











 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUÇÃO
A Fabaceae Vigna unguiculata (L.) Walp., originária do continente africano, mais comumente conhecida em Portugal como feijão-frade e no Brasil como feijão-caupi, é cultivada em regiões áridas e semiáridas, principalmente, pelas suas sementes secas e destina-se à alimentação humana. As vagens também podem ser colhidas em verde e consumidas como o feijão-verde. Em várias regiões do mundo, as folhas também são utilizadas como forragem ou silagem ou incorporadas no solo como adubo verde (Pereira e Meneses, 2019).
A partir do nordeste da África, por volta do ano 2300 a.C., o feijão-frade foi introduzido no sudoeste da Ásia e a partir desta região chegou à Europa (Araújo e Watt, 1988). Na Europa é cultivado em países de clima mediterrânico, como Portugal, Espanha, Itália e Grécia (Almeida et al., 2012; Pereira e Meneses, 2019). No Brasil constitui um componente alimentar básico das populações rurais e urbanas das regiões Norte e Nordeste . A produção de feijão-caupi no Brasil ocorre especialmente em primeira e segunda safra nas regiões Norte, Nordeste e Centro-Oeste (Silva et al., 2016a).
A produtividade da cultura pode ser negativamente afetada por uma série de estresses bióticos e abióticos, que alteram o crescimento e o desenvolvimento vegetal (Silva et al. 2012). Entre os principais fatores abióticos destaca-se a salinidade, uma vez que o excesso de sais pode comprometer as funções fisiológicas e bioquímicas das plantas, causando estresse osmótico, o que resulta em distúrbios das relações hídricas, alterações na absorção e utilização de nutrientes essenciais além da acumulação de iões tóxicos (Assis Júnior et al., 2007; Calvet et al., 2013).
Segundo Santos et al. (2009), as culturas respondem diferentemente à salinidade podendo ser desde sensíveis a altamente tolerantes ao ambiente salino. De acordo com Ayers e Westcot (1999), o feijão-caupi é considerado uma espécie moderadamente tolerante à salinidade da água de irrigação apresentando salinidade limiar de 3,3 dS m-1. Entretanto, esse valor limiar varia amplamente entre as cultivares, e depende não só do tipo de sal, mas também do tempo de exposição e do seu estádio de desenvolvimento (Deuner et al., 2011).
As culturas agrícolas apresentam sensibilidade diferenciada em relação aos tipos de sais, uma vez que as diferenças químicas existentes entre eles podem originar diferenças nos resultados de germinação e vigor das sementes, mesmo em concentrações salinas similares (Conus et al., 2009). Por isso, estudos com sementes de algumas espécies, como feijão, milho e sorgo, têm sido desenvolvidos com diversas soluções salinas, principalmente, a base de NaCl, CaCl2, KCl e MgCl2, para simular condições de estresse salino (Moterle et al., 2006; Custódio et al., 2009; Coelho et al., 2010, 2014; Silva et al., 2016b). Todavia, no que concerne ao feijão-caupi as informações são escassas, sendo necessário desenvolver investigação neste domínio.
Considerando que o aumento da salinidade dos solos está associado à redução dos seus potenciais a ponto de limitar a germinação de sementes, estudos sobre a germinação e o desenvolvimento inicial das plântulas nessas condições são importantes (Munns e Tester, 2008). Larcher (2006) afirma que a avaliação do processo de germinação pode indicar a sensibilidade das plantas à salinidade em estádios subsequentes do crescimento e do desenvolvimento.
Face ao exposto, os objetivos deste trabalho consistiram em avaliar os efeitos do estresse salino induzido por diferentes condutividades elétricas das soluções de cloreto de sódio (NaCl), cloreto de cálcio (CaCl2) e da combinação de CaCl2 + NaCl sobre a germinação e o crescimento inicial de plântulas de Vigna unguiculata cultivar ‘BR 17 Gurguéia’.
