











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
La resistencia a herbicidas sintéticos y la contaminación debida al abuso de agroquímicos son problemas actuales que afectan a la agricultura en todo el mundo. Implementar y desarrollar herramientas eficaces y respetuosas con el medio ambiente para el manejo de arvenses es un desafío necesario para lograr una agricultura sostenible. Varios grupos de investigación buscan nuevos compuestos con modos de acción diferentes a los herbicidas convencionales. El metabolismo secundario vegetal es una enorme fuente de moléculas con diferentes estructuras y actividades biológicas.
La trans-chalcona (chalcona) es un flavonoide de demostrada actividad bactericida, antiinflamatoria, antitumoral y antimalárica, entre otras. También es altamente fitotóxica, siendo capaz de inhibir el crecimiento radicular de varias especies de malas hierbas, incluida la especie modelo Arabidopsis thaliana (L.) Heynh., y resultando inocua para los cultivos a los que se encuentran frecuentemente asociadas (Díaz-Tielas et al., 2014). Ha mostrado una concentración inhibitoria del 50% del crecimiento de la raíz (CI50) de Arabidopsis thaliana (Arabidopsis) de tan solo 35 µM, induciendo además muerte celular programada de tipo apoptótico en las raíces tras 14 días de crecimiento en medio con chalcona (Díaz-Tielas et al., 2012). Estos estudios demuestran que la chalcona es una firme candidata para su uso como regulador del crecimiento vegetal, un bioherbicida. En este trabajo se ha llevado a cabo un estudio del perfil metabolómico de Arabidopsis a lo largo del tiempo de exposición a la chalcona (3, 10 y 20 horas) con la intención de determinar los cambios en el metaboloma más relevantes en su modo primario de acción.
MATERIALES Y MÉTODOS
Se sembraron 24 semillas de Arabidopsis thaliana ecotipo Columbia (Col-0) en placas con agar+nutrientes+sacarosa, y se colocaron en posición vertical en cámara de cultivo (22±2ºC, 16 h luz, 120 µmol m-2 s-1). Tras 14 días, las plántulas se trataron con una solución de chalcona (trans-benzalacetofenona, 97%, Sigma-Aldrich) (0 y 35 µM) en H2Od+0.1%EtOH durante 3, 10 y 20 horas. En la cosecha se separó la raíz de la parte aérea, se pulverizó en N2 líquido y se almacenó a -80ºC. Se realizaron cuatro réplicas de cada muestra. Para el análisis metabolómico no dirigido se procedió a la extracción y derivatización de los compuestos presentes en las muestras siguiendo el protocolo descrito por Lisec et al. (2006), empleando metanol y cloroformo para la extracción, y clorhidrato de metoxilamina y N-Metil-N-(trimetilsilil)trifluoroacetamida (MSTFA) para la derivatización. Se añadieron 60 µL de ribitol (0,2 mg/mL) como patrón cuantitativo interno. Para la separación y fragmentación de los metabolitos se empleó un equipo de cromatografía de gases (Agilent 789A GC) acoplado a espectrometría de masas de cuadrupolo único (Agilent 5975C) en una columna capilar MEGA-5MS y precolumna de 10 metros.
Los datos se analizaron con el software MSDIAL usando librerías de impacto electrónico públicas. Tras la extracción de los picos, filtración y calibración de los datos, alineamiento de los picos, análisis de espectros deconvolucionados e identificación e integración de la altura de los picos, se normalizaron los datos de los metabolitos anotados y se eliminaron los que estaban 2 veces por debajo de la media de los blancos. El análisis estadístico se realizó el software MetaboAnalyst 5.0. Los datos se transformaron (log10) y escalaron (Pareto) antes de proceder al análisis con la opción “time-series + 1 factor” para ANOVA de dos factores (post-hoc Tukey’s test, FDR ≤ 0.05), Análisis de Componentes Principales (PCA) y Bayes empírico multivariado (MEBA). También se analizaron las rutas metabólicas más impactadas en cada tiempo de medida con la función “Pathway analysis”.
RESULTADOS Y DISCUSIÓN
El análisis metabolómico permitió identificar y anotar un total de 93 metabolitos en la raíz y 138 en la parte aérea de Arabidopsis. El PCA mostró una buena separación de los grupos por tratamientos y en los distintos tiempos de medida (Figura 1A). Esta separación fue más clara en la raíz, ya que en la parte aérea se observa mayor agrupamiento a medida que el tiempo aumenta. El porcentaje de varianza explicado por los componentes principales (PC) fue mayor en la raíz que en la parte aérea; con un 38.7% (PC1), 11.6% (PC2) y 10.3 % (PC3) para la raíz y un 26.8% (PC1), 13.5% (PC2) y 7.6 % (PC3) para la parte aérea. Los resultados del ANOVA mostraron un total de 76 y 101 metabolitos afectados en raíz y parte aérea respectivamente, indicando que en ambos casos la mayoría de los metabolitos anotados se vieron afectados, bien por el tratamiento con chalcona, por el tiempo, o por la interacción de ambos factores (Figura 1B).
