











 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUÇÃO
A Cerejeira-do-mato (Eugenia involucrata DC.) pertence à família Myrtaceae e possui ocorrência natural em vários estados brasileiros, distribuindo-se do Rio Grande do Sul até Minas Gerais, além de outros países da América do Sul, como Argentina, Uruguai e Paraguai (Lorenzi, 2016). A espécie apresenta diversas características desejáveis sendo muito utilizada para uso madeireiro, frutícola, paisagístico, ambiental e medicinal. Entretanto, sua propagação pela via seminal é bastante dificultada, pois suas sementes são consideradas recalcitrantes, isto é, não se mantêm viáveis durante longos períodos de armazenamento (Backes e Irgang, 2002; Carvalho, 2014).
Por apresentar elevado potencial econômico e ambiental, justificam-se estudos relacionados à produção de mudas via propagação vegetativa (Degenhardt et al., 2007; Sartoretto et al., 2008), e, particularmente, pela micropropagação. Esta modalidade da cultura de tecidos é um método muito estudado em diferentes espécies vegetais e que mais tem se difundido e encontrado aplicações práticas comprovadas. Entretanto, o emprego da micropropagação em escala comercial visando à produção de mudas pode ser limitado, devido, entre outros fatores, ao elevado custo para obtenção da muda (Erig e Schuch, 2005), para isso cada espécie necessita de ajustes em seus protocolos a fim de otimizar as condições de cultivo das plantas (Sartor et al., 2013), e reduzir custos.
Dentre os fitorreguladores passíveis de utilização na micropropagação, o Ácido giberélico (GA3), que é uma das mais importantes giberelinas como produto comercial (Xavier et al., 2013), pode promover um aumento na divisão celular. Essa giberelina, geralmente, tem, como efeito fisiológico, a indução ao alongamento dos brotos, favorecendo a propagação in vitro (Taiz et al., 2017).
Além dos fitorreguladores, podem ser adicionados ao meio nutritivo, também, aditivos, como é o caso da Polivinilpirrolidona (PVP), que diminuam a intoxicação dos explantes pelos compostos fenólicos oxidados produzidos pelos próprios tecidos vegetais. Estes compostos são, geralmente, liberados nas zonas em que os explantes sofrem cortes, dificultando, assim, o desenvolvimento in vitro da planta (Bezerra et al., 2014). Altas concentrações desses compostos fenólicos podem ocasionar a morte do material vegetal e a abscisão foliar precoce, que é acarretada pelo acúmulo de etileno nos tecidos cultivados in vitro, o qual torna o explante mais frágil e reduz o seu desenvolvimento in vitro (Kerbauy, 2008). A oxidação fenólica é uma das principais limitações da propagação in vitro, especialmente quando se trabalha com espécies lenhosas (Sartor et al., 2013).
O PVP é um antioxidante que tem sido amplamente utilizado nos cultivos in vitro e tem como principal vantagem a captação de elétrons, sendo indicado no controle da oxidação fenólica (Cid e Teixeira, 2014). Adicionalmente à função antioxidante o PVP pode, também, regular o crescimento in vitro das plantas (Xavier et al., 2013).
Diante do exposto, este trabalho teve por objetivo avaliar o efeito do antioxidante PVP no meio nutritivo, no controle da oxidação fenólica, e, também, o efeito do GA3 no alongamento in vitro de segmentos nodais de Eugenia involucrata.
MATERIAL E MÉTODOS
O trabalho, realizado no Laboratório de Biotecnologia e Melhoramento, do Departamento de Fitotecnia, Universidade Federal de Santa Maria (UFSM), Santa Maria, RS, constituiu-se de dois experimentos, independentes um do outro, conduzidos em delineamento inteiramente casualizado. O primeiro, teve a finalidade de avaliar somente o efeito da Polivinilpirrolidona (PVP), apresentou arranjo unifatorial e foi constituído por concentrações do antioxidante, e o segundo, bifatorial (4x2), somente o efeito de combinações de concentrações do Ácido Giberélico (GA3) e períodos de cultivo no alongamento in vitro de explantes de Eugenia involucrata.
