











 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUÇÃO
As ovelhas Serra da Estela constituem atualmente o maior efetivo das raças autóctones portuguesas exploradas para a produção de leite. O nível produtivo médio estimado destas ovelhas, 155,1±47,6 litros aos 150 dias de lactação (SPOC, 2023), possibilita que a maior parte da produção de leite possa ser efetuada exclusivamente com a ingestão de erva. Nas condições climáticas mediterrânicas, este maneio alimentar pode ser exequível com a utilização de pastagens de sequeiro e alguma rega de apoio, permitindo uma disponibilidade contínua de erva para pastoreio entre setembro e maio, contornando assim a imprevisibilidade da precipitação atmosférica, que obrigam os agricultores ao sistemático recurso à suplementação e a alimentos conservados. Uma utilização eficiente dos recursos pratenses dependerá do ajustamento entre a quantidade e qualidade da forragem disponível e do cuidado a ter com o fornecimento de alimentos concentrados em energia e proteína, que em ovelhas Serra da Estrela se refere apenas às primeiras semanas de lactação, em que produção leiteira aumenta acentuadamente e as ovelhas com partos múltiplos têm a sua ingestão de alimentos fibrosos diminuída, resultando numa mobilização acentuada das reservas corporais, que se pretende seja limitada temporalmente.
A eficiência de captura do azoto pelos microrganismos do rúmen depende da degradabilidade da proteína da dieta e também da existência de energia facilmente utilizável, indicando Botts et al. (1979) que o excesso no fornecimento de proteína na dieta provoca perdas energéticas relacionadas com a síntese da ureia no fígado, a partir da amónia em excesso. Assim, o fornecimento de fontes proteicas de diferente degradabilidade no rúmen tem como objetivo, não só suprir a quantidade de proteína total de acordo com as necessidades produtivas (NRC, 2007), proveniente da síntese microbiana e do alimento, mas também a melhoria da composição em aminoácidos (AA) dessa proteína (Whitelaw et al., 1986). As pastagens de gramíneas e leguminosas das regiões temperadas são ricas em proteína muito degradável, superior a 70% da PB total (Hongerholt & Muller, 1998). Em vacas leiteiras, vários autores referem que a suplementação com proteína não degradável no rúmen (RUP) aumenta o fluxo de AA para o intestino delgado, além do proporcionado pelos AA provenientes da proteína microbiana produzida no rúmen. Broderick et al. (2002) referiram um aumento da produção de leite, utilizando bagaço de soja por pressão como suplemento. Em estudos com ovelhas leiteiras, Mikolayunas-Sandrock et al. (2009) utilizando feno como alimento base e Mikolayunas et al. (2011) com ovelhas em pastoreio, estudaram o efeito do nível da RUP nas dietas. Também Belo (2000), num ensaio com ovelhas Serra da Estrela estabuladas no início da lactação alimentadas com erva, e posteriormente com ovelhas em pastoreio (Belo et al., 2019), conseguiram melhores produções de leite em ovelhas suplementadas com glúten de milho em comparação com as suplementadas com milho.
O presente estudo pretende conhecer: 1) a importância do glúten de milho como suplemento de ovelhas a consumir forragem produzida em pastagens de gramíneas e leguminosas, durante a fase inicial da lactação; 2) avaliar o nível proteico mais adequado para a produção e composição do leite e; 3) identificar os parâmetros metabólicos que melhor caracterizam o metabolismo intermédio das ovelhas nesta fase da lactação.
MATERIAL E MÉTODOS
Foram utilizadas 27 ovelhas da raça Serra da Estrela (nove por tratamento) escolhidas com os seguintes critérios: condição corporal (CC) ao parto ≥ 2,5; coincidência em relação à data de parto e equilíbrio dos grupos em relação ao número de borregos aleitados por ovelha. As ovelhas foram alojadas em parques individuais na última semana de gestação. Após o nascimento os borregos permaneceram com as mães todo o dia, até aos 21 dias de idade. A partir desta altura (desmame parcial) até ao desmame total aos 42 dias de idade, os borregos foram separados diariamente, entre as 17:30 e as 08:00h do dia seguinte, juntando-se às mães após estas serem ordenhadas manualmente. Durante o período de separação, os borregos tiveram acesso a feno de luzerna (17% de PB na MS) e alimento concentrado de iniciação (18% de PB na MS).
As ovelhas receberam erva cortada diariamente de uma pastagem composta por azevém (Lolium perene, cv. Victorian), trevo branco (Trifolium repens, cv. Pitau), trevo morango (Trifolium fragiferum, cv. Palestine) e festuca (Festuca arundinacea, cv. Clarine). A erva foi distribuída ad libitum, sendo considerada uma ingestão de 1,30 kg de matéria seca (MS) ovelha.dia-1 baseada num trabalho anterior (Belo, 2000) e um teor de 20,5% de PB, resultado de avaliações prévias da pastagem. Foram estabelecidos 3 suplementos isoenergéticos, que foram distribuídos numa quantidade que garantiu o fornecimento de 0,9 Mcal de energia metabolizável (EM) e três níveis de PB na dieta (17, 21 e 25% na MS). Os suplementos alimentares utilizados foram mandioca (MAND) e glúten de milho (CGM), e as quantidades atribuídas aos tratamentos (17PB, 21PB e 25PB) encontram-se no Quadro 1. A erva da pastagem foi cortada diariamente, pesada e colocada nos comedouros individuais às 9:00 e retirada às 17:30h, sendo pesados os refugos. Foi considerado um excesso de 15% na erva distribuída, tendo em conta a média do consumo dos dois dias anteriores. Às 18:00h procedeu-se à distribuição do suplemento. A avaliação da qualidade nutritiva da erva distribuída baseou-se em duas amostras colhidas no início e a meio de cada semana, imediatamente antes da distribuição aos animais. A composição florística foi determinada por amostragem semanal feita na faixa que iria ser cortada nesse dia.
