











 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUÇÃO
A mesorregião sudeste do Estado do Pará, Brasil, destaca-se por ter um número considerável de assentamentos rurais da reforma agrária, entre eles encontra-se o Projeto de Assentamento 26 de Março, no município de Marabá, que teve sua base sócio econômica marcada por ciclos de exploração dos recursos naturais, iniciados com a extração de Castanheira (Bertholletia excelsa) e, posteriormente, a derrubada da floresta para atender a demanda de atividades madeireiras e agropecuárias (Castro e Watrin, 2013; Neves e Schimitz, 2018).
Atualmente, o Projeto de Assentamento26 de Março se destaca principalmente pelo cultivo de mandioca (Manihot esculenta) e banana (Musa sp.), advindos dos estabelecimentos agrícolas familiares, com dinâmica de preparo de áreas através da prática do corte e da queima e com predominância de sistemas de cultivos que não adotaram a calagem para correção da acidez do solo e não realizaram adubações de fontes minerais e/ou orgânicas.
Os solos paraenses apresentam boas condições físicas como textura, estrutura e porosidade. Mas, quanto a condições químicas, apresentam baixa fertilidade, por serem muito intemperizados (Gama et al., 2020). Sabe-se que uma das limitações para a sustentabilidade de áreas de plantio é a constituição química do solo, sendo o nitrogênio (N) considerado um dos nutrientes mais limitantes para o crescimento vegetal. Por outro lado, uma das formas de disponibilizar o N, em áreas de cultivo, é a utilização de espécies vegetais capazes de efetuar simbioses com bactérias que nodulam leguminosas (BNL) e fixam nitrogênio atmosférico. O processo ocorre no interior dos nódulos que são formados a partir de sinalizações moleculares entre a planta e a bactéria, podendo ser influenciado por fatores edafoclimáticos que podem originar uma redução na eficiência das estirpes (Dabessa et al., 2018).
Estudos que permitam uma avaliação, mesmo que parcial, da ocorrência e da funcionalidade das populações de BNL podem fornecer informações úteis sobre as condições edáficas, além de serem importantes sob dois importantes aspectos. O primeiro é que o solo é um reservatório importante de recursos genéticos para a seleção de estirpes eficientes, componentes das populações nativas (Soares et al., 2006). O segundo refere-se ao conhecimento das populações nativas edáficas e suas alterações com as mudanças no manejo e nos atributos do solo, que podem constituir um indicador das intervenções antrópicas sobre a estrutura populacional do simbionte alvo em estudo (Melloni et al., 2006). Para a análise da ocorrência e da potencialidade destas populações, em estudos exploratórios, a planta armadilha constitui mais uma variável a ser considerada, pois influencia na captura das bactérias nativas, já que sua capacidade de estabelecer simbiose é restrita a determinadas espécies de bactérias (Moreira e Siqueira, 2006). Dentre as leguminosas consideradas mais promíscuas encontra-se o feijão caupi (Melloni et al., 2006; Almeida et al., 2018).
Desta forma, o isolamento e a caracterização de rizóbios noduladores de feijão caupi em solos de áreas com diferentes manejos e usos podem possibilitar a identificação e futura seleção de estirpes adaptadas a condições edafoclimáticas presentes. Importante ressaltar que a região amazônica tem se destacado por apresentar estirpes de bactérias com alto potencial de utilização na promoção do crescimento de diversas culturas não só por meio do processo da fixação biologia de N2 (Almeida et al., 2010) mas, também, por outros, tais como: inibição do crescimento de fungos fitopatogênicos, fixação biológica de nitrogênio de vida livre, solubilização de fosfato de cálcio e produção de ácidos indolacético (Oliveira-Longatti et al., 2014) e indução de tolerância a acidez do solo e outros estresses abióticos (Soares et al., 2014). Portanto, estudos utilizando bactérias isoladas da região amazônica, além de contribuírem para o maior conhecimento da diversidade deste importante grupo de microrganismos nesta região, ainda podem ser promissores para o isolamento, por promoverem o crescimento vegetal de diversas culturas de importância social.