Foram utilizados, como explantes, segmentos nodais que não fossem extremamente lenhosos, com aproximadamente 1cm de comprimento e 167 mm de diâmetro, de espécimes de Eugenia involucrata cultivados em casa de vegetação com aproximadamente 9 anos de idade. No laboratório, as brotações foram lavadas com o auxílio de água corrente e detergente comercial e, após, foram enxaguadas duas vezes com água destilada. Na sequência, em câmara de fluxo laminar, foram expostos, por 1min, à solução de etanol a 70% (v/v), e, a seguir, submetidos à imersão em solução de hipoclorito de cálcio a 3,0% (v/v) durante 15 min e, após, em solução de hipoclorito de sódio a 2,0% (v/v) por 15min. Por fim, os explantes foram enxaguados três vezes com água destilada e autoclavada.
Em ambos experimentos, utilizou-se o meio nutritivo MS (Murashige e Skoog, 1962), cuja concentração de sais foi reduzida à metade (½MS), o qual foi acrescido de 10 g L-1 de sacarose, 50 mg L-1 de mio-inositol, 4 g L-1 de ágar e pH, ajustado para 6,0, conforme metodologia de Stefanel (2016). A unidade experimental de ambos os experimentos foi composta por um frasco de vidro com capacidade para 150 mL, contendo 30 mL de meio nutritivo, e três explantes. O primeiro experimento foi composto por quatro tratamentos e 10 repetições, totalizando 40 unidades experimentais e 120 explantes. O segundo experimento foi composto por oito tratamentos e 10 repetições, totalizando 80 unidades experimentais e 240 explantes.
Anteriormente à inoculação dos explantes e após a adição do ágar, o meio nutritivo foi autoclavado a 121ºC e 1 atm durante 15 min. Os frascos foram vedados com papel alumínio e as culturas foram mantidas em sala de crescimento com temperatura controlada de 25±2°C, fotoperíodo de 16 h e intensidade luminosa de 20 μmol m-2 s-1, obtida a partir de lâmpadas fluorescentes brancas frias tipo luz do dia.
No primeiro experimento, os tratamentos consistiram das concentrações de PVP (0; 0,5; 1,0 ou 1,5 g L-1), sendo a ausência de PVP no meio nutritivo o tratamento testemunha. Foram avaliadas, após 30 dias de cultivo in vitro, as variáveis: sobrevivência in vitro (indicada pela coloração verde do explante), estabelecimento in vitro (determinado pelo desenvolvimento de primórdios foliares no explante) e oxidação fenólica (foram contabilizados como oxidados os explantes que se apresentaram totalmente e/ou com pelo menos a metade do seu tamanho na coloração marrom escuro), todas expressas em porcentagem.
Já no segundo experimento, os tratamentos consistiram da combinação das concentrações de GA3 (0; 1; 2 ou 4 µM), sendo a ausência de GA3 no meio nutritivo, o tratamento testemunha, com os dois períodos de cultivo in vitro (30 e 60 dias). Foram avaliadas as variáveis: sobrevivência in vitro (indicada pela coloração verde do explante) expressa em porcentagem, altura média dos brotos (cm) e número de brotos. A variável altura de brotos foi mensurada, por meio de uma régua, avaliando-se todas as brotações por explante para cada tratamento e repetição.
Após avaliar a normalidade dos erros pelo teste de Kolmogorov-Smirnov e a homogeneidade de variâncias por meio do teste de Bartlett, os dados foram transformados pela função √𝑥+0,5 e submetidos à análise de variância. Quando o valor de “F” foi significativo, médias de tratamentos qualitativos foram submetidos à comparação de médias por meio do teste de Tukey ao nível de 5% de probabilidade de erro. Médias de tratamentos quantitativos foram submetidas à análise de regressão polinomial. Os resultados apresentados são as médias originais obtidas. Foi utilizado o pacote estatístico Sisvar (Sistema para Análise de Variância) para Windows® versão 5.1 (Ferreira, 2014) para a análise estatística dos dados. Para determinar a precisão dos ensaios foi estimado o Índice de Variação (IV), calculado por CV √𝑁 , em que o IV é igual ao coeficiente de variação (CV) dividido pela raiz quadrada do número de repetições (N) (Pimentel-Gomes, 2009).