Quadro 1 Ingredientes dos suplementos distribuídos
| Suplementos | |||
| (g de MS/dia) | 17% PB | 21% PB | 25% PB |
| Mandioca (MAND)1,2 | 310 | 226 | 142 |
| Glúten de milho (CGM)1,3 | 0 | 78 | 155 |
Notas:
1 MAND: 83,0% MS, 2,9% de PB na MS; CGM: 86,1% MS, 81,3% de PB na MS
2 2,91 Mcal.kg MS-1 de EM (Feedipedia, 2012-2022); 3 3,18 Mcal.kg MS-1 de EM (NRC, 2007)
As ovelhas foram pesadas após o parto e no final do período de ensaio. A avaliação da condição corporal (Russel et al., 1969) foi igualmente realizada nestas duas alturas e no início do período de ordenha parcial (aos 21 dias de lactação). Os borregos foram pesados semanalmente. A quantidade de leite produzido no período de aleitamento total dos borregos foi estimada pelo método da dupla injeção de ocitocina (Geenty & Sykes, 1986), na 2ª semana de lactação. No período de ordenha parcial, as ovelhas foram ordenhadas às 8:00h. Nesta fase procedeu-se ainda à estimativa da quantidade de leite disponibilizado pelas ovelhas aos borregos durante o período das 8:00 às 17:30h e da proporção de leite residual retido após a ordenha das 8:00h. Foi medida a quantidade de leite e recolhidas amostras de todas as frações de leite obtido para a realização de determinações analíticas. Foram determinados os teores em gordura, proteína, lactose e sólidos totais, por espetrofotometria de infravermelhos (Milkoscan™ 133B, Foss Electric).
Realizaram-se colheitas de sangue no 10º, 25º e 39º dias de lactação, às 8:00, 17:00, 24:00h. Os parâmetros doseados foram os seguintes: ácidos gordos não esterificados (AGNE), β-hidroxibutirato (BHBA), creatinina (CREAT), glucose (GLU), lípidos totais (LT), proteína total (PT), triglicéridos (TRIG), ureia (Ureia-N), insulina (INS) e triiodotironina (T3). Os doseamentos foram realizados utilizando kits comerciais, à exceção da GLU em que foi utilizado o método da ortotoluidina em ácido acético glacial (Hyvarinen & Nikkila, 1962), por métodos colorimétricos ou de radioimunoensaio (T3 e INS).
A análise estatística dos dados recolhidos foi baseada em análises de variância e testes de comparações múltiplas. Para as medições repetidas no tempo, como as produções leiteiras e crescimento dos borregos, foi adotado o procedimento MIXED MODEL (SAS Institute Inc., Cary, NC, EUA) com um modelo linear para analisar as diferentes colheitas; no caso do doseamento dos parâmetros sanguíneos, o modelo linear considerou duas medidas repetidas no tempo, colheita e horas de colheita, utilizando o produto de Kronecker, uma matriz de covariância estruturada, de acordo com Bobe et al. (2003).
Sempre que o efeito da interação não foi significativo, este termo foi retirado do modelo. Os testes de comparação múltipla das médias, teste de Tukey, cujos efeitos resultaram significativos, foram realizados utilizando as médias dos mínimos quadrados (SAS Institute Inc., Cary, NC, EUA), considerando o nível de significância de 95%. Foram testados os pressupostos da análise de variância para cada modelo ajustado, nomeadamente a normalidade e a homogeneidade de variâncias aplicando os testes de Shapiro-Wilk e Bartlett, respetivamente, recorrendo à transformação das variáveis quando necessário. No entanto, os valores médios apresentados nos diversos quadros e gráficos são sempre as médias de mínimos quadrados sem transformação de modo a tornar os resultados mais compreensíveis.
RESULTADOS
Qualidade e Composição Florística da Pastagem
Os valores médios, em percentagem da MS, da avaliação analítica das amostras de erva recolhidas estão no Quadro 2. A EM calculada (Boletim Técnico nº33, M.A.F.F., 1975) foi de 2,2±0,685 Mcal.kg MS-1, verificando-se que o teor de PB da MS da erva foi ligeiramente superior ao utilizado para formulação das dietas (20,5%PB). As espécies dominantes foram a festuca e o azevém que, em conjunto, contribuíram com cerca de 60% da MS total disponível na pastagem. O trevo branco e o trevo morango representaram cerca de 22% da MS total, tendo o trevo branco a proporção mais elevada (14,5% do total de MS).
Quadro 2 Composição analítica das amostras de erva recolhidas, e coeficientes de variação
| % da MS | |
| MS1 | 14,9±8,92 |
| PB2 | 21,6±3,24 |
| NDF3 | 46,6±4,01 |
| ADF3 | 30,2±1,94 |
| Cinzas | 13,1±4,93 |
| Digestibilidade da MO4 | 67,1±3,15 |
Notas: Análises realizadas de acordo com a metodologia em vigor na Estação Zootécnica Nacional, INIAV
1 NP ISO 6496:2018; 2 NP EN ISO 005983-2:2009; 3 NP EN ISO 16472:2013 e EN ISO 13906:2008
Consumo de MS e PB
Os consumos de erva registados ao longo das seis semanas não diferiram entre as três dietas (P>0,05), embora as ovelhas submetidas ao tratamento 17PB tivessem consumido em média 1,133 kg MS.dia-1, menos cerca de 0,2 kg MS.dia-1 em relação à média dos outros dois tratamentos. Ao longo do ensaio a ingestão de erva foi semelhante nos três grupos, a média foi de 0,904 kg MS.dia-1 na 1ª semana, valor significativamente inferior (P<0,05) ao observado nas restantes semanas. Em relação ao peso metabólico (kg0,75), o consumo de MS foi crescente para os três tratamentos ao longo das seis semanas (P<0,05) (Figura 1). Na 1ª semana foi semelhante, média de 44,36 g MS.dia-1 por kg PV0,75, mas a partir da 4ª semana, as ovelhas nos tratamentos 21PB e 25PB consumiram quantidades semelhantes de MS, média de 72,28 g.dia-1 por kg PV0,75, e significativamente superiores (P<0,05) ao registado para as ovelhas em 17PB (53,33 g.dia-1 por kg PV0,75).