Apesar da relevância de pesquisas sobre o conhecimento de características peculiares de isolados bacterianos nodulíferos nativos, são escassos os trabalhos em ambientes como o Sudeste paraense. Neste contexto, o objetivo deste trabalho foi isolar e caracterizar estirpes de bactérias que nodulam feijão caupi oriundos de diferentes áreas no Projeto de Assentamento 26 de Março, em Marabá, Estado do Pará, Brasil.
MATERIAL E MÉTODOS
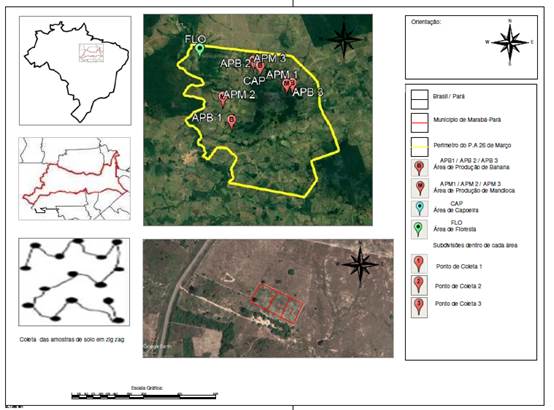
O experimento foi conduzindo em casa de vegetação, no Instituto Federal de Educação, Ciência e Tecnologia do Pará, Campus Marabá Rural, com coordenadas geográficas (-5° 34’ 11,972’’ S; -49° 6’ 2,041” W), em Marabá, Pará, entre março e abril de 2020. As amostras de solos utilizadas no experimento foram oriundas de lotes produtores de mandioca, banana e áreas de capoeira e de floresta localizadas no Projeto de Assentamento 26 de Março, região sudeste do Pará, no município já mencionado, sem a utilização de inoculantes comerciais. A identificação das áreas de coleta e características do manejo encontram-se no Quadro 1.
Quadro 1 Identificação das áreas de coleta de solo em áreas de produção de banana, de mandioca e em áreas de referência no Projeto de Assentamento PA 26 de Março, no ano de 2018/2019
| Área de produção de Banana Lote 1 - APB 1 |
|---|
| Área de produção de 3,32 ha, com a retirada de 480 cachos/mês; correção do solo com calcário. A dinâmica de preparo das áreas ocorre através da prática do corte e da queima e ausência de sistema de preparo de área mecanizado. Não é adotada a calagem para correção da acidez do solo nem adubações de fontes minerais e/ou orgânicas. É realizada a limpeza da área e a poda apenas uma vez ao ano, plantio solteiro; aplicação de herbicida Glifosato; plantas com idade de dois a três anos. Presença das cultivares Brs Conquista e Banana da terra. |
| Área de produção de Banana 2 - APB 2 |
| Área de produção de 2,42 ha com a retirada de 120 cachos/mês; não realiza correção do solo; a dinâmica de preparo das áreas ocorre através da prática do corte e de queima e ausência de sistema de preparo de área mecanizado. Não é adotada a calagem para correção da acidez do solo nem adubações de fontes minerais e/ou orgânicas. É realizada a limpeza da área e a poda de acordo com a necessidade da cultura plantio solteiro; plantas com idade de quatro anos. Presença das cultivares Brs Conquista, Couruda e Prata. |
| Área de produção de Banana 3 - APB 3 |
| Área de produção de 9,68 ha com a retirada de 10 cachos/mês; não se realiza correção e adubação do solo; a dinâmica de preparo das áreas ocorre através da prática do corte e da queima e ausência de sistema de preparo de área mecanizado. Não é adotada a calagem para correção da acidez do solo nem adubações de fontes minerais e/ou orgânicas. Não é realizada poda, plantio solteiro; plantas com idade de três a quatro anos. Presença das cultivares Brs Conquista, Banana-maçã e Pacovan. |
| Área de produção de Mandioca 1 - APM 1 |