RESULTADOS E DISCUSSÃO
No primeiro experimento, em que se testaram diferentes concentrações de PVP, não houve efeito significativo dos tratamentos para as variáveis sobrevivência (p= 0,4033), estabelecimento in vitro (p= 0,2731) e oxidação fenólica (p= 0,6688) (Quadro 1). Observou-se uma média geral alta para a sobrevivência e o estabelecimento in vitro dos explantes (83,10% e 73% respectivamente) e uma média geral relativamente baixa para a variável oxidação fenólica (16,55%).
Quadro 1 Sobrevivência (%), estabelecimento in vitro (%) e oxidação fenólica (%), observadas em segmentos nodais de Eugenia involucrata, após a inoculação em meio ½MS contendo diferentes concentrações de PVP, aos 30 dias de cultivo in vitro
| PVP (g L-1) | Sobrevivência (%) | Estabelecimento in vitro (%) | Oxidação fenólica (%) |
|---|---|---|---|
| Testemunha | 66,40 a* | 53,00 a | 20,00 a |
| 0,5 | 93,20 a | 73,00 a | 13,20 a |
| 1,0 | 86,40 a | 79,60 a | 26,40 a |
| 1,5 | 86,40 a | 86,40 a | 6,60 a |
| Média | 83,10 | 73,00 | 16,55 |
| IV | 3,44 | 3,77 | 5,65 |
*Na coluna, médias seguidas pela mesma letra minúscula, não diferem significativamente pelo teste F ao nível de 5% de probabilidade de erro. A letra ‘a’ representa o resultado mais favorável para as variáveis analisadas. IV (Índice de variação) = 𝐶𝑉 √𝑁 , em que CV= coeficiente de variação e N= número de repetições.
A alta sobrevivência e estabelecimento observadas, corroboram com outro estudo realizado com a espécie em que, da mesma maneira que no presente trabalho (Figura 1), a presença de oxidação fenólica não inviabilizou o estabelecimento nem, tampouco, o desenvolvimento dos segmentos nodais de Eugenia involucrata (Golle et al., 2012). Ademais, nas avaliações do ensaio, observou-se que, geralmente, a região oxidada desprendia-se facilmente do explante e, abaixo dessa região, existiam novos tecidos. A descamação é uma característica comum da espécie durante o seu crescimento, o que pode ter sido o motivo que desencadeou a oxidação observada. Por isso, pode-se justificar que, por ser uma característica comum da espécie (Golle, 2010), a oxidação fenólica presente nos explantes não tenha prejudicado sua sobrevivência e estabelecimento in vitro.
De maneira contrária ao obtido, no cultivo in vitro de Handroanthus chrysotrichus foi observada uma maior sobrevivência dos segmentos nodais da espécie quando inoculados em meio ½WPM (Lloyd e McCown, 1980) contendo 1g L-1 de PVP (Rabaiolli, 2014) sugerindo, com isso, que a presença ou não de PVP no meio nutritivo é relativo de cada espécie estudada.

Figura 1 Oxidação fenólica em segmentos nodais de Eugenia involucrata, inoculados em meio nutritivo MS com a concentração de sais reduzida à metade (½MS) contendo 1,0 g L-1 de PVP, após 30 dias de cultivo in vitro, cuja oxidação não prejudicou o desenvolvimento das gemas axilares pré-existentes. Barra = 1cm. Santa Maria, RS, UFSM, 2017.
A presença do antioxidante, independentemente da concentração utilizada (0,5; 1,0 ou 1,5 g L-1), não proporcionou eficiência no controle da oxidação fenólica dos segmentos nodais de Eugenia involucrata. Isso pode ter acontecido, provavelmente, em função de que a espécie é lenhosa, necessitando, assim, de elevadas concentrações do antioxidante para diminuir os compostos fenólicos.
Corroborando em parte, para Dalbergia nigra, a presença de 1 g L-1 de PVP, tanto no meio nutritivo MS quanto no WPM, ocasionou as maiores média de oxidação dos explantes (Sartor et al., 2013). Entretanto, o contrário foi observado em Theobroma grandiflorum, em que a presença de somente 0,4 g L-1 de PVP no meio DKW (McGranahan et al., 1987) foi eficiente no controle da oxidação fenólica dos explantes (Almeida et al., 2010). Estes resultados sugerem, novamente, que a resposta à concentração de PVP é muito dependente da espécie.