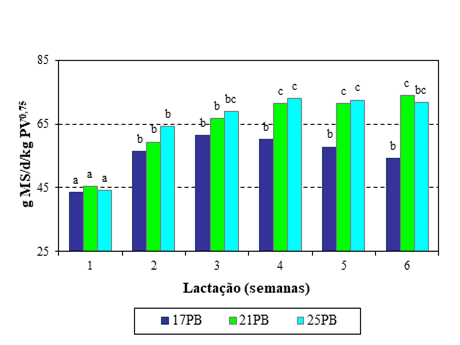
Em relação aos valores diários de ingestão de PB resultantes do consumo de erva, não se registaram diferenças significativas (P>0,05) entre os três grupos de ovelhas. Todavia, as diferenças foram significativas para a dieta total (P<0,05) entre os três níveis de suplementação, valores médios de 253,7, 357,2,6 e 420,0 g PB.dia-1, a que corresponderam concentrações proteicas das dietas ligeiramente superiores ao inicialmente pretendido, respetivamente de 17,6, 21,9 e 25,6% de PB.
Evolução dos pesos vivos e da condição corporal
O peso vivo (PV) médio ao parto foi de 57,8 kg, semelhante para as ovelhas nas três dietas (P>0,05). A maior perda de PV durante as 6 semanas correspondeu às ovelhas a receber a dieta 17PB (-6,44 kg), que foi significativamente superior (P<0,05) à variação de peso (-1,43 kg) das ovelhas com a dieta 25PB, enquanto as ovelhas com 21PB perderam 3,56 kg de PV. A CC avaliada ao parto: 2,58, 2,56 e 2,64 respetivamente para os níveis crescentes de suplementação proteica, não sofreu diferenças significativas (P>0,05) no decorrer do estudo. A perda de CC ocorreu apenas entre o parto e o 21ºdia de lactação, em que as ovelhas com as dietas 17PB, 21PB e 25PB perderam -0,17, -0,25 e -0,08 pontos de CC respetivamente.
A evolução do PV dos borregos foi idêntica para todos os tratamentos ao longo do período de ensaio. A média dos PV registados foi de 4,25, 8,89 e 11,81 kg, respetivamente para os PV ao nascimento, ao desmame parcial (21 dias de idade) e ao desmame total (42 dias de idade).
Produção e qualidade do leite
Os níveis de PB na dieta não tiveram um efeito significativo (P>0,05) na quantidade e composição do leite correspondente à fase de aleitamento total. No entanto, as estimativas da produção de leite obtidas no período de 4 horas para as ovelhas nos níveis 17PB (255,2 ml) e 21PB (257,3 ml) foram superiores às do tratamento 25PB (203,3 ml). Também a composição do leite obtido nesta fase foi semelhante para as ovelhas dos três tratamentos, médias de 5,40%, 3,54%, 3,90% e 13,53% para os teores em gordura (TG), proteína (TP), lactose (TL) e sólidos totais (TST).
A quantidade de leite estimada na fase de aleitamento parcial, correspondente ao período em que os borregos estavam com as mães (9:30h por dia), foi superior, embora não significativamente (P>0,05), para as ovelhas em 21PB (538,1 ml) e semelhantes para os grupos 17PB e 25PB, respetivamente 367,5 e 341,2 ml. A composição do leite revelou teores de TG, TP, TL e TST semelhantes para os três tratamentos, médias de 5,43%, 4,04%, 4,05% e 14,11% respetivamente.
A quantidade média de leite ordenhado, durante a fase de aleitamento parcial dos borregos, foi mais elevada nas ovelhas a receber a dieta 21PB (683,1 ml.d-1; P<0,05) do que nas ovelhas em 17PB (455,2 ml.d-1) e 25PB (465,7 ml.d-1). Durante este período da lactação observou-se um aumento de produção para os três tratamentos, sendo significativa (P<0,05) a diferença entre os valores médios registados na 4ª e na 6ª semana, respetivamente 507,7 e 562,6 ml.d-1. Em relação à composição percentual do leite ordenhado não se verificaram diferenças significativas (P>0,05) entre os níveis de PB na dieta para os três componentes analisados, valores médios de 3,58% para TG, 5,04% para TP e 5,04% para TL.
A quantidade de leite retido pelas ovelhas após a ordenha (leite residual), medido na 5ª semana, foi semelhante (P>0,05): 122,0 ml, 165,0 ml e 114,4 ml, respetivamente para 17PB, 21PB e 25PB. A composição química não foi significativamente diferente (P>0,05) entre os três grupos de ovelhas, registando-se os teores médios de 3,72%; 3,29% e 3,75% para a TG, TP e TL, respetivamente.
Parâmetros metabólicos e hormonais
Em relação à GLU não se verificaram diferenças significativas entre tratamentos (quadro 3), obtiveram-se as concentrações médias de 4,282, 4,289 e 4,655 mmol.l-1 para os tratamentos 17PB, 21PB e 25PB, respetivamente. As concentrações aumentaram no decorrer da lactação e significativamente entre o 10º e o 25º dia, valores de 3,882, 4,575 e 4,770 mmol.l-1 respetivamente. O quadro 4 evidencia as diferentes concentrações de glucose verificadas durante o dia, maiores para o tratamento 17PB, em que as determinações às 17:00h (3,965 mmol.l-1) e às 24:00h (4,748 mmol.l-1) foram significativamente diferentes (P<0,05).