| A área total de produção apresenta 4,84 ha, destinada à fabricação de farinha, cujo rendimento é 4.000 kg farinha ha-1, comercializado por meio de atravessadores. O sistema é realizado em consócio com Zea mays. A dinâmica de preparo das áreas ocorre através da prática do corte e da queima e ausência de sistema de preparo de área mecanizado. Não é adotada a calagem para correção da acidez do solo nem adubações de fontes minerais e/ou orgânicas. Presença da variedade Amazonas. |
| Área de produção de Mandioca Lote 2 - APM 2 |
| A área total de produção apresenta 2,42 ha, destinada à fabricação de farinha, cujo rendimento é 3.000 kg ha-1, comercializado por meio de atravessadores. O sistema de cultivo da mandioca é realizado em consorciação com Zea mays e Euterpe oleracea. A dinâmica de preparo das áreas ocorre através da prática do corte e da queima e ausência de sistema de preparo de área mecanizado. Não é adotada a calagem para correção da acidez do solo nem adubações de fontes minerais e/ou orgânicas. Presença da variedade Amazonas. |
| Área de produção de Mandioca Lote 3 - APM 3 |
| A área total de produção apresenta 1,81 ha, destinada à fabricação de farinha, cujo rendimento é 2.400 kg ha-1, comercializado por meio de atravessadores. Adota o sistema de cultivo em monocultivo. A dinâmica de preparo das áreas ocorre através da prática do corte e da queima e ausência de sistema de preparo de área mecanizado. Não é adotada a calagem para correção da acidez do solo nem adubações de fontes minerais e/ou orgânicas. Presença da variedade Amazonas. |
| Área de Capoeira - CAP |
| Cultivo anterior de pastagem. Área com quatro anos em regeneração natural. |
| Área de Floresta - FLO |
| Floresta estabelecida. Área anteriormente utilizada com pastagem. Desde 1980, encontra-se em processo de regeneração natural. O Manejo Florestal ocorreu com enriquecimento da área com mudas de Cupuaçu (Theobroma grandiflorum), Seringueira (Hevea brasiliensis) e Castanha do Pará (Bertholletia excelsa). |
Cada área foi subdividida em três parcelas sendo destas coletadas 15 amostras simples na forma de zig zag na profundidade de 0-20 cm (Figura 1). Totalizando 45 pontos de coletas por área de estudo. Parte do solo foi utilizada para o experimento em vaso com a capacidade de 0,5 kg cada, com três repetições. Outra parte foi destinada a caracterização física e química das amostras de solo (Quadro 2).
Quadro 2 Granulometria, classificação textural e médias de pH, teores de cálcio (Ca2+), magnésio (Mg2+), potássio (K+), fósforo (p), Zinco (Zn), teores de alumínio (Al), hidrogênio + alumínio (H + Al), saturação por bases (V), saturação por alumínio (m) e matéria orgânica (MO) em três áreas de produção de Banana (APB1, APB2, APB3), três de Mandioca (APM1, APM2, APM3), Capoeira (CAP) e Floresta Plantada (FLO), no Projeto de Assentamento 26 de Março, Marabá, Pará, no ano de 2019
| Áreas | Areia | Silte | Argila | Classe Textural | ||||||||||
| -------------------%-------------------- | ||||||||||||||
| APB1 | 57 | 9 | 34 | Franco argilo arenoso | ||||||||||
| APB2 | 65 | 8 | 27 | Franco argilo arenoso | ||||||||||
| APB3 | 50 | 10 | 40 | Argilo Arenosa | ||||||||||
| APM1 | 64,50 | 7,5 | 28 | Franco-argilo-arenosa | ||||||||||
| APM2 | 58,25 | 8,5 | 33,25 | Franca | ||||||||||
| APM3 | 87 | 4 | 9 | Areia franca | ||||||||||
| CAP | 53 | 9 | 38 | Argilo Arenosa | ||||||||||
| FLO | 75,75 | 5,5 | 18,75 | Franco-arenosa | ||||||||||
