - Parâmetros fisiológicos de mudas de copaíba sob diferentes substratos e condições de sombreamento. Ciência Rural, vol. 42, n. 7, p. 1212-1218. https://doi.org/10.1590/S0103-84782012005000048 [ Links ]
Gomes, J.A.O.; Teixeira, D.A.; Marques, A.P.S. & Bonfim, F.P.G. (2015) - Diferentes substratos por estaquia de assa peixe (Vernonia polyanthes Less.). Revista Brasileira de Plantas Medicinais, vol. 17, n. 4, p. 1177-1181. https://doi.org/10.1590/1983-084x/15_008 [ Links ]
Gonçalves, J.F.C.; Melo, E.G.F.; Silva, C.E.M.; Ferreira, M.J. & Justino, G.C. (2012) - Estratégias no uso da energia luminosa por plantas jovens de Genipas pruceana Steyerm submetidas ao alagamento. Acta Botanica Brasilica, vol. 26, n. 2, p. 391-398. https://doi.org/10.1590/S0102-33062012000200014 [ Links ]
Junglos, F.S.; Junglos, M.S.; Dresch, D.M.; Pereira, N.S.; Kodama, F.M. & Scalon, S.P.Q. (2016) - Recovery of the photosynthetic capacity of Campomanesia adamantium (Myrtaceae) after water deficit. Brazilian Journal of Botany, vol. 39, n. 2, p. 541–546. https://doi.org/10.1007/s40415-016-0275-x
Kinupp, V.F. & Lorenzi, H. (2014) - Plantas Alimentícias Não Convencionais (PANC) no Brasil: Guia de Identificação, Aspectos Nutricionais e Receitas Ilustradas. Instituto Plantarum de Estudos da Flora. São Paulo, Brasil. 768 p. [ Links ]
Lenhard, N.R.; Paiva Neto, V.B.; Scalon, S.P.Q. & Alvarenga, A.A. (2013) - Crescimento de mudas de pau-ferro sob diferentes níveis de sombreamento. Pesquisa Agropecuária Tropical, vol. 43, n. 2, p. 178-186. https://doi.org/10.1590/S1983-40632013000200012 [ Links ]
Lichtenthaler, H.K. (1987) - Chlorophylls and carotenoids: pigment photosynthetic biomembranes. Methods in Enzymology, vol. 148, p. 362-385. https://doi.org/10.1016/0076-6879(87)48036-1 [ Links ]
Lima, M.C.; Amarante, L.; Mariot, M.P. & Serpa, R. (2011) – Crescimento e produção de pigmentos fotossintéticos em Achillea millefolium L. cultivada sob diferentes níveis de sombreamento e doses de nitrogênio. Ciência Rural, vol. 41, n. 1, p. 45-50. https://doi.org/10.1590/S0103-84782011000100008
Mota, L.H.S.; Scalon, S.P.Q. & Heinz, R. (2012) - Sombreamento na emergência de plântulas e no crescimento inicial de Dipteryx alata Vog. Ciência Florestal, vol. 22, n. 3, p. 423-431. https://doi.org/10.5902/198050986611 [ Links ]
Naschitz, S.; Naor, A.; Sax, Y.; Shahak, Y. & Rabinowitch, H.D. (2015) - Photo-oxidative sunscald of apple: effects of temperature and light on fruit peel photoinhibition, bleaching and short-term tolerance acquisition. Scientia Horticulturae, vol. 197, p. 5-16. https://doi.org/10.1016/j.scienta.2015.11.003 [ Links ]
Nery, F.C.; Prudente, D.O.; Alvarenga, A.A.; Paiva, R. & Nery, M.C. (2016) - Desenvolvimento de mudas de guanandi (Calophyllum brasiliens Cambess.) sob diferentes condições de sombreamento. Revista Brasileira de Biociências, vol. 14, n. 3, p. 187-192. [ Links ]
Rosa, G.G.; Zanandrea, I.; Mayer, N.A. & Bianchi, V.J. (2017) - Propagação de porta-enxerto de Prunus spp. por estaquia: efeito do genótipo, do estádio de desenvolvimento do ramo e tipo de estaca. Revista Ceres, vol. 64, n. 1, p. 090-097. https://doi.org/10.1590/0034-737x201764010013 [ Links ]
Sadler, E.J. & Evans, D.E. (1989) - Vapor pressure deficit calculations and their effect on the equation combination equation. Agricultural and Forest Meteorology, vol. 49, n. 1, p. 55-80. https://doi.org/10.1016/0168-1923(89)90062-2 [ Links ]
Santos, R.G.; Sousa, I.M.; Albuquerque, C.C. & Silva, K.M.B. (2016) - Tipo de estaca e substrato na propagação vegetativa de Lippia gracilis Schauer. Arquivos do Instituto Biológico, vol. 83, n. e0012014, p. 1-4. https://doi.org/10.1590/1808-1657000012014 [ Links ]