MATERIAL E MÉTODOS
O estudo foi conduzido no Laboratório de Tecnologia de Sementes da Universidade Estadual do Sudoeste da Bahia, Campus de Vitória da Conquista, BA, em outubro de 2014, utilizando sementes de feijão-caupi (Vigna unguiculata), cultivar ‘BR17 Gurguéia’ (safra 2013), cedidas pela Embrapa Meio-Norte.
Para a promoção do estresse salino foram preparadas soluções de cloreto de sódio (NaCl), cloreto de cálcio (CaCl2) e da combinação de CaCl2 + NaCl, na proporção equivalente de 3:2. As soluções salinas utilizadas foram calibradas, em condutivímetro “Digimed” modelo DM-31, para as seguintes condutividades elétricas (CE): 0,0 (água destilada sem adição de sais, compondo a testemunha); 2,5; 5,0; 7,5; 10 e 12,5 dS m-1 (Quadro 1). Essas soluções corresponderam a potenciais osmóticos (Ψos) de aproximadamente 0,0; -0,09; -0,18; -0,27; -0,36, e -0,45 MPa, respetivamente.
Quadro 1 Condutividade elétrica da solução de embebição e respetivas concentrações de NaCl, CaCl2 e da combinação de CaCl2 + NaCl
| Condutividade elétrica | NaCl | CaCl2 | CaCl2 + NaCl (3:2) | |
|---|---|---|---|---|
| (dS m-1) | _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ (g L-1) _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ | |||
| 0,0 | 0,00 | 0,00 | 0,0: 0,0 | |
| 2,5 | 1,37 | 1,92 | 0,9: 0,6 | |
| 5,0 | 2,73 | 3,83 | 1,8: 1,2 | |
| 7,5 | 4,10 | 5,75 | 2,7: 1,8 | |
| 10,0 | 5,46 | 7,66 | 3,6: 2,4 | |
| 12,5 | 6,83 | 9,58 | 4,5: 3,0 | |
A sementeira foi realizada em substrato (papel Germitest®) umedecido com as soluções salinas em quantidade equivalente a 2,5 vezes a sua massa. Em seguida, foram confeccionados rolos, sendo envolvidos em sacos plásticos e colocados em germinador regulado a 25 °C, dispostos no sentido vertical, de maneira que o geotropismo se manifestasse naturalmente facilitando as avaliações das plântulas. Foram utilizadas quatro repetições de 50 sementes para cada tratamento, com as contagens realizadas aos cinco (1ª contagem) e oito dias após a sementeira, determinando-se a porcentagem de plântulas normais e anormais, conforme as Regras para Análise de Sementes (Brasil, 2009).
O índice de velocidade de germinação (IVG) foi determinado mediante contagem diária das plântulas normais, sendo o índice calculado pela fórmula proposta por Maguire (1962).
Após o teste de germinação, realizou-se análise de crescimento de plântulas, determinando-se o comprimento (raiz até a inserção dos cotilédones) das plântulas normais de cada repetição, com o auxílio de uma régua graduada em centímetros, sendo os resultados expressos em centímetros por plântula. Para obtenção da massa seca, as plântulas, provenientes da avaliação do comprimento, foram colocadas em sacos de papel do tipo Kraft® e acondicionadas em estufa com circulação forçada de ar, regulada a 65 °C, onde permaneceram até atingir massa constante. Em seguida, foram pesadas em balança analítica com precisão de 0,001 g, sendo os resultados expressos em gramas por plântula.
O delineamento experimental adotado foi o inteiramente casualizado, em esquema fatorial 3 x 6, sendo três indutores de salinidade (NaCl, CaCl2 e CaCl2 + NaCl) e seis níveis de salinidade (0,0; 2,5; 5,0; 7,5; 10,0 e 12,5 dS m-1), com quatro repetições de 50 sementes para cada tratamento. Os dados foram submetidos à análise de homogeneidade das variâncias (teste de Bartlett) e de normalidade (Lilliefors). Em seguida, foi realizada análise de variância pelo teste F e para a comparação das médias utilizou-se o teste de Tukey, a 5% de probabilidade. Para os efeitos quantitativos foi realizada análise de regressão polinomial, selecionando-se o modelo de maior grau significativo.