| Áreas | pH | Ca | Mg | K | P | Zn | Al | H+Al | CTC | V% | m% | MO | ||
| CaCl2 | cmolcdm-³ | -----mgdm-³------ | ----cmolcdm-3---- | ------%------ | g dm-³ | |||||||||
| APB1 | 3,10 | 1,20 | 0,40 | 84,40 | 2,60 | 1,16 | 1,40 | 7,60 | 9,41 | 19,53 | 43,54 | 19,00 | ||
| APB2 | 4,43 | 2,30 | 0,60 | 92,36 | 5,16 | 1,96 | 0,40 | 2,80 | 5,93 | 52,78 | 11,31 | 19,00 | ||
| APB3 | 4,06 | 1,56 | 0,43 | 86,83 | 4,13 | 1,0 | 1,40 | 7,56 | 9,79 | 22,78 | 38,60 | 20,66 | ||
| APM1 | 4,37 | 1,62 | 0,62 | 200,7 | 7,8 | 2,37 | 0,57 | 3,4 | 6,16 | 45,01 | 17,77 | 17,75 | ||
| APM2 | 3,87 | 0,72 | 0,25 | 92,37 | 5,17 | 1,87 | 1,20 | 6,02 | 7,23 | 17,20 | 49,41 | 18,00 | ||
| APM3 | 5,27 | 2,25 | 0,35 | 62,75 | 6,00 | 1,22 | 0,02 | 1,80 | 4,56 | 59,32 | 0,97 | 16,50 | ||
| CAP | 3,86 | 1,26 | 0,46 | 78,23 | 2,20 | 1,13 | 1,53 | 7,36 | 9,30 | 21,98 | 44,02 | 19,00 | ||
| FLO | 4,77 | 1,75 | 0,72 | 44,92 | 1,5 | 0,95 | 0,20 | 2,67 | 5,26 | 46,87 | 10,88 | 22,25 | ||
O delineamento experimental utilizado foi o inteiramente casualizado, com três repetições e oito tratamentos constituídos pelas áreas do Quadro 1. Em cada vaso foram colocadas cinco sementes de feijão caupi cultivar BRs 17 Gurguéia. Após sete dias do plantio, o desbaste foi realizado deixando apenas duas plantas por vaso. As plantas foram regadas periodicamente, de forma manual, com auxílio de um Becker.
Após 35 dias da emergência, as plantas foram colhidas para as determinações do número de nódulos (NN) e matéria seca da parte aérea (MSPA) e da raiz (MSR). As raízes foram lavadas cuidadosamente em água corrente para posteriormente serem retirados os nódulos com auxílio de uma pinça, sendo os nódulos conservados em sílica gel. Foi usada uma estufa de circulação de ar forçada a 65 oC para determinação da matéria seca da parte área e da raiz, até atingir peso constante.
Os dados do número de nódulos e matéria seca da parte aérea foram submetidos à análise de variância e analisados estatisticamente pelo teste Scott Knott ao nível de 5% de confiança, utilizando O software R livre versão 3.5.2.3. Apenas as médias de número de nódulos e a massa seca de raiz foram transformados para (x+1)^0,5.
Para realizar o isolamento, dez maiores nódulos de cada tratamento foram lavados em água corrente e armazenados em tubos contendo sílica gel. No isolamento, os nódulos foram reidratados em água destilada autoclavada e, posteriormente, esterilizados com álcool 95% (30 segundos), hipoclorito de sódio 2% (2 minutos) (Hungria e Araújo, 1994) e em seguida, submetidos a lavagens sucessivas em água destilada estéril.
Em seguida, foram macerados em placas contendo meio de cultura sólido à base de manitol-extrato de levedura (YMA) (Fred e Waksman, 1928) com corante azul de bromotimol, sendo o material espalhado em forma de estrias compostas para a obtenção de colônias isoladas.
A caracterização morfológica foi realizada em meio sólido YMA com o indicador ácido básico azul de bromotimol (Fred e Waksman, 1928). Os isolados foram caracterizados através dos seguintes parâmetros: 1) Forma da colônia: punctiforme; circular; irregular; filamentosa; 2) Elevação da colônia: plana, lenticular, convexa, pulvinada; 3) Borda da colônia: lisa rugosa, papilada (Melloni et al., 2006).