Silva, F.C. (2009) - Manual de análises químicas do solo, plantas e fertilizantes. 2. ed. rev. Ampliada. Brasília, DF: Embrapa Informação Tecnológica, 627 p. [ Links ]
Silva, F.G.; Dutra, W.F.; Dutra, A.F.; Oliveira, I.M.; Filgueiras, L.M.B. & Melo, A.S. (2015) - Trocas gasosas e fluorescência da clorofila em plantas de berinjela sob lâminas de irrigação. Revista Brasileira de Engenharia Agrícola e Ambiental, vol. 19, n. 10, p. 946–952. https://doi.org/10.1590/1807-1929/agriambi.v19n10p946-952
Silva, P.E.M.; Santiago, E.F.; Silva, E.M.; Súarez, Y.R. & Daloso, D.M. (2011) - Fluorescência da clorofila-a e variação da simetria como ferramentas de investigação de plantas sob estresse. IDESIA, vol. 29, n. 3, p. 45-52. https://doi.org/10.4067/S0718-34292011000300007 [ Links ]
Souza, M.R.M.; Pereira, P.R.G.; Magalhães, I.P.B.; Sediyama, M.A.N.; Vidigal, S.M.; Milagres, C. S.F. & Baracat-Pereira, M.C. (2016) - Mineral, protein and nitrate contents in leaves of Pereskia aculeata subjected to nitrogen fertilization. Pesquisa Agropecuária Tropical, vol. 46, n. 1, p. 43-50. https://doi.org/10.1590/1983-40632016v4637959 [ Links ]
Souza, M.S.S.; Barbalho, S.M.; Guiguer, E.L.; Araújo, A.C.; Bueno, P.C.S.; Farinazzi-Machado, F.M.V.; Lima, L.M.L.; Silva, B.C. & Mendes, C.G. (2015) - Effects of Pereskia aculeata Miller on the biochemical profiles and body composition of wistar rats. Journal of Biosciences and Medicines, vol. 3, n. 7, p. 82-89. https://doi.org/10.4236/jbm.2015.37010 [ Links ]
Souza, R.M.F.; Lira, C.S.; Rodrigues, A.O.; Morais, S.A.L.; Queiroz, C.R.A.A.; Chang, R.; Aquino, F.J.T.; Muñoz, R.A.A. & Oliveira, A. (2014) - Atividade antioxidante de extratos de folhas de Ora pro nóbis (Pereskia aculeata Mill.) usando métodos espectrofotométricos e voltamétricos in vitro. Bioscience Journal, vol. 30, n. 1, p. 448-457. [ Links ]
Suassuna, J.F.; Melo, A.S.; Sousa, M.S.S.; Costa, F.S.; Fernandes, P.D.; Pereira, V.M. & Brito, M.E.B. (2010) - Desenvolvimento e eficiência fotoquímica em mudas de híbrido de maracujazeiro sob lâminas de água. Bioscience Journal, vol. 26, n. 4, p. 566-571. [ Links ]
Taiz, L. & Zeiger, E (2013) – Fisiologia Vegetal. 5ª ed. Porto Alegre: Artmed. 918 p.
Veloso, A.C.R.; Silva, P.S.; Siqueira, W.K.; Duarte, K. L.R.; Gomes, I.L.V.; Santos, T. & Fagundes, M. (2017) - Intraspecific variation in seed size and light intensity affect seed germination and initial seedling growth of a tropical shrub. Acta Botanica Brasilica, vol. 31, n. 4, p. 736-741. https://doi.org/10.1590/0102-33062017abb0032 [ Links ]
Wondracek, D.C.; Faleiro, F.G.; Sano, S.M.; Vieira, R.F. & Agostini-Costa, T.S. (2011) - Composição de carotenoides em passifloras do Cerrado. Revista Brasileira de Fruticultura, vol. 33, n. 4, p. 1222-1228. https://dx.doi.org/10.1590/S0100-29452011000400022 [ Links ]
Xu, H.; Vavilin, D. & Vermaas, W. (2001) - Chlorophyll b can serve as the major pigment in functional photosystem II complexes of cyanobacteria. Proceedings of the National Academy of Sciences of the USA, vol. 98, n. 24, p. 14168–14173. https://dx.doi.org/10.1073%2Fpnas.251530298
Yan, N.; Xu, X.F.; Wang, Z.D.; Huang, J.Z. & Guo, D.P. (2013) - Interactive effects of temperature and light intensity on photosynthesis and antioxidant enzyme activity in Zizania latifolia Turcz. plants. Photosynthetica, vol. 51, n. 1, p. 127-138. https://doi.org/10.1007/s11099-013-0009-2 [ Links ]
AGRADECIMENTOS
À CAPES e CNPq, pela concessão das bolsas, e à FUNDECT, pelo apoio financeiro.
Recebido/received: 2018.06.30
Aceite/accepted: 2019.03.06