RESULTADOS E DISCUSSÃO
Para todas as variáveis avaliadas houve interação significativa entre os fatores sais e os níveis de salinidade (p<0,05), exceto para os dados de plântulas anormais, em que só houve efeito significativo isolado dos níveis de salinidade.
Em relação ao desempenho germinativo das sementes, para os indutores NaCl, CaCl2 e a combinação de CaCl2 + NaCl, verificaram-se os pontos de máxima germinação nas condutividades estimadas correspondentes a 5,4; 4,6 e 4,8 dS m-1, respetivamente, com 97; 99 e 98% de germinação, respetivamente. Entretanto, com o aumento da concentração salina no substrato, observou-se pequeno decréscimo na germinação, sendo atingidos os menores resultados em 12,5 dS m-1 (94; 88 e 91%, respetivamente). Apesar da redução observada, a germinação manteve-se acima de 85%, evidenciando que o potencial germinativo do feijão-caupi é pouco alterado mesmo em condições de estresse salino (Figura 1A).
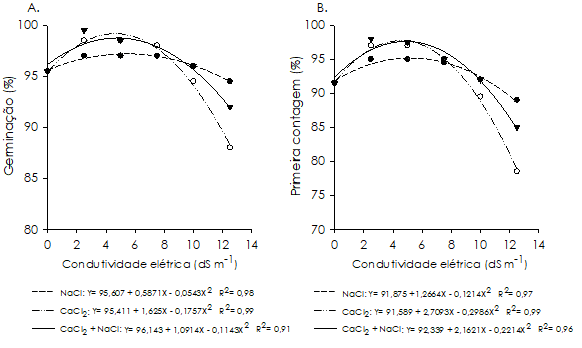
Figura 1 Germinação (A) e primeira contagem da germinação (B) de sementes de Vigna unguiculata cultivar ‘BR 17 Gurguéia’, sob estresse salino induzido por soluções de NaCl, CaCl2 e CaCl2 + NaCl com diferentes condutividades elétricas.
O fato de o processo de germinação do feijão-caupi ser pouco afetado em níveis de salinidade de até 12,5 dS m-1, pode ser um indicativo de tolerância à salinidade da cultivar ‘BR 17 Gurguéia’. Comportamento semelhante foi observado em sementes de sorgo forrageiro (genótipo CSF 18) e de milho (Oliveira e Gomes-Filho, 2009; Silva et al., 2016b). No entanto, para essas espécies os estádios de desenvolvimento subsequentes foram afetados fortemente pelo excesso de sais. Coelho et al. (2010) afirmam que a fase de germinação é menos sensível ao estresse salino quando comparado aos estádios posteriores, o que justificaria as elevadas percentagens de germinação nessas condições.
O efeito dos sais sobre a germinação e o estabelecimento das plântulas pode correlacionar-se tanto aos efeitos de natureza física, uma vez que a salinidade diminui o potencial osmótico da solução do solo, o que faz com que a absorção de água pelas sementes e raízes seja mais difícil, e devido a efeitos tóxicos passíveis de ocorrer por ação direta de iões específicos ou por desordem nutricional (Calvet et al., 2013).
A correlação com esses efeitos (osmóticos e tóxicos) contribui para a compreensão de determinados resultados encontrados neste estudo, como a redução do desempenho germinativo das sementes e aumento da ocorrência de anormalidades na formação das plântulas feijão-caupi em condutividades a partir de 5,0 dS m-1, devido, possivelmente, a redução da disponibilidade hídrica necessária para germinação e o acumulação de iões tóxicos no protoplasma celular.