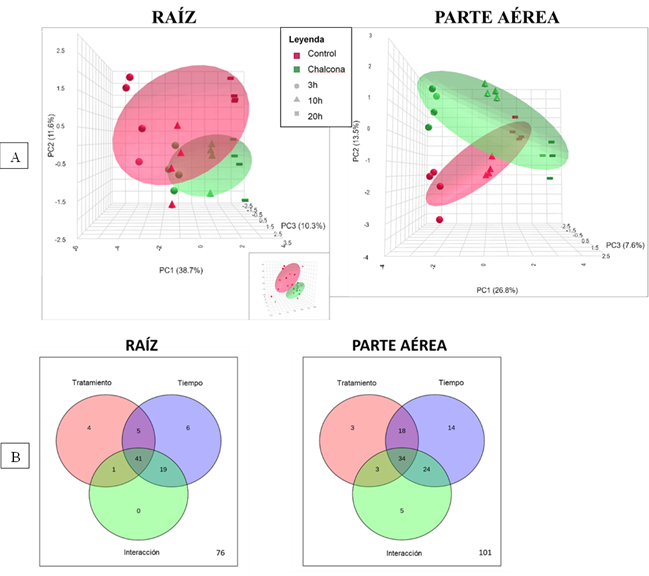
Figura 1 A: Análisis de componentes principales de los metabolitos detectados en la raíz y parte aérea de plántulas de Arabidopsis tratadas con chalcona 0 µM (control) y 35 µM durante 3, 10 y 20 horas. B: Diagramas de Venn mostrando el número de metabolitos significativamente afectados en raíz y parte aérea debido al tratamiento con chalcona, al tiempo de tratamiento y a la interacción de ambos factores (ANOVA, FDR≤0.05).
El análisis de las rutas metabólicas implicadas reveló que las más impactadas son comunes en la raíz y en la parte aérea (Tabla 1). La chalcona impactó notablemente el metabolismo de la alanina, aspartato y glutamato, relacionado también con el GABA shunt. El ácido glutámico y glicina, precursores del glutatión, se vieron afectados de forma distinta en raíz y parte aérea. Mientras que el ácido glutámico disminuyó en la parte aérea, en la raíz se observó un incremento de ambos aminoácidos a las 10 horas de tratamiento, así como un incremento de GABA, indicando una posible activación de mecanismos de tolerancia al estrés. Tanto el GABA shunt como el metabolismo de la L-alanina están muy relacionadas con el ciclo de Krebs. El incremento de L-alanina, asparagina y aspartato; así como la disminución de fumarato, ácido succínico, glutamina, ácido glutámico y GABA (excepto raíz 10h) sugieren una posible reducción del ciclo de Krebs debido al tratamiento con chalcona.
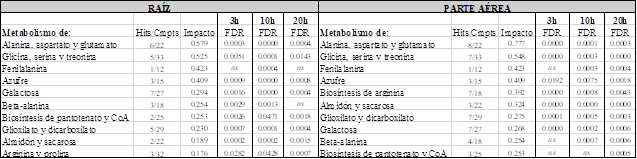
Tabla 1 - Rutas metabólicas más impactadas en la raíz y en la parte aérea de plántulas de Arabidopsis a lo largo del tratamiento con chalcona (3, 10 y 20 horas). FDR: tasa de descubrimiento falso, ns: no significativo
A las 3 h de tratamiento con chalcona se produjo en la parte aérea una disminución de glicina, serina, glicerato, glutamato y glutamina, indicando una posible disminución de la fotorrespiración (Figura 2A). A las 10 y 20 h se produjo un aumento de serina y glicerato por encima del control, sugiriendo una activación de la síntesis de serina por la vía del glicerato, quizás en compensación de la inhibición de la vía fosforilativa. También a las 3 h de tratamiento se detectó un fuerte incremento de prolina en la parte aérea, pudiendo ser una respuesta al estrés inducido por la alteración de la fotorrespiración. En la raíz no se encontraron diferencias significativas en la acumulación de glicerato y glutamina; la serina se acumuló por encima del control a las 3 y 10 h, y la glicina se incrementó a las 10 h pero disminuyó a las 20 h (Figura 2B). Estos resultados sugieren una activación de la biosíntesis de serina por la vía fosforilativa, ya que la del glicolato ocurre solo en los cloroplastos, y la posterior síntesis de glicina a partir de la serina. La serina podría ser un importante mediador en las respuestas de las plantas al estrés ambiental, y varios estudios han mostrado su acumulación ante diversos factores estresantes.
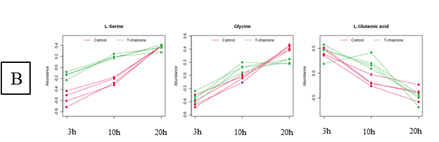
Figura 2 Perfil temporal, obtenido tras análisis MEBA, de metabolitos relacionados con procesos de fotorrespiración en (A) la parte aérea y (B) la raíz de plántulas de Arabidopsis tratadas con chalcona 0 µM (control) y 35 µM durante 3, 10 y 20 horas. Las líneas de color rojo indican el control y las de color verde el tratamiento.