Quadro 3 Valores dos parâmetros metabólicos obtidos de acordo com os tratamentos e dias de colheita, e nível de significância dos efeitos
| Dietas | Dias de lactação | Significância | |||||||
| 17PB | 21PB | 25PB | 10º | 25º | 39º | Dietas | Dias lact. | ||
| GLU, mmol.l-1 | 4,282 | 4,289 | 4,655 | 3,882a | 4,575b | 4,770b | NS | <0,0001 | |
| AGNE, mmol.l-1 | 0,431 | 0,496 | 0,385 | 0,624c | 0,391b | 0,297a | NS | <0,0001 | |
| INS, µUI.ml-1 | 14,63b | 12,87a | 20,47c | 15,18 | 15,87 | 16,92 | 0,0002 | NS | |
| BHBA, mmol.l-1 | 0,687 | 0,697 | 0,615 | 0,770 b | 0,639a | 0,591a | NS | <0,0001 | |
| LT, g.l-1 | 2,31b | 2,07a | 2,05a | 2,26b | 2,12ab | 2,04a | 0,0160 | 0,0113 | |
| PT, g.l-1 | 70,88a | 74,04ab | 77,40b | 74,67 | 73,22 | 74,62 | 0,0018 | NS | |
| Ureia-N, mg.dl-1 | 19,17a | 26,84b | 34,76c | 26,15a | 25,60a | 28,69b | <0,0001 | 0,0304 | |
| TRIG, mmol.l-1 | 0,148 | 0,130 | 0,135 | 0,141 | 0,140 | 0,132 | NS | NS | |
| CREAT, μmol.l-1 | 75,16 | 71,49 | 72,83 | 74,50 | 72,57 | 72,41 | NS | NS | |
| T3, ng.ml-1 | 1,406 | 1,338 | 1,309 | 1,376 | 1,359 | 1,317 | NS | NS | |
Nota: médias com letras diferentes são significantemente diferentes, para P<0,05
Quadro 4 Valores dos parâmetros metabólicos obtidos de acordo com a interação entre os tratamentos e horas de colheita, e nível de significância
| 17PB | 21PB | 25PB | ||||||||
| 08:00h | 17:00h | 24:00h | 08:00h | 17:00h | 24:00h | 08:00h | 17:00h | 24:00h | Significância | |
| GLU, mmol.l-1 | 4,118ab | 3,965a | 4,748b | 4,351ab | 4,167ab | 4,435ab | 4,715ab | 4,586ab | 4,757b | 0,0008 |
| AGNE, mmol.l-1 | 0,691b | 0,369a | 0,219a | 0,661b | 0,328a | 0,433ab | 0,613b | 0,244a | 0,300a | <0,0001 |
| INS, µUI.ml-1 | 12,29ab | 14,43abc | 17,16bc | 8,85a | 13,65ab | 16,12bc | 15,65bc | 20,05c | 25,72d | <0,0001 |
| BHBA, mmol.l-1 | 0,659ab | 0,771c | 0,623ab | 0,710ab | 0,755bc | 0,616a | 0,621ab | 0,653ab | 0,569a | <0,0001 |
| LT, g.l-1 | 2,44c | 2,26b | 2,22b | 2,18b | 2,02a | 2,01a | 2,18b | 2,00a | 1,94a | <0,0001 |
| PT, g.l-1 | 71,66 | 69,80 | 71,07 | 75,17 | 73,37 | 73,72 | 78,26 | 76,80 | 77,68 | NS |
| Ureia-N, mg.dl-1 | 15,47a | 24,45bc | 17,58a | 24,62bc | 32,30d | 23,61b | 34,67e | 39,82f | 29,80cd | <0,0001 |
| TRIG, mmol.l-1 | 0,193 | 0,129 | 0,121 | 0,156 | 0,109 | 0,126 | 0,168 | 0,124 | 0,115 | <0,0001 |
| CREAT, μmol.l-1 | 73,70ab | 78,31b | 73,47ab | 72,28ab | 74,65b | 67,53a | 73,09ab | 74,86b | 70,55a | <0,0001 |
| T3, ng.ml-1 | 1,337 | 1,460 | 1,452 | 1,184 | 1,483 | 1,396 | 1,216 | 1,436 | 1,258 | NS |
Nota: médias com letras diferentes são significantemente diferentes, para P<0,05
O Quadro 3 mostra que os valores da concentração de AGNE foram semelhantes entre os tratamentos, evidenciando-se um decréscimo significativo (P<0,05) ao longo da lactação, entre 0,624 mmol.l-1 no 10º dia e 0,297 mmol.l-1 no 39º dia. Em relação à variação diária, os níveis médios de AGNE foram significativamente superiores (P<0,05) na determinação das 8:00h, em todos os tratamentos (Quadro 4).
A concentração média de INS (Quadro 3) foi significativamente diferente (P<0,05) nos três tratamentos, maior em 25PB (20,47 µU.ml-1), menor em 21PB (12,87 µU.ml-1), registando um valor intermédio (14,63 µU.ml-1) em 17PB e manteve-se constante ao longo da lactação. O Quadro 4 mostra que a concentração sérica de INS aumentou significativamente (P<0,05) ao longo dos dias de amostragem, sendo superior às 24:00h para os três tratamentos. Porém, foi no tratamento 25PB que se registou o valor de concentração mais elevado (25,72 µU.ml-1) (Figura 2A), bastante superior ao verificado para os tratamentos 17PB e 21PB, respetivamente 17,16 e 16,12 µU.ml-1.
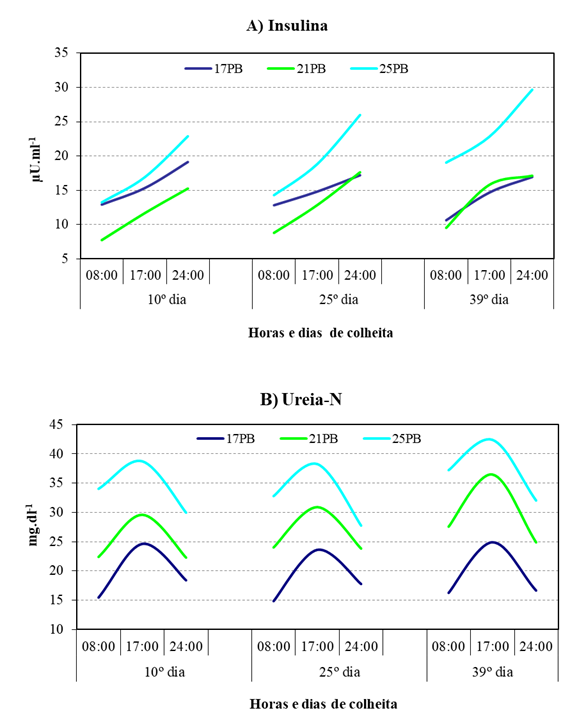
(Nota: a distribuição de alimento foi: erva às 9:00h e suplemento às 18:00h, após retirar a erva não consumida às 17:30h.)