Nos níveis moderados de salinidade (2,5 e 5,0 dS m-1), a germinação do feijão-caupi pode ter sido beneficiada pelo condicionamento osmótico (priming) ocasionado pela adição de sais na solução de embebição, uma vez que a redução do potencial hídrico do substrato pode proporcionar entrada de água mais lenta e uniforme aos tecidos das sementes, possibilitando maior capacidade de reorganização destes de forma a favorecer o processo germinativo (Dantas et al., 2007; Silva et al., 2016b).
Na ocasião da primeira contagem (Figura 1B), para os tratamentos com NaCl, CaCl2 e a combinação de CaCl2 + NaCl, verificaram-se valores máximos da germinação nas condutividades estimadas equivalentes a 5,2; 4,5 e 4,9 dS m-1, respetivamente, com valores da ordem de 95; 97 e 97%, respetivamente. Entretanto, com o aumento do nível de estresse houve redução da germinação na primeira contagem até os 12,5 dS m-1, em que foram obtidos os menores resultados (88; 78 e 84%, respetivamente).
Os resultados obtidos estão de acordo com os de Deuner et al. (2011) que verificaram redução na germinação de sementes de genótipos de feijão-caupi (cvs. ‘Amendoim’, ‘Baio’, ‘Mosqueado’ e ‘Preto’) na primeira contagem, a partir da condutividade de 5,0 dS m-1 de NaCl, retardando o processo germinativo.
Na avaliação dos sais dentro de cada condutividade elétrica (Figura 1), verificou-se, para a germinação e primeira contagem, diferenças significativas (p<0,05) entre as soluções salinas apenas na condutividade de 12,5 dS m-1, sendo o tratamento com CaCl2 mais severo em relação aos restantes, promovendo decréscimo no potencial germinativo das sementes. Resultados similares foram obtidos por Coelho et al. (2010), ao observarem que os efeitos do CaCl2 foram mais drásticos sobre a germinação de sementes de feijão (Phaseolus vulgaris L.), em relação aos do NaCl e KCl.
Quanto ao índice de velocidade de germinação (Figura 2), o NaCl e o CaCl2 promoveram desempenho semelhante na indução do estresse salino, obtendo-se máxima velocidade de germinação nas condutividades estimadas de 3,2 e 3,4 dS m-1, respetivamente. A partir destas concentrações houve decréscimo na velocidade de germinação até o maior nível de salinidade (12,5 dS m-1), registrando-se redução de 31 e 28%, respetivamente, em relação à testemunha; indicando que em concentrações mais elevadas, o estresse salino induzido por estes sais retarda a germinação das sementes de feijão-caupi.
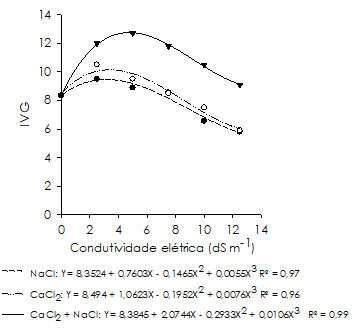
Figura 2 Índice de velocidade de germinação (IVG) de sementes de Vigna unguiculata cultivar ‘BR 17 Gurguéia’, sob estresse salino induzido por soluções de NaCl, CaCl2 e CaCl2 + NaCl com diferentes condutividades elétricas.
A explicação para este comportamento é devido às condições salinas, ocorrendo redução do potencial osmótico do meio, e consequentemente, aumento do tempo de embebição de água pelas sementes, ocasionando o prolongamento do processo germinativo (Khan e Panda, 2008). Atraso na germinação de sementes ocasionado pelo excesso de sais também foi observado por outros autores em diversas culturas de importância económica, como milho, feijão e sorgo (Moterle et al., 2006; Custódio et al., 2009; Oliveira e Gomes-Filho, 2009).