Para determinar qué metabolitos son los que más se vieron afectados por el tratamiento a lo largo del tiempo se realizó el análisis MEBA, que compara los perfiles de cada variable a lo largo del tiempo entre dos condiciones de estudio (chalcona 0 y 35 μM) aportando una lista ordenada de los metabolitos que más varían de acuerdo con estos dos factores en función del valor T2 de Hotelling obtenido. Así, los metabolitos con mayor valor T2 de Hotelling son los que más varían a lo largo del tiempo y en función del tratamiento con chalcona (Tabla 2). Entre ellos abundan los relacionados con metabolismo de fenoles, como el pirogalol, el catecol, la hidroquinona y el ácido tereftálico y el ácido protocatéquico. Todos ellos se encontraron reducidos en las plantas tratadas, con excepción del ácido protocatéquico, que aumentó en la parte aérea a las 3 y 10 h. Además, en las raíces se encontró un aumento del ácido p-hidroxibenzoico a las 3 y 10 h, y un aumento del ácido benzoico a las 10 h de tratamiento.
Tabla 2 Valores de T2 de Hotelling de los 10 metabolitos que más varían a lo largo del tiempo entre el control y el tratamiento con chalcona en la raíz y parte aérea de Arabidopsis
| Raíz | Parte aérea | |||
|---|---|---|---|---|
| Metabolitos | Hotelling-T2 | Metabolitos | Hotelling-T2 | |
| Ácido dehidroabiético | 356.6 | Ácido protocatéquico | 477.5 | |
| Asparagina | 261.1 | Glucosa 1-fosfato | 366.9 | |
| Ácido 6-hidroxinicotínico | 149.2 | L-prolina | 214.4 | |
| L-alanina | 128.8 | L-alanina | 151.9 | |
| L-lisina | 121.4 | Ácido dehidroabiético | 137.0 | |
| Ácido tereftálico | 121.1 | Sacarosa | 109.8 | |
| Hidroquinona | 115.2 | L-ácido glutámico | 107.5 | |
| Glucuronato | 101.6 | Catecol | 105.9 | |
| Sarcosina | 98.5 | Sinigrina | 94.5 | |
| Pirogalol | 98.1 | L-glutamina | 89.5 | |
La biosíntesis del ácido protocatéquico ocurre en los plastidios directamente de intermediarios de la ruta del ácido siquímico, mientras que la biosíntesis de ácidos benzoicos es a partir de intermediarios de la ruta de los fenilpropanoides. El ácido siquímico se redujo y la fenilalanina aumentó en la parte aérea en los tres tiempos (Figura 3A). En la raíz la fenilalanina aumentó a las 10 h, y el ácido siquímico aumentó a las 3 h y disminuyó drásticamente a las 20 h, al igual que el ácido p-hidroxibenzoico (Figura 3B). Estos resultados sugieren una activación de la ruta de los fenilpropanoides en la raíz y de la ruta del siquimato en la parte aérea (en detrimento de la ruta de los fenilpropanoides). Esto es consistente con la inhibición la enzima fenilalanina amoniaco liasa (PAL) (Chen et al., 2011) en la parte aérea y con la inhibición de la 4-cumarato-CoA ligasa (4CL) (Yun et al., 2005) en la raíz. De nuevo, el modo de acción de la chalcona se muestra diferente en las raíces y en la parte aérea de plántulas de Arabidopsis.
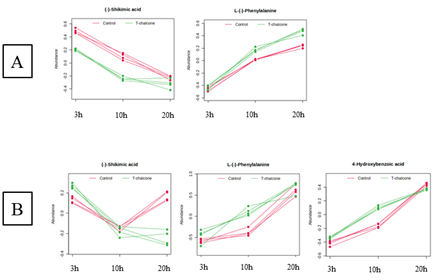
Figura 3 Perfil temporal, obtenido tras análisis MEBA, de metabolitos relacionados con las rutas del ácido siquímico y de los fenilpropanoides en (A) la parte aérea y (B) la raíz de plántulas de Arabidopsis tratadas con chalcona 0 µM (control) y 35 µM durante 3, 10 y 20 horas. Las líneas de color rojo indican el control y las de color verde el tratamiento.
CONCLUSIONES
Las alteraciones en la acumulación de metabolitos son diferentes en la raíz y en la parte aérea de plántulas de Arabidopsis thaliana ya a las 3 horas de tratamiento con chalcona, indicando distintos modos de acción en cada órgano. Aunque las alteraciones metabólicas detectadas concuerdan con la inhibición de los enzimas PAL y 4CL, éstos deben estar implicados en el modo de acción de la chalcona, pero no son su diana molecular. La chalcona afecta de manera determinante al metabolismo del carbono y aminoácidos, y es posible que su diana molecular se encuentre en rutas del metabolismo central, especialmente las relacionadas con el ciclo de Krebs y la fotorrespiración, si bien son necesarios más estudios para hallarla.