No segundo experimento, em que foram testadas diferentes concentrações de GA3, foi observado efeito significativo somente do fator principal período de cultivo para a variável sobrevivência in vitro (p=0,0073) (Quadro 2). Já para altura média dos brotos (p=0,0275) e número de brotos (p=0,0004), observaram-se efeito significativo somente do fator principal GA3. Para a altura média dos brotos, a equação estimada que resultou no melhor ajuste foi a linear crescente (Figura 2), ao passo que, para o número de brotos, foi uma equação linear decrescente (Figura 3).
Em relação ao período de cultivo, a maior média de sobrevivência ocorreu aos 30 dias (88,10%), observando-se um decréscimo significativo aos 60 dias (61,45%) (Quadro 2), o que pode ter sido ocasionado pela menor quantidade de nutrientes disponíveis aos explantes, devido ao maior tempo de permanência no mesmo meio nutritivo. Isso também foi observado em outro estudo com a mesma espécie, em que foi obtida, aos 30 dias de cultivo, 71,12% de sobrevivência dos explantes enquanto aos 60 dias foi observado um decréscimo significativo, com apenas 52,48% dos explantes vivos (Stefanel, 2016). Isso pode sugerir que a queda na sobrevivência de Eugenia involucrata aos 60 dias de cultivo pode ser considerada uma característica comum do comportamento in vitro dessa espécie.
Quadro 2 Sobrevivência (%) observada em segmentos nodais de Eugenia involucrata aos 30 e 60 dias de cultivo in vitro, após a inoculação em meio ½MS contendo diferentes concentrações de GA3
| Dias | Sobrevivência (%) |
|---|---|
| 30 | 88,10 a* |
| 60 | 61,45 b |
| Média | 74,77 |
| IV | 4,30 |
*Na coluna, médias seguidas pela mesma letra minúscula, não diferem significativamente pelo teste F ao nível de 5% de probabilidade de erro. A letra ‘a’ representa o resultado mais favorável para as variáveis analisadas. IV (Índice de variação) = 𝐶𝑉 √𝑁 , em que CV= coeficiente de variação e N= número de repetições.
Para a variável altura média dos brotos obteve-se a maior média (0,52 cm) na presença da maior concentração da giberelina (4 μM), ao passo que, quando se diminuiu a concentração de GA3 no meio nutritivo, a altura dos brotos também reduziu, sendo que, na ausência da giberelina obteve-se a menor média (0,12 cm) (Figura 2). O resultado obtido era esperado, visto que o GA3 é uma substância conhecida por possuir efeito no alongamento celular das plantas (Taiz et al., 2017).
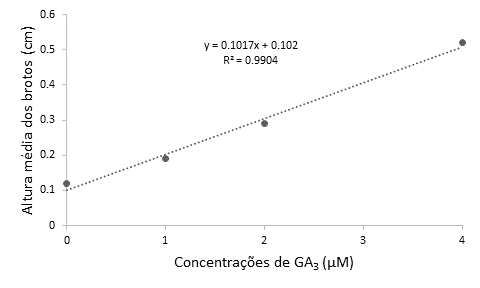
Figura 2 Altura média dos brotos (cm) obtidos a partir de segmentos nodais de Eugenia involucrata, em função das concentrações de GA3 (0; 1; 2 ou 4 μM) adicionadas ao meio nutritivo MS, cuja concentração de sais foi reduzida à metade (½MS), independentemente do período de cultivo in vitro. Santa Maria, RS, UFSM, 2017.
Os resultados obtidos corroboram e parte com o verificado em Maclura tinctoria, em que as concentrações de 10,97 μM e 16,46 μM de GA3 no meio nutritivo WPM induziram o crescimento dos brotos (Gomes et al., 2010). Da mesma maneira, para o cultivo in vitro de Cattleya tigrina a adição de GA3 no meio nutritivo se fez necessária para a obtenção de brotações mais alongadas (Fritsche, 2012). Porém, o contrário foi observado em diferentes genótipos de Eucalyptus dunnii Maiden, em que a presença do ácido giberélico no meio nutritivo ½MS não foi favorável ao alongamento dos explantes, além disso, a giberelina proporcionou menor tamanho das brotações e formação de estruturas calogênicas (Navroski et al., 2013), o que pode ter sido decorrente de um possível efeito inibitório da giberelina sobre o alongamento celular da espécie.