A caracterização fisiológica também foi realizada em meio YMA (Fred e Waksman, 1928) indicador azul de bromotimol, considerando os parâmetros: 1) tempo de crescimento: muito rápido (1 dia), rápido (2 a 3 dias), intermediário (4 a 5 dias), lento (6 a 15 dias) (Martins et al., 1997); 2) formação de ácidos e álcalis (ácida, neutra, alcalina); 3) volume do muco (abundante, moderada, pouco escassa); 4) elasticidade do muco (gomosa, viscosa, seca); 5) Cromogenia ou Pigmentação da colônia (amarela, branca, creme, incolor) (Melloni et al., 2006).
RESULTADOS E DISCUSSÃO
A nodulação do feijão caupi cultivado em condições controladas, em solo de diferentes áreas, é indicativo da ocorrência de rizóbios. Não há relatos de histórico de inoculação de rizóbios nesta cultura na região. Portanto, as bactérias que induziram a nodulação do feijão caupi são de comunidades nativas. Da mesma forma, a variação na nodulação e a resposta do caupi, sob influência dos diferentes sistemas de uso do solo e dos manejos estudados, evidenciam a natureza da composição e a afinidade das comunidades de rizóbios suportadas por cada área.
A avaliação do NN é um dos critérios que faz parte do protocolo de avaliação da eficiência simbiótica entre rizóbios e leguminosas. Porém, para o feijão caupi, não existem informações conclusivas sobre o número mínimo de nódulos necessários para garantir bom desempenho da FBN (Melo e Zilli, 2009), como é observado para a cultura da soja que indica como suficientes 15 a 20 nódulos na coroa da raiz principal (Hungria e Bohrer, 2000). No entanto, estudos com a cultivar BR 17 Gurguéia indicaram que a capacidade de inoculação dela é maior em relação a outros genótipos de feijão caupi de origem brasileira, nigeriana e norte-americana (Xavier et al., 2006). Melo e Zilli (2009) também relatam maior capacidade de inoculação para essa cultivar.
O número de nódulos encontrados no presente estudo variou de 1,22 a 53,55 planta-1 para os tratamentos APB1 e APM3, respectivamente. Ferreira et al. (2018), em condições de campo com plantas de feijão caupi na fase de pré-florescimento, no município de Marabá, Pará, encontraram uma média de 20 NN planta-1. Já Cavalcante et al. (2017) encontraram uma média de 25,80 NN planta-1 em feijão cultivado sem inoculante, com experimento em campo no município de Bananeiras, Paraíba, Brasil. Esses resultados foram referentes ao comportamento da nodulação do feijão caupi sob diferentes condições.
O tratamento APM3 foi o que apresentou maior número de nódulos. Este resultado pode estar relacionado não só com a população nativa, mas também com as características de fertilidade deste solo (Quadro 2). O solo que constitui o tratamento APM3 apresenta o maior pH, concentração de P e saturação por base. As menores médias para a referida variável (APB1, APM2, CAP e FLO) foram obtidas em plantas cultivadas em solos oriundos com pH inferior a 4, com exceção da área de Floresta (FLO), que apresenta pH 4,77 e menor concentração de P. Desta forma, acredita-se que a acidez e o teor de P foram os fatores mais limitantes para formação de nódulos por estirpes nativas nessas áreas.
O P é um nutriente importante para o estabelecimento de nodulação, pois aumenta o número de pelos radiculares, proporcionando mais sítios de infecção para as bactérias fixadoras de N2 (Okeleye e Okelana, 1997). Além disso, a eficiência do processo de fixação do N2 é dependente da disponibilidade de P devido à sua participação no processo simbiótico (Burity et al., 2000). Othman et al. (1991) verificaram redução no número e na massa seca de nódulos, em feijão caupi, sob níveis baixos de P no solo. Igualmente, Okeleye e Okelana (1997) demonstraram aumento significativo na nodulação do feijão caupi em doses crescentes de P.
A menor produção de matéria seca da parte aérea do feijão caupi foi obtida nos tratamentos APB2 e CAP, com 0,8 e 0,79 g planta-1, respectivamente. Nas demais áreas, não houve diferença significativa, sendo que a maior média foi de 1,98 g planta -1. Melo e Zilli (2009), cultivando feijão caupi cultivar BRs 17 Gurguéia, encontraram para a massa seca da parte aérea aos 35 dias após a emergência uma média de 0,2 g planta-1 nos tratamentos com nodulação por estirpes nativas. Já para as plantas inoculadas e para as que receberam fertilizante nitrogenado obtiveram-se para a massa seca da parte aérea valores médios de 10,8 e 8,2 g planta-1, respectivamente.