As concentrações séricas de BHBA não evidenciaram diferenças significativas para qualquer dos tratamentos. Ao longo da lactação observou-se uma diminuição significativa (P<0,05) dos valores médios obtidos para os três tratamentos, variando entre 0,770 mmol.l-1 ao 10º dia e 0,591 mmol.l-1 ao 39º dia (quadro 3). Durante o dia a variação foi diminuta, com tendência para aumentar na determinação às 17:00h (Quadro 4).
A concentração dos LT registou valores significativamente superiores (P<0,05) para a dieta 17PB, 2,31 vs 2,07 e 2,05 g.l-1 para as dietas 21PB e 25PB respetivamente. Também os valores de LT no 10º dia de lactação foram significativamente superiores (P<0,05), quando comparados com os registados no 39º dia (Quadro 3). O quadro 4 mostra que os valores diários da concentração de LT foram significativamente superiores (P<0,05) na determinação das 8:00h nos três tratamentos, destacando-se o valor 2,44 g.l-1 verificado nas ovelhas em 17PB.
Os valores médios da concentração sérica de PT das ovelhas no nível 25PB (77,40 g.l-1) foram significativamente superiores (P<0,05) em relação aos das ovelhas em 17PB (70,88 g.l-1), tendo as ovelhas em 21PB registado um valor intermédio (74,04 g.l-1). Não se verificaram variações ao longo da lactação, nem entre horas de amostragem para qualquer dos tratamentos (Quadros 3 e 4).
O quadro 3 mostra que a concentração de Ureia-N variou significativamente (P<0,05) entre os tratamentos, registando-se as médias de 19,17, 26,84 e 34,76 mg.dl-1 em 17PB, 21PB e 25PB respetivamente. Os valores da concentração também foram significativamente superiores (P<0,05) no 39º dia de lactação (28,69 mg.dl-1), em relação aos dos outros dois dias de amostragem, em que foram semelhantes. O quadro 4 refere que a concentração média obtida às 17:00h foi significativamente superior (P<0,05) para qualquer dos tratamentos e dias de lactação, mas com valores médios mais elevados nos grupos 21PB (32,30 mg.dl-1 ) e 25PB (39,83 mg.dl-1) do que em 17PB (24,45 mg.dl-1 ). Todavia, enquanto que, para o tratamento 21PB as concentrações foram semelhantes às 24:00 e 8:00h, respetivamente 23,07 e 24,62 mg.dl-1, para o tratamento 25PB a concentração registada às 8:00 h (34,68 mg.dl-1) foi bastante superior à obtida às 24:00 h (29,50 mg.dl-1) (Figura 2B).
Para os parâmetros TRIG, CREAT e T3 não se registaram diferenças significativas entre tratamentos e dias de amostragem (Quadro 3). A variação das concentrações diárias mostra um padrão comum para os três tratamentos, mas distinto entre os três parâmetros, concentrações superiores na colheita das 8:00 h nos TRIG e na colheita das 17:00 h no caso da CREAT e da T3 (Quadro 4).
DISCUSSÃO
A ingestão de MS de erva das ovelhas suplementadas com MAND (0,9 Mcal EM.dia-1), grupo 17PB, foi em média de 1,133 kg.dia-1 (54,05 g.dia-1/kg0,75 à 6ªsemana), semelhante ao verificado em ovelhas suplementadas com milho (1,072 kg.dia-1; 57,5 g/kg 0,75 à 6ªsemana) num estudo de protocolo semelhante (Belo, 2000), em que a suplementação energética foi de 1,33 Mcal EM.dia-1 e se pretendia garantir a passagem de algum amido ao intestino delgado, pela sua menor degradabilidade ruminal (Owens et al., 1986). No presente estudo, sendo o amido da MAND maioritariamente utilizado ao nível do rúmen (Caldas et al., 2007; Nocek & Tamminga, 1991), a síntese de proteína microbiana terá sido potenciada, aumentando o seu fluxo para o intestino delgado e o seu contributo para a formação da proteína do leite (Poore et al., 1993).
O consumo total de MS de todos os tratamentos foi inferior aos valores referidos em NRC (2007), 2,2 e 2,5 kg de MS em dietas com 14,1 a 15,9% de PB, para ovelhas com 55kg de PV a amamentarem 1 ou 2 borrego respetivamente. Uma das causas desta diferença terá sido o menor tempo disponível para a ingestão da erva (8:30h), comum nos dois estudos que vimos referindo, mas não justificativo da menor ingestão total, comum às suplementações com amido, e ao facto de a ingestão das ovelhas com este tipo de suplementação não ter aumentado ao longo das seis semanas.
A suplementação com amido, visando o aumento da densidade energética das dietas, tem sido objeto de muita investigação, sobretudo em vacas leiteiras, mas leva à diminuição do pH do rúmen (Mould & Orskov, 1983) e à necessidade de administração de fibra para evitar perturbações digestivas (Giger-Reverdin & Sauvant, 1991). Abijaoudé et al. (2000), num estudo com cabras leiteiras, afirmaram que, apesar do aumento da acidez ruminal, o amido de maior degradabilidade aumenta a ingestão e a produção de leite. Aliás, a dieta 17PB com um teor médio de 38,4% de NDF, está de acordo com os estudos de Avondo & Cannas (2001) que indicam os 37% de NDF para a obtenção de boas ingestões de MS e de produção de leite, também cumprindo as recomendações do NRC (2001), que sugere dietas em que 75% do NDF seja proveniente de forragem, para que seja assegurado um bom funcionamento do rúmen. Assim, a menor ingestão de MS das ovelhas suplementadas com amido pode estar relacionada com o fornecimento do suplemento numa única refeição, o que pode ter potenciado o decréscimo do pH do rúmen, ocasionando uma menor digestibilidade da fibra da erva (Bachman, 1992) e a consequente quebra da ingestão.