Para a variável número de brotos obteve-se a maior média na ausência da giberelina no meio nutritivo (4,5), ao passo que na presença da sua maior concentração (4 μM de GA3) observou-se o menor número de brotos (0,15) (Figura 3). Esses resultados sugerem um efeito inibitório do GA3 sobre o número de brotos em Eugenia involucrata à medida em que se aumentou a concentração da giberelina no meio nutritivo até 4 µM (Figura 4). Esse comportamento é reforçado pelos resultados de alguns estudos que relataram que o uso de GA3 pode inibir a proliferação de partes aéreas em plantas (Grattapaglia e Machado, 1998).
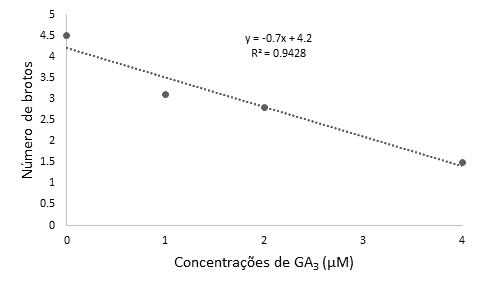
Figura 3 Número de brotos obtidos a partir de segmentos nodais de Eugenia involucrata, em função das concentrações de GA3 (0; 1; 2 ou 4 μM) adicionadas ao meio nutritivo MS, cuja concentração de sais foi reduzida à metade (½MS), independentemente do período de cultivo in vitro. Santa Maria, RS, UFSM, 2017.

Figura 4 Representação ilustrativa das brotações de Eugenia involucrata cultivadas in vitro, em função de diferentes concentrações de GA3 adicionadas ao meio nutritivo ½MS, independentemente do período de cultivo in vitro. Observa-se em “A” brotações na ausência do fitorregulador; “B” brotações na presença de 1 µM de GA3; “C” brotações na presença de 2 µM de GA3; e “D” brotações na presença de 4 µM de GA3. Barra = 1cm. Santa Maria, RS, UFSM, 2017.
Comportamento similar foi observado em outro estudo com Eugenia involucrata, em que, na ausência da giberelina, foi obtido o maior número de brotos, já, na presença de 20 µMm a 40 µM, os brotos entraram em senescência e acabaram se inviabilizando, o que conforme Golle (2010) teria sido devido à toxidez causada pelas concentrações elevadas dessa giberelina. Também, na espécie Mentha x Piperita, o maior número de brotos por explantes foi observado na presença de 0,15 µM de GA3 associado à auxina 6-benzilaminopurina (BAP) a aproximadamente 2 µM, sendo obtidos até 6,08 brotos/explante (Morais et al., 2014).
Em contrapartida, em explantes juvenis de Carica papaya, a aplicação de aproximadamente 0,3 µM de GA3 em meio MS, além de contribuir para o maior tamanho em altura das plantas, proporcionou um maior número de brotações por explante (1,84), principalmente quando a giberelina foi associada a aproximadamente 0,5 µM de 2-isopenteniladenina (2iP) (Vidal et al., 2013). Isso pode ter ocorrido devido a um sinergismo entre a citocinina e o ácido giberélico, indicando que associação desses fitorreguladores podem estimular a multibrotação dos explantes.
De uma maneira geral, concentrações superiores a 2 µM de GA3 prejudicam a multiplicação in vitro de Eugenia involucrata, porque, apesar de alongar os brotos, acabam reduzindo o número de brotos por explantes, o que não é desejável na micropropagação, pois acaba reduzindo a taxa de multiplicação da espécie.
CONCLUSÕES
É dispensável a utilização de PVP no meio nutritivo no cultivo in vitro de segmentos nodais de Eugenia involucrata.
O ácido giberélico (GA3) apresenta efeito positivo no alongamento in vitro de Eugenia involucrata.
Na presença de concentrações superiores a 2 µM de GA3 em meio nutritivo ½MS, há uma redução no número de brotos por explante.

