











 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUÇÃO
Os solos agrícolas são fundamentais na produção de alimentos, e apresentam uma importância cada vez mais vital tendo em conta o aumento da população mundial e as necessidades alimentares crescentes (Silver et al., 2021). A aplicação de lamas das estações de tratamento de águas residuais (ETARs) em solos agrícolas é uma prática que visa compensar a perda de fertilidade dos solos, contribuir para o aumento da matéria orgânica e de nutrientes vegetais para as plantas, e resolver o problema da gestão de resíduos das lamas como um excedente. No entanto, esta aplicação pode levar ao aumento da concentração de diversos contaminantes no solo tais como as substâncias psicoativas (SPAs) e de outros compostos bioativos como fármacos, uma vez que estes compostos podem ser adsorvidos nas lamas (Álvarez-Ruiz et al., 2015). Desta forma, a aplicação de lamas contaminadas nas parcelas agrícolas pode representar uma ameaça para o solo e para os diversos organismos terrestres (Langford, 2011) e as suas funções ecológicas, bem com algum risco para o Homem.
O aumento do consumo de SPAs para fins recreativos tem gerado preocupação porque estes compostos, uma vez consumidos, são excretados na sua forma original ou como metabólitos, chegando às ETARs. Devido às suas propriedades biológicas, as SPAs podem causar efeitos adversos nos organismos expostos (Carter et al., 2020; Ribeiro et al., 2023), sendo considerados poluentes pseudopersistentes (Gualano et al., 2014) mas podem também afetar diversos serviços do ecossistema envolvidos mesmo em concentrações muito baixas (Santana-Viera et al., 2023).
A metafedrona (3-MMC) é uma catinona sintética pertencente à classe das feniletilaminas, com efeitos psicoestimulantes (Ferreira et al., 2019). Este contaminante emergente chega aos ecossistemas aquáticos através da excreção pela urina ou descarte inapropriado, e consequentemente aos solos (Subedi & Kannan, 2015) podendo causar efeitos adversos nos organismos edáficos e na ecologia do solo.
As minhocas são organismos importantes para a estrutura, fertilidade e ecologia do solo, sendo facilmente afetadas pelos diversos tipos de contaminação do solo. Deste modo, as minhocas da espécie Eisenia fetida e Eisenia andrei apresentam uma série de vantagens como bioindicadores relevantes sendo recomendados nos ensaios ecotoxicológicos padronizados pelas entidades internacionais (OECD, 1984).
Apesar da crescente contaminação dos ecossistemas com SPAs (Santana-Viera et al., 2023), a avaliação dos efeitos potenciais destas substâncias nos organismos do solo é escassa. Assim, o principal objetivo deste trabalho foi estudar os potenciais efeitos da 3-MMC no comportamento de fuga e reprodução da E. fetida.
MATERIAIS E MÉTODOS
A 3-MMC foi adquirida na LGC Standards (Wesel, Alemanha). Para a realização dos ensaios foram seguidas as diretrizes para o estudo de substâncias químicas usando o ensaio de fuga (ISO, 2008) e o ensaio de reprodução (ISO, 2012; OECD, 2016). Para isso, foi preparado um solo artificial constituído por 70 % de areia de silício (0,7 mm, Axton), 20% de caulino em pó (MIBAL, Portugal) e 10 % de turfa seca (SIRO 100), com base no peso seco (ps). Posteriormente, foi determinada a capacidade máxima de retenção de água (CMRA) e o valor de humidade foi ajustado para valores de 40% da CMRA. Adicionalmente, o valor de pH inicial do solo artificial foi corrigido para valores de 6,0 ± 0,5 através da adição de CaCO3 (Labsolve, Portugal).
Antes do início dos ensaios, foram pesadas e selecionadas cerca de 300 minhocas adultas da espécie E. fetida (com clitelo bem desenvolvido) com biomassa compreendida entre as 300 e 600 mg, e colocadas em solo artificial durante 24 a 48 horas para se aclimatarem. Para os ensaios de fuga e de reprodução, foram testadas três concentrações de 3-MMC, 25, 250 e 2500 µg kg-1 (de forma a incluir as concentrações encontradas nas águas de superfície e residuais (Mwenesongole et al., 2013; Gao et al., 2017)) e um controlo, sendo realizadas três replicas por controlo e cada concentração estudada.
De acordo como design experimental, foram utilizadas 12 caixas de PVC, como recipiente-teste para cada ensaio (controlo, exposição), com as respetivas tampas devidamente furadas, para possibilitar as trocas gasosas, mas impedir a saída das minhocas. Os ensaios decorreram sob condições laboratoriais constantes de temperatura (20±2 ºC), fotoperíodo (16h luz/8h escuro), intensidade de luz, arejamento e humidade.