A massa seca de raiz variou de 0,63 a 1,52 g planta-1. Contudo, não se verificou diferença entre os tratamentos (Figura 2). Reis et al. (2017), estudando inoculantes alternativos no feijão caupi, encontraram média de 1,67 g em plantas que não receberam tratamento.
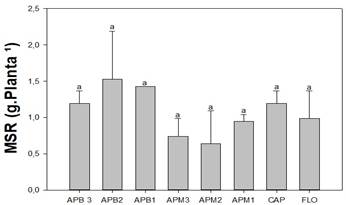
Figura 2 Avaliação do número (NN), matéria seca da parte área (MSPA), raiz (MSR) de feijão caupi cultivados em solo de três áreas de produção de Banana (APB1, APB2, APB3), três de Mandioca (APM1, APM2, APM3), Capoeira (CAP) e Floresta (FLO), no Projeto de Assentamento 26 de Março, Marabá, Pará. Médias seguidas de letras diferentes dentro das colunas indicam diferença significativa (p<0,05) teste Scott Knott.
Os tratamentos CAP e APM3 foram os únicos que apresentaram correlação entre as variáveis NN, MSPA e MSR. Sendo que no tratamento CAP e APM3 foram encontradas as menores e as maiores médias, respectivamente, para as variáveis relacionadas. Essa correlação pode ser explicada pela fertilidade do solo (Quadro 2) e pela eficiência das populações de BNL nativas. Para Silva et al. (2010), a disponibilidade de nutrientes está entre os principais fatores que influenciam a fixação biológica de nitrogênio (FBN).
Foram obtidos um total de 53 estirpes: sob cultivo de banana (16 estirpes), mandioca (20 estirpes), áreas de capoeira (8 estirpes) e floresta plantada (9 estirpes), entre as quais foram encontradas uma grande variedade de estirpes com distintas características fenotípicas (Figura 2). Moreira et al. (1998) destacam que as principais características a indicar a presença de diferentes gêneros de BNLs são o tempo de crescimento e a modificação do pH do meio de cultura.
Morfologicamente, todas as estirpes exibiram colônias circulares, com predominância de elevação planas, com bordas inteiras. Apenas uma estirpe oriunda da área de capoeira apresentou elevação convexa (Figura 3).

Figura 3 Características morfológicas e fisiológicas dos isolados de rizóbios noduladores de feijão caupi, oriundos de solo sob cultivo de mandioca, banana e áreas de capoeira e floresta no Projeto de Assentamento de 26 Março, Marabá, Pará. FC (forma da colônia); EL (Elevação a borda); FB (forma da borda); TC (Tempo de Crescimento); pH (formação de ácidos e álcalis); PM (produção de muco); CM (consistência do muco); Cor.
Quanto à caracterização fenotípica das estirpes nodulíferas através da morfologia cultural das colônias, verificou-se que 88,67% das bactérias apresentaram crescimento rápido, 5,66% intermediário e 5,66% lento (Figura 3). A presença de estirpes nodulíferas que apresentam crescimento rápido é comum em regiões com limitações de umidade e de altas temperaturas. Esta característica constitui uma estratégia de sobrevivência, já que são mais tolerantes à seca do que as de crescimento lento e se multiplicam rapidamente em curto espaço de tempo úmido, o que explicaria sua maior freqüência nos solos das regiões semi-áridas (Santos et al., 2007; Medeiros et al., 2009). O presente trabalho foi realizado numa zona com elevada umidade, característica típica da região amazônica. No entanto, com um período de estiagem aproximadamente de seis meses, o que pode justificar o elevado número de estirpes de crescimento rápido. Também a prática de corte e de queima adotado pelo sistema de manejo das áreas sob produção de mandioca e de banana, proporcionam condições de estresses.