Para a combinação de CaCl2 + NaCl (Figura 2), a maior velocidade de germinação foi registrada na condutividade estimada de 4,8 dS m-1 (12,7). Logo, em concentrações superiores houve decréscimo na velocidade de germinação, atingindo índice de 9,1 em 12,5 dS m-1, que apesar da redução sofrida, foi superior ao obtido na testemunha (8,4). Tal fato demonstra que o estresse salino induzido pela combinação destes sais interfere positivamente na velocidade do processo germinativo do feijão-caupi.
Entretanto, contrariando esses resultados, ao avaliar a emergência e o vigor de genótipos de feijão-caupi sob estresse salino, em condição de casa de vegetação e semeadura em substrato inerte (areia lavada), Almeida et al. (2012) observaram que o aumento da salinidade provocada pela mistura dos sais NaCl, CaCl2 e MgCl2 (7:2:1) (0; 2,5; 5,0 e 7,5 dS m-1) na água de irrigação reduziu o índice de velocidade de emergência das plântulas.
A combinação de CaCl2 + NaCl foi menos drástica para o índice de velocidade de germinação (IVG) em relação às soluções de NaCl e CaCl2, as quais não diferiram entre si, com exceção das condutividades de 2,5 e 10,0 dS m-1, em que o tratamento com NaCl apresentou resultados mais severos para esta variável (Figura 2). Para Tobe et al. (2003), a diminuição do processo de absorção de água e a entrada dos iões em quantidade suficiente para provocarem toxicidade às sementes, especialmente o Na+ e o Cl-, são apontadas como as principais causas da redução da velocidade dos processos fisiológicos e bioquímicos.
Na avaliação de plântulas anormais na condição de estresse salino (Figura 3), constatou-se, a partir da condutividade de 5,0 dS m-1, aumento progressivo na incidência de anormalidades, alcançando maior percentagem em 12,5 dS m-1 (6%); demonstrando que em concentrações mais elevadas, o estresse salino promove aumento da ocorrência de plântulas anormais de feijão-caupi.
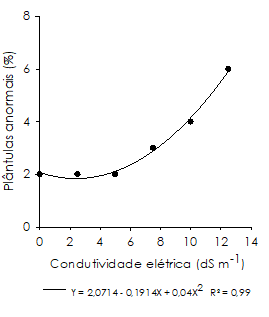
Figura 3 Plântulas anormais de Vigna unguiculata cultivar ‘BR 17 Gurguéia’, oriundas de sementes submetidas ao estresse salino.
Para as sementes que germinam sob condições de salinidade, os efeitos negativos são acumulativos desde a sinalização e ativação do mecanismo de germinação, aumentando a probabilidade de ocorrerem degenerações durante o desenvolvimento da plântula (Assis Júnior et al., 2007).
Na análise de crescimento de plântulas, observou-se que, na indução por NaCl e na combinação de CaCl2 + NaCl, os valores máximos de comprimento de plântulas foram alcançados nas condutividades estimadas correspondentes a 3,7 e 4,2 dS m-1, respetivamente; revelando que, provavelmente, o feijão-caupi apresenta maior crescimento inicial em condições moderadas de salinidade (Figura 4A), efeito semelhante observado em sorgo forrageiro por Coelho et al. (2014).
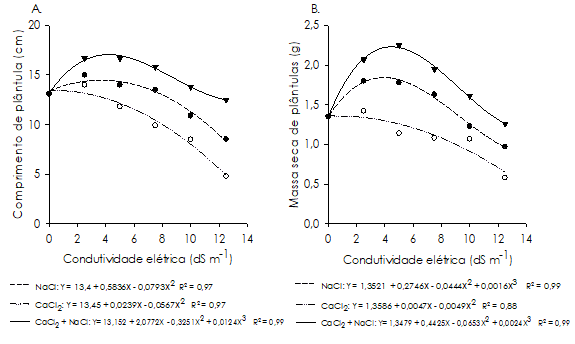
Figura 4 Comprimento (A) e massa seca (B) de plântulas de Vigna unguiculata cultivar ‘BR 17 Gurguéia’, oriundas de sementes submetidas ao estresse salino induzido por soluções de NaCl, CaCl2 e CaCl2 + NaCl com diferentes condutividades elétricas.