Os valores de AGNE para os três tratamentos, e o seu escalonamento ao longo da lactação, estão de acordo com a avaliação da CC, apenas com algum significado nas primeiras três semanas, em que se verificaram quebras maiores de CC em 21PB e 17PB. Também em Belo (2000), o teor médio diário de AGNE foi mais relevante ao 10ºdia de lactação, superior para as ovelhas suplementadas com “glúten de milho” (CGF) em relação às suplementadas com milho, respetivamente 0,686 e 0,499 mmol l-1 (com teores de 1,041 e 0,804 mmol l-1 às 8:00h do 10ºdia), ovelhas que também perderam mais CC entre o parto e o 21ºdia de lactação, -0,4 vs -0,31 pontos.
A regulação homeorrética da lactação privilegia a partição dos nutrientes para a glândula mamária, alterando a capacidade relativa dos tecidos corporais para responder aos fatores homeostáticos, como a insulina (Bauman & Currie, 1980; Bines & Hart, 1981), tendo Bauman & Elliot (1983) afirmado que as baixas concentrações de insulina reduzem a sua captação pelos tecidos muscular e adiposo, facilitando o aumento da utilização da glucose pela glândula mamária, que é independente da concentração de insulina. Laarveld et al. (1981) sugeriram que, na regulação do metabolismo da glândula mamária, a disponibilidade de substratos é mais importante do que a influência hormonal e que, com altos níveis de ingestão de nutrientes, as necessidades da lactação podem ser satisfeitas enquanto alguma proporção de energia é retida como gordura corporal. Por sua vez, Metcalf & Weekes (1990) verificaram que, em ovelhas lactantes alimentadas ad libitum e em aumento de peso, existe um ajustamento na partição de nutrientes, estimulado pela insulina, com um aumento da sensibilidade da utilização de glucose por outros tecidos corporais, além da glândula mamária.
Em Belo (2000) foi possível observar que ovelhas leiteiras da mesma raça suplementadas com milho ou CGF, apresentaram concentrações médias de INS, medidas nos primeiros 25 dias de lactação de, respetivamente 18,32 μUI ml-1 e de 12,26 μUI ml-1, resultados que, de acordo com Sauvant (1994), podem ser a reação aos níveis de glucose no intestino, provenientes de substratos de menor degradação no rúmen, tendo também algum significado as diferenças entre as concentrações médias de INS, medidas no mesmo período, nas ovelhas suplementadas com milho com produção leiteira de 1300 ml.d-1 e 930 ml.d-1, respetivamente 11,48 vs 18,44 μUI ml-1. Estes processos metabólicos são descritos na literatura como tendo lugar em fases mais avançados da lactação, quando as ovelhas aproveitando o excesso de energia fornecido, em relação às necessidades da produção leiteira, a utilizam para reporem as suas reservas corporais. Contudo, tal como referiram Cannas et al. (1998), em ovelhas de raças menos apuradas para a produção de leite, os factos indicados poderão ocorrer mais cedo, quando são utilizadas dietas com maiores teores de hidratos de carbono não provenientes de forragens. Neste estudo, os níveis médios de INS foram sempre superiores e crescentes ao longo da lactação nas ovelhas com 25PB, atingindo a concentração média das três amostragens diárias de 23,86 µU.ml-1 no 39º dia, enquanto as ovelhas nos outros dois tratamentos apresentavam um valor médio de 14,14 µU.ml-1. Aliás, a concentração máxima de INS no tratamento 25PB, 29,67 µU.ml-1, foi atingida na colheita das 24:00h do 39ºdia, em que as amostragens para os tratamentos 17PB e 21PB, registaram valores de 16,98 e 17,16 µU.ml-1, respetivamente. Os teores de INS referidos para as ovelhas do tratamento 25PB, apreciados em conjunto com as concentrações de Ureia-N obtidas para o mesmo tratamento, indicam que uma parte substancial da proteína do CGM, que escapou à digestão ruminal, foi utilizada na gluconeogénese hepática. Neste estudo, tal como em Belo (2000), os níveis de INS mostram que alguns substratos podem ter sido desviados para a reposição das reservas corporais, como indicou Van Knegsel et al. (2007) em estudos com vacas leiteiras. Os autores referiram ser a insulina plasmática um intermediário na alteração da partição de energia no início da lactação, acrescentando que a alimentação com dietas ricas em nutrientes glucogénicos resultam num balanço energético negativo menor, devido aos efeitos anabólicos resultantes da alta concentração de INS. Todavia, a menor produção de leite das ovelhas sujeitas ao tratamento 25PB pode também estar relacionada com o custo energético da síntese da ureia, avaliado por Cannas (2004) como sendo equivalente à energia necessária para a produção de 200 g de leite por cada 100 g de PB em excesso das necessidades das ovelhas.
A erva consumida neste estudo teve um conteúdo proteico de 21,6% na MS (3,6 g N.100 g MS-1), valor bem acima dos 2,5-2,7 g N.100 g MS-1 sugeridos por Beever et al. (1986) como valores máximos para limitar a extensa proteólise que ocorre no rúmen quando a erva é a fonte proteica disponibilizada. Mais tarde, Tamminga (1996) indicou que a eficiência da utilização do N no rúmen foi a estratégia mais efetiva para reduzir as suas perdas, seja por redução da degradação da proteína no rúmen ou por um aumento da sua utilização pelos microrganismos ruminais e Bach et al. (2005) indicaram como um objetivo razoável, o fornecimento de 42 g de N disponível por kg de matéria orgânica fermentada no rúmen.