Para a realização do ensaio de fuga (Figura 1), foi colocada uma placa divisória (separador) a meio de cada recipiente-teste, e posteriormente cada secção foi preenchida com 250 g de solo artificial (equivalente a ps) a 40% da CMRA, ao qual foi adicionado 3-MMC nas diferentes concentrações selecionadas e um solo controlo. Depois da remoção da placa divisória, foram inseridas 10 minhocas na linha central entre as duas secções de solo e deixadas a incubar durante 48 horas. Posteriormente, a placa divisória foi inserida novamente na linha central, procedendo-se à contagem do número de organismos em cada secção, de forma a determinar a percentagem de fuga (𝓍) de acordo com a seguinte equação:
em que n c representa o número de minhocas presentes no solo controlo, n t representa o número de minhocas presentes no solo teste e N o número total de minhocas por recipiente-teste.
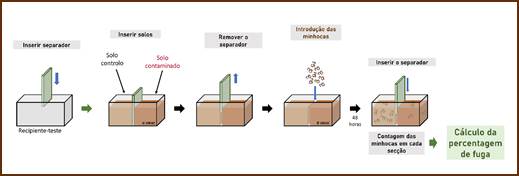
Figura 1 Esquema do delineamento experimental do ensaio de comportamento realizado com minhocas adultas (autores Ondina Ribeiro e João Soares Carrola, adaptado de Stander et al. (2019)).
Para o ensaio de reprodução (Figura 2) foram colocadas 500 g de solo (equivalente a ps), a 40% de CMRA com a respetiva concentração de 3-MMC. Também foi realizado um tratamento controlo, sem a adição de 3-MMC ao solo. De seguida, foram colocadas 10 minhocas adultas em cada recipiente-teste, previamente aclimatadas e depuradas, e adicionou-se 1 g de aveia biológica moída e previamente esterilizada numa câmara de luz ultravioleta. A cada 7 dias, procedeu-se ao controlo gravimétrico do teor de humidade e à alimentação. Após 28 dias de exposição, as minhocas adultas foram retiradas dos recipientes-teste e os casulos foram mantidos por mais 28 dias, ao fim dos quais foi realizada a contagem dos juvenis eclodidos em cada tratamento e replica.
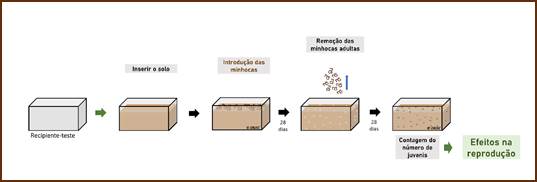
Figura 2 Esquema do delineamento experimental do ensaio de reprodução realizado com minhoca (autores Ondina Ribeiro e João Soares Carrola).
Os resultados obtidos no ensaio de fuga foram analisados pelo teste exato de Fisher, como descrito em Natal-da-Luz et al. (2004). Este procedimento estatístico permite comparar a distribuição dos organismos em relação a uma distribuição esperada assumindo a inexistência de uma resposta de fuga a um determinado tipo de solo (hipótese nula). Foi utilizado um teste unicaudal no qual a hipótese nula assume que metade dos organismos permanece no solo que está a ser avaliado, não havendo fuga. Para os testes duais, foi testado um teste bicaudal, no qual a hipótese nula assume uma distribuição igual dos organismos em ambas as secções do recipientes-teste. As hipóteses nulas foram rejeitadas para probabilidades iguais ou inferiores a 0,05.
Os dados da reprodução foram sujeitos a uma análise de normalidade na distribuição e homogeneidade de variâncias, pelos testes de Shapiro-Wilk e Bartlett, respetivamente. Posteriormente, os dados foram sujeitos a uma ANOVA de 1 fator, seguida do teste de Dunnett para um grau de probabilidade de p≤0,05.
RESULTADOS E DISCUSSÃO
Os resultados observados para o ensaio de fuga, relativos à distribuição das minhocas adultas em cada uma das secções, são apresentados na Figura 3. A presença de 3-MMC no solo não induziu um comportamento de fuga nas minhocas para todas as concentrações de substância testadas, uma vez que não foram observadas diferenças significativas entre os resultados do solo controlo vs. solo teste (p > 0,05). Para a concentração de 25 µg kg-1 observou-se uma percentagem de fuga de 40%, enquanto que para as concentrações mais elevadas (250 e 2500 µg kg-1) observou-se uma percentagem de fuga de 6,67% em relação ao controlo. Apesar da percentagem de fuga ser mais elevada na concentração mais baixa, este comportamento não foi significativo (p = 0,084), nem muito evidente ou consistente com o aumento das concentrações.