No entanto, o aumento do nível de salinidade no substrato ocasionou redução no comprimento de plântulas, atingindo os menores resultados em 12,5 dS m-1, verificando-se decréscimo de 38% na indução por NaCl e 4,5% para a combinação de CaCl2 + NaCl, quando comparados à testemunha (Figura 4A); demonstrando que elevadas concentrações salinas, principalmente de NaCl, comprometem significativamente o comprimento das plântulas de feijão-caupi, com efeitos mais severos que os observados na germinação (Figura 1A).
Esses resultados coincidem com os apresentados por Santos et al. (2009), em que o vigor de plântulas de feijão foi mais afetado que a germinação quando submetido ao estresse salino induzido por soluções de NaNO3, NaCl, NaHCO3, provocando maior redução de comprimento e acumulação de massa seca. Para Silva et al. (2012), as variáveis de crescimento são particularmente sensíveis ao efeito salino; assim, a taxa de crescimento e a produção de biomassa são critérios adequados à avaliação da condição de salinidade e da capacidade da planta na superação do estresse.
Para indução com CaCl2 (Figura 4A), observou-se que o comprimento de plântulas diminui à medida que se aumentou a concentração salina no substrato, com os maiores valores registrados na testemunha (13,45 cm) e os menores na condutividade de 12,5 dS m-1 (4,8 cm). Essa mesma tendência foi observada por Moterle et al. (2006), ao constatarem redução linear no comprimento da parte aérea e raiz primária de plântulas de cultivares de milho-pipoca (Zea mays) à medida que o potencial osmótico da solução de KCl foi diminuído.
Essa redução no comprimento das plântulas deve-se às mudanças na turgescência celular, em função da diminuição da síntese de proteína nas condições de estresse salino (Munns e Tester, 2008). Taiz e Zeiger (2013) relatam que o primeiro efeito mensurável do estresse salino é a diminuição no crescimento, causada pela redução da expansão celular.
Para a massa seca de plântulas (Figura 4B), na indução por NaCl e na combinação de CaCl2 + NaCl, observou-se maior acumulação de massa nos níveis estimados equivalentes a 4,0 e 4,5 dS m-1, respetivamente, apresentando valores de 1,84 e 2,23 g, respetivamente; evidenciando a tolerância do feijão-caupi à salinidade, uma vez que concentrações moderadas destes sais contribuíram para uma maior acumulação de massa seca pelas plântulas em relação à testemunha. No entanto, a partir destas condutividades houve decréscimo na massa seca das plântulas, com os menores resultados verificados na condutividade de 12,5 dS m-1, constatando-se redução de aproximadamente 28 e 4%, respetivamente, em relação à testemunha.
Desempenho semelhante foi observado em plântulas de híbridos de milho (Silva et al. (2016b) e de feijão (Dantas et al., 2007), sendo este fato atribuído a um possível benefício do NaCl em controlar a entrada de água nas sementes e assim prevenindo possíveis danos às membranas celulares.
No tratamento com CaCl2 houve redução na massa seca de plântulas com o aumento do nível de estresse, sendo os maiores valores obtidos na testemunha (1,35 g) e os menores na condutividade de 12,5 dS m-1 (0,65 g) (Figura 4B). Quando em concentrações mais elevadas, o estresse salino provocou redução no crescimento das plântulas. Este fator é atribuído à diminuição na absorção de água, seguido por hidrólise limitada de reservas alimentares a partir de tecidos de armazenamento, bem como devido à translocação deficiente dessas reservas para o eixo embrionário em desenvolvimento (Khan e Panda, 2008).