O excesso de PB na alimentação de vacas leiteiras tem sido difícil de identificar pois as necessidades em PB variam com a quantidade de leite produzida, a qualidade das fontes proteicas e a ingestão e composição da MS. Num estudo com vacas leiteiras alimentadas com silagem e alimentos concentrados e sem limitação de hidratos de carbono fermentescíveis, Law et al. (2009) verificaram aumentos significativos nas concentrações séricas de ureia e de proteína total (PT), com o aumento do conteúdo proteico da dieta, indicando os níveis de ureia, um aumento de detoxificação da amónia no fígado e a PT a absorção intestinal da proteína. Estes autores concluíram que, no início da lactação, um aumento da proteína até 173 g.kg MS-1 foi benéfico para a ingestão de MS e para a quantidade de leite produzida, contudo, a eficiência da utilização do N para a produção de leite foi reduzida, como indicam os níveis crescentes de ureia plasmática: 1,56; 2,59 e 4,32 mmol.l-1, para os níveis de proteína de 114, 144 e 173 g.kg MS-1 respetivamente. Reynal & Broderick (2005), também com vacas leiteiras alimentadas com dietas compostas por 50% de alimentos fibrosos (silagem de milho e de luzerna) e 50% de alimentos concentrados, ao testarem níveis de proteína degradável no rúmen (RDP), na procura da eficiência da utilização do alimento sem comprometer a produção de leite e reduzindo as perdas ambientais de N, concluíram que um decréscimo do RDP de 13,2 para 11,7% e a PB da dieta de 18,8 para 17% não afetou a produção de leite mas reduziu a excreção ambiental da Ureia-N em 20%. Porém, em trabalhos incluindo erva na alimentação de vacas leiteiras, salienta-se as investigações de Hoffman et al. (1993) e de Berzaghi et al. (1996) que indicavam que as pastagens das regiões temperadas frequentemente continham mais do que 25% de PB e 20% de RDP, valores bem acima das necessidades das vacas leiteiras de alta produção (NRC, 2001).
Relativamente à utilização de proteína de menor degradabilidade no rúmen (RUP), Hongerholt & Muller (1998), que disponibilizaram dietas com 20% PB e níveis de RUP de 8,4 e 7% fornecidos pelo CGM ou pelo bagaço de soja, respetivamente, a vacas leiteiras, concluíram que a inclusão de RUP na suplementação teve uma menor influência no aumento da quantidade de leite em vacas conduzidas em pastoreio rotacional de Dactylis glomerata. Contudo, na primeira rotação do pastoreio, em que registaram concentrações séricas de 22,3 mg.dl-1 de Ureia-N e de 484 µeq. de NEFA, as vacas multíparas mostraram uma maior tendência para um aumento da quantidade de leite e de proteína com a mistura de concentrado com a maior percentagem de RUP, salientando que a deficiência em energia das dietas foi mais importante do que a possível falta de alguns AA específicos para absorção no intestino delgado. Mais tarde, McCormick et al. (1999), substituíram parte do bagaço de soja por CGM e farinha de sangue tendo verificado que, em dietas com 22% de PB, conseguiam um aumento na quantidade de leite no início da lactação que, todavia, não persistia nas fases seguintes. Como as vacas foram alimentadas com Lolium multiflorum (40% da ingestão) e com silagem de milho, no estudo seguinte McCormick et al. (2001) incluíram um tratamento com alimentação exclusiva em pastoreio e com suplementos de RUP fornecidos em duas proporções. Os resultados mostraram que os aumentos de RUP melhoraram a produção leiteira das vacas no início da lactação.
Num estudo com ovelhas leiteiras, iniciado após os 100 dias de lactação, Mikolayunas-Sandrock et al. (2009), formularam três dietas isoenergéticas com “cubos” de luzerna e Phleum pratense (60% da MS), variando nas concentrações de RDP e RUP, respetivamente 12%-4%; 12%-6% e 14%-4%. As dietas 12-6 e 12-4 apresentaram uma eficiência de utilização do N semelhante, mas as ovelhas alimentadas com a dieta 12-6 produziram mais leite, gordura e proteína, em comparação com a dieta 12-4. O teor da ureia-N no leite (MUN) revelou-se mais relacionado com a concentração da PB do que com a sua degradabilidade, verificando-se os valores mais elevados em 12-6 e 14-4 (média de 26,86 mg.dl-1). Mikolayunas et al. (2011) também estudaram o efeito da suplementação com RUP em ovelhas leiteiras (após o 100º dia de lactação) alimentadas à manjedoura com diferentes proporções de Dactylis glomerata (11,7% PB) e luzerna (19,4% PB), e com um alimento concentrado, incluindo ou não uma fonte de RUP. A suplementação com RUP não aumentou a ingestão de MS nem a quantidade de leite e de proteína obtidos, facto que os autores atribuíram às menores necessidades das ovelhas, devido à fase adiantada da lactação ou, à menor quantidade de RUP fornecida, estimada em 5,3 e 3,7% da MS. A suplementação não afetou a eficiência de utilização do N e o MUN foi significativamente superior nas ovelhas suplementadas com RUP.
Num outro estudo com ovelhas leiteiras em pastoreio de azevém, iniciado após o 60º dia de lactação, Wilkinson et al. (2000) testaram seis alimentos concentrados isoenergéticos que forneceram três níveis de RDP e dois de RUP digestível (70 e 45 g.kg MS-1) e em que a PB total dos tratamentos variou entre 247 e 180 g.kg MS-1. Os autores observaram que, com o aumento dos níveis de RDP, obtiveram uma maior quantidade de leite e das concentrações de proteína e lactose que atribuíram a um aumento da síntese de proteína microbiana e de proteína metabolizável, o que terá resultado numa maior proporção de aminoácidos que foram desaminados e utilizados como precursores para a síntese da lactose.