Relativamente ao ensaio de reprodução verificou-se que as diferentes concentrações de 3-MMC não afetam significativamente (p > 0,05) a capacidade reprodutiva das minhocas à semelhança do ensaio de fuga, tal como apresentado na Figura 4.
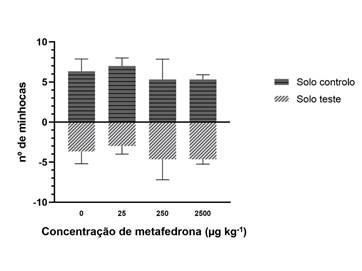
Figura 3 Resultados do ensaio de fuga das minhocas (E. fetida), que mostram o número de minhocas em função da concentração testada (valores representados em forma de médias e desvio-padrão; n=3).
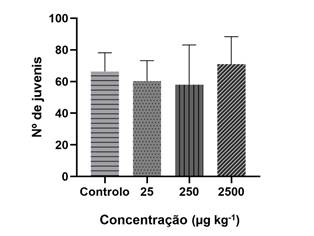
Figura 4 Resultados do ensaio de reprodução com minhocas da espécie E. fetida após 56 dias de exposição de (valores expressos sob a forma de média ± desvio-padrão, n=3).
Analisando os resultados obtidos para os dois ensaios realizados, estes sugerem que a 3-MMC não induz toxicidade na espécie E. fetida nem altera a qualidade da matriz do solo. Carter et al. (2016) observaram que a exposição de minhocas (E. fetida) a diferentes fármacos, como carbamazepina, diclofenac, fluoxetina e orlistato em concentrações de 26, 25, 28 e 44 μgkg-1, respetivamente, não induziram efeitos no comportamento de escavação dos organismos expostos. No entanto, a exposição ao antibiótico lasalocida induziu uma resposta de fuga significativa com o aumento das concentrações testadas em minhocas da espécie E. andrei (Žižek & Zidar, 2013).
Uma vez que o ensaio de fuga é de curta duração (48 horas), isso pode limitar o nível de resposta das minhocas e não ser suficiente para se observar os efeitos adversos da substância testada. Desta forma, foi realizado o ensaio de reprodução (ensaio crónico, 28 + 28 dias) para possível avaliação do risco uma vez que a 3-MMC fica mais tempo biodisponível. No entanto, este maior tempo de exposição não foi suficiente para se detetar efeitos tóxicos da 3-MMC na reprodução desta espécie.
Apesar desta SPA poder induzir efeitos adversos em organismos aquáticos (Ferreira et al., 2019; Ribeiro et al., 2023), a sorção de SPAs não depende apenas da sua lipofilicidade mas também da solubilidade, temperatura, pH e conteúdo da matéria orgânica do solo (Santana-Viera et al., 2023), o que pode alterar a sua toxicidade nos organismos do solo. Assim, a ausência da toxicidade da 3-MMC presente no solo pode estar relacionada com os fenómenos de adsorção e complexação que os constituintes do solo (matéria orgânica e argilas) exercem no composto. Pode ainda existir um efeito tampão que diminui a biodisponibilidade da 3-MMC (Silva & van Gestel, 2009) e mitiga os efeitos adversos nos organismos expostos.
CONCLUSÕES
Os estudos sobre os efeitos de SPAs em minhocas são escassos e até ao momento não foi encontrado nenhum trabalho que avalie o efeito tóxico da 3-MMC na espécie E. fetida o que mostra a importância e inovação deste trabalho.
Os resultados obtidos mostram que a 3-MMC não induz efeitos tóxicos na espécie E. fetida, para a gama de concentrações testadas (25, 250 e 2500 µg kg-1). Estes dados preliminares indicam que a 3-MMC não interfere no comportamento de evitamento nem na reprodução da E. fetida mesmo na concentração mais alta.
No entanto, é importante realizar mais estudos para complementar estes dados, desde avaliar a possível degradação destas substâncias no solo, estudar os efeitos desta substância isolada ou de misturas de várias SPAs nas minhocas e também noutros invertebrados edáficos (com papel relevante na ecologia dos solos agrícolas) bem como investigar a sua bioacumulação ao longo da cadeia alimentar.
Assim, torna-se importante a realização de mais estudos para uma avaliação do risco potencial mais completa e rigorosa para as populações de minhocas e de outros organismos terrestres bem como para a saúde do solo.