A indução por CaCl2, a partir da condutividade de 5,0 dS m-1, proporcionou resultados mais severos para o comprimento e massa seca de plântulas, seguido do NaCl e por último da combinação de CaCl2 + NaCl (Figura 4). Uma possível interpretação para este resultado é que o ião Ca2+ pode promover a modulação de processos celulares por meio de proteínas (calmodulinas), as quais, por sua vez, regulam uma cascata de eventos relacionados com as respostas das plantas a estresse biótico e abiótico (Dombrowski e Bergey, 2007), o que poderia ter potencializado a tendência de redução no crescimento das plântulas observado no tratamento com CaCl2. Por outro lado, contrariando os resultados obtidos, Conus et al. (2009) verificaram que a indução por CaCl2 proporcionou maiores resultados de comprimento da parte aérea e raiz primária de plântulas de milho em relação às soluções de KCl e NaCl.
A combinação de CaCl2 + NaCl permitiu maior transferência das reservas da semente para o crescimento das plântulas, proporcionando maiores resultados de comprimento e massa seca de plântulas na indução do estresse salino (Figura 4). Isto deve-se, provavelmente, ao fato de ser o Ca2+ essencial para a seletividade de canais de catiões na plasmalema de células das raízes, tendo, consequentemente, efeito sobre o influxo passivo de Na+ (Aragão et al., 2010).
O Ca+2, além de reduzir a absorção de iões potencialmente tóxicos, como o Na+, favorece a absorção de iões essenciais, como o K+ e o próprio Ca2+ (Aquino et al., 2007). Assim, devido aos antagonismos iónicos em ambientes salinos, a tolerância à salinidade entre genótipos ou mesmo entre espécies é dependente da composição iónica da água de irrigação.
Corroborando com os resultados obtidos, Lacerda et al. (2004) constataram que o aumento na concentração de Ca2+ na solução nutritiva reduziu os teores de Na+ e Cl- nas folhas de plântulas de genótipos de sorgo sob estresse salino. Esta redução deve-se, provavelmente, ao melhor controle na absorção radicular e no transporte de Na+ e Cl- para a parte aérea que ocorre em níveis adequados de Ca2+ no meio de nutrição e tem sido apontada como um dos principais efeitos benéficos do Ca2+ em plantas sob condições de estresse salino (Munns e Tester, 2008).
De maneira geral, a germinação e o crescimento inicial das plântulas foram incrementados em níveis de salinidade entre 2,5 e 5,0 dS m-1, no entanto, em concentrações salinas mais elevadas o desempenho das sementes de feijão-caupi foi afetado negativamente. Além disso, o CaCl2 proporcionou efeitos mais drásticos sobre o crescimento das plântulas em relação ao NaCl, enquanto que a combinação de CaCl2 + NaCl atenuou o estresse salino, originando plântulas mais vigorosas. Os resultados revelam a importância da avaliação dos níveis de salinidade do substrato, considerados críticos para a germinação das sementes e estabelecimento das plântulas de feijão-caupi, além da necessidade de se determinar as potencialidades dos diferentes sais na indução do estresse salino.
Vale ressaltar que são necessários novos estudos que avaliem a resposta de outras cultivares em condições de estresse salino, a fim de se obter informações acerca do grau de tolerância de genótipos de feijão-caupi à salinidade nos estágios iniciais de desenvolvimento, pois de acordo com Santos et al. (2009), a tolerância à salinidade varia entre espécies, entre variedades/genótipos de uma mesma espécie e até mesmo entre estádios fenológicos de um mesmo genótipo.
CONCLUSÕES
Níveis de salinidade entre 2,5 e 5,0 dS m-1 favorecem a germinação e o crescimento inicial de plântulas, sugerindo tolerância moderada do feijão-caupi, cultivar ‘BR 17 Gurguéia’, à salinidade.
O desempenho germinativo e o crescimento de plântulas de feijão-caupi foram afetados negativamente pelo estresse salino em níveis acima de 5,0 dS m-1, independente do sal utilizado.
O CaCl2 promove efeitos mais severos sobre o crescimento das plântulas de feijão-caupi quando comparado ao NaCl, enquanto que a combinação de CaCl2 + NaCl atenua o estresse salino, originando plântulas mais vigorosas.