As vantagens produtivas da utilização de fontes proteicas de baixa degradabilidade ruminal foram evidenciadas anteriormente em ovelhas em pastoreio por Penning et al. (1988) que estudaram a resposta produtiva à suplementação proteica e obtiveram resultados positivos na produção leiteira e no ganho médio diário de borregos, ao utilizarem farinha de peixe. O mesmo foi verificado por Purroy e Jaime (1995), ao comparar duas fontes de proteína (bagaço de soja e farinha de peixe) em dietas com 20% de PB, tendo encontrado uma resposta superior nas ovelhas suplementadas com farinha de peixe.
No presente estudo, a produção leiteira, superior nas ovelhas do tratamento 21PB, mostrou a vantagem da suplementação no início da lactação com proteína de baixa degradabilidade ruminal. As concentrações séricas médias de Ureia-N para os tratamentos 21PB e 25PB, respetivamente 26,84 e 34,76 mg.dl-1 são semelhantes às obtidas por Belo (2000), em que ovelhas na mesma fase da lactação, alimentadas com uma dieta à base de erva, suplementadas com milho (dieta com 20% de PB) e CGF (dieta com 24,5 % de PB), apresentaram teores semelhantes de Ureia-N, respetivamente 24,79 e 32,87 mg.dl-1. A diferença nos teores de Ureia-N entre os tratamentos 21PB e 25PB, corresponde em parte à menor quantidade de MAND fornecida, mas também à proteína proveniente do CGM, uma vez que as ovelhas tiveram níveis de ingestão de erva muito semelhantes. Também a variação diurna entre os tratamentos é indicativa, pois, enquanto no tratamento 21PB as concentrações de Ureia-N às 24:00 e às 8:00h do dia seguinte foram semelhantes (23,61 e 24,62 mg.dl-1, respetivamente), no tratamento 25PB aumentaram nesse período, de 29,80 para 34,67 mg dl-1 .
Se, simultaneamente com aqueles valores se considerarem os teores séricos médios da PT para as ovelhas em 17PB, 21PB e 25PB, respetivamente 70,88 g.l-1, 74,04 g.l-1 e 77,40 g.l-1, verificamos a influência do fornecimento de CGM. Enquanto a passagem da dieta com 17PB para a dieta contendo 21PB originou um aumento de ingestão e de produção leiteira, o aumento do nível de CGM de 21PB para 25PB conduziu a um decréscimo de produção de leite. Os aumentos das concentrações de Ureia-N referem-se ao aumento da ingestão de erva e ao decréscimo da quantidade de MAND na ração, acentuada no tratamento 25PB, que não permitiu a presença de glúcidos em quantidade suficiente para a utilização do NH3 produzido no rúmen. O aumento da concentração de Ureia-N acentuou-se nas ovelhas sob o tratamento 25PB e estará ligado ao incremento da taxa de gluconeogénese da proteína do CGM que escapou à digestão ruminal. O aumento da gluconeogénese entre os tratamentos 21PB e 25PB, originou um excesso de energia disponível que, como indicam os teores de insulina ao 39º dia de lactação, respetivamente 29,67 vs 17,16 µU.ml-1 (o 2º valor é próximo do registado para as ovelhas 17PB), pode ter sido desviada para a reposição das reservas lipídicas. Às ovelhas do grupo 25PB foi proporcionado um excesso de energia não aproveitado para a produção de leite que também foi prejudicada pelo excesso de Ureia-N que, ao ser metabolizado, teve custos energéticos que contribuíram para o decréscimo de produção observado.
A suplementação de ovelhas leiteiras conduzidas em pastoreio tem como principal condicionante o teor de proteína da pastagem, tanto maior quanto mais jovem for a erva e maior a proporção de leguminosas, fatores essenciais para promover uma maior ingestão de MS deste alimento. A suplementação deverá privilegiar o fornecimento de glúcidos de diferente degradabilidade ruminal, tentando sincronizar a disponibilidade em energia com o aumento do NH3 proveniente da degradação da proteína da erva, já que o acesso ao suplemento, contrariamente ao que ocorre com dietas completas, em ovelhas em pastoreio está frequentemente condicionado aos tempos de ordenha. A escolha do alimento proteico de menor degradabilidade ruminal é essencial no início da lactação ou, nas fases seguintes, para as ovelhas de maior produção leiteira, ajudando a incrementar a quantidade de proteína que fica disponível para a síntese da proteína do leite ou para o apoio à gluconeogénese. Neste estudo, considerando que a RUP da erva representa 30% da sua PB (Hongerholt & Muller, 1998) e a do CGM atinge os 69% da PB do alimento (NRC, 2001), as ovelhas nas dietas 17PB e 21PB tiveram acesso a cerca de 5 e 9% de RUP na MS ingerida. Na literatura não se encontram ensaios que refiram os efeitos de concentrados com RUP em ovelhas nesta fase da lactação e conduzidas em pastoreio, mas os teores de Ureia-N indicados acima, levam-nos a pensar que, o desempenho das ovelhas do grupo 21PB poderia ter melhorado se tivessem sido utilizados glúcidos de diferente grau de solubilidade ruminal que, promovendo uma maior utilização do N proveniente da pastagem, poderia também proporcionar um decréscimo na suplementação com CGM.
CONCLUSÃO
O estudo de uma fonte proteica de menor degradabilidade ruminal visou contribuir para o conhecimento do processo metabólico, relacionado com a utilização da energia proveniente da mobilização da gordura corporal, importante no início da lactação.
A utilização de CGM na composição do suplemento na proporção inferior (78 g de MS, tratamento 21PB), conduziu a uma ingestão de erva de 72,28 g.dia-1 por kg PV0,75 e à produção de 683,1 ml de leite ordenhado.ovelha.dia-1, com os teores de 3,58%, 5,04%, 5,04% de TG, TP e TL respetivamente. Os borregos apresentaram um PV de 11,81 kg aos 42 dias de idade.
Durante as seis semanas as ovelhas do tratamento 21PB perderam 3,56 kg de PV e mobilizaram 0,25 pontos de CC, que apenas foi significativa nas primeiras três semanas de lactação,
A determinação dos teores séricos de AGNE, INS, Ureia-N e PT revelaram-se importantes para identificação dos processos metabólicos durante a fase inicial da lactação.