No que diz respeito a acidificação do meio de cultivo proporcionado pelas estirpes, Tan e Broughton (1981) sugerem que essa alteração do pH no meio de cultivo é promovida pela utilização preferencial das estirpes de crescimento rápido pelos açúcares presentes no meio de cultura e pela excreção de ácidos orgânicos após o consumo desse açúcar. No presente estudo, quanto à alteração do pH do meio, 47,16 % apresentaram reação ácida, 54,94 % neutra e 1,88% alcalina em meio YMA acrescido de azul de bromotimol (Figura 3).
O pH é um dos atributos edáficos que limita a presença de microrganismos no solo (Brockwell et al., 1991). Figueiredo et al. (1996) verificaram que a ocorrência de rizóbio de crescimento rápido ou lento está relacionada com o pH do solo. Giongo et al. (2007), avaliando fatores ambientais do solo que afetavam a diversidade de populações de Brady rhizobium spp., isolados de nódulos de soja, observaram que o pH do solo foi a principal característica que afetou a diversidade das populações, e que a menor diversidade foi encontrada nos solos com pH mais ácido. Essa característica está relacionada com os resultados obtidos neste trabalho, em que os isolados que apresentaram reação ácida foram isolados de áreas com pH variando de 3,10 a 5,27 (Quadro 2).
Outra evidência de adaptação é a produção de muco como um mecanismo de adaptação e sobrevivência a condições ambientais adversas (Teixeira et al., 2010; Araújo e Gualter, 2017; Almeida et al., 2018; Amorim et al., 2019), o que reforça o efeito nos resultados encontrados nas diferentes áreas estudadas em virtude dos sistemas de manejo e uso: do total de 53 estirpes analisadas, 43 apresentaram produção moderada de muco, 5 abundante e 1 muito abundante.
A produção de muco pode ser associada à habilidade de crescimento rápido apresentada pelo total de estirpes. Dentre os gêneros que apresentaram estas características fisiológicas estão Rhizobium (Hungria et al., 2016), Sinorhizobium (Matsubara e Zúñiga-Dávila, 2015), Allorhizobium, e Mesorhizobium (Marchetti et al., 2017). A diversidade cultural encontrada no presente estudo demonstra a resiliência das bactérias diante das modificações implementadas pelos diferentes sistemas de cultivo de mandioca e de banana.
Essa diversidade cultural encontrada comprova o caráter promíscuo do feijão caupi em relação à nodulação, corroborando resultados encontrados por outros autores (Melloni et al., 2006; Guimarães et al., 2012; Almeida et al., 2018). Na literatura há relatos da nodulação pelos gêneros Bradyrhizobium, Burkholderia, Azorhizhobium (Zilli et al., 2006; Zhang et al., 2007; Guimarães et al., 2012) e por outros gêneros (Agrobacterium, Pseudomonas, Enterobacter, Pantoea, Bacillus e Paenibacillus), os quais, apesar de serem abundantes no solo e serem reconhecidamente eficientes em promover o crescimento vegetal através de outros processos diferentes da FBN, não são reconhecidos como nodulando leguminosas (Kan et al., 2007; Li et al., 2008; Shiraishi et al., 2010; Marra et al., 2012; Costa et al., 2013). Segundo Kan et al. (2007) essas bactérias, possivelmente, penetram no tecido vegetal juntamente com as nodulíferas durante a infecção e a formação dos nódulos. Porém, algumas bactérias endofíticas de nódulos poderão evoluir para bactérias simbióticas por meio da transferência horizontal de genes simbióticos (Shiraishi et al., 2010; Li et al., 2008).
CONCLUSÕES
A nodulação ocorreu em todos os solos estudados no Projeto de Assentamento 26 de Março, em Marabá, Estado do Pará, Brasil, apresentando uma grande variação de número de nódulos e de produção de matéria seca da parte aérea. Os isolados apresentaram elevada diversidade cultural com características encontradas nos gêneros nodulíferos e não nodulíferos do feijão caupi. Estudos futuros sobre caracterização molecular e sobre a eficiência destes isolados na fixação biológica de N2 em feijão caupi serão importantes para encontrar estirpes inoculantes e para estudos de promoção de crescimento por meio de outros mecanismos.

