











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Amaranthus palmeri, S. Watson (A. palmeri) es una especie de maleza invasora peligrosa, que puede reducir drásticamente la producción de cultivos gracias a su rápido crecimiento, adaptabilidad y producción prolífica de semillas (Ehleringer, 1983). A. palmeri puede acumular rasgos de resistencia a múltiples modo de acción herbicidas, entre ellos los inhibidores de la ALS y de la EPSPS (Küpper et al., 2017). Los inhibidores de ALS son una clase de herbicidas que inhiben la acetolactato sintasa, una enzima esencial en la biosíntesis de los aminoácidos de cadena ramificada valina, leucina e isoleucina (Duggleby et al., 2008). Los inhibidores de la EPSPS causan la muerte de las plantas al inhibir la enzima 5-enolpiruvilshikimato-3-fosfato sintasa, presente en la ruta de síntesis de los aminoácidos aromáticos: fenilalanina, tirosina y triptófano (Grube et al., 2019). En el caso de la ALS la resistencia es debida a mutaciones puntuales en los codones A122, P197, A205, D376, R377, W574, S653 and G654 impiden al herbicida de ligarse y interactuar con el sitio activo de la enzima. Por otro lado, la sobreexpresión del gen EPSPS es el mecanismo mayormente reportado en esta especie donde la presencia de un exceso de copias del gen impide que el herbicida inhiba todas ellas de manera efectiva (Powles & Yu, 2010). Debido a la continua introducción de nuevas semillas y a la presión de selección herbicida que las mismas reciben una vez llegadas, es probable que existan poblaciones con resistencia múltiple a estos dos MoA. En este estudio se evalúa la resistencia a los inhibidores de la ALS y ESPSP en dos poblaciones españolas de A. palmeri procedentes de margen de carreteras y se caracteriza la presencia de mecanismo de resistencia múltiple tanto a nivel de población como de plantas.
MATERIALES Y MÉTODOS
Material vegetal
Tres poblaciones de A. palmeri fueron incluidas en este estudio: dos fueron recolectadas en un margen de carretera en Montblanc (MB) y en Tarragona (TA) en el noreste de España, y una de Carolina del Norte, EE. UU., que se utilizó como control sensible (WT). Las semillas se recolectaron de diferentes plantas madre, se limpiaron y se almacenaron a temperatura ambiente hasta que comenzaron los experimentos.
Tratamiento con inhibidores de la ALS
Las distintas poblaciones se sembraron en turba, se colocaron en bandejas de aluminio y se cultivaron en condiciones de invernadero. Después de cinco días, se trasplantaron 20 plántulas por población a bandejas de plástico (325 × 265 × 95 cm³) con un suelo de arcilla, turba y perlita (60%, 20%, 20%) regándolas diariamente. Al estadio de 4-5 hojas se trataron con thifensulfuron-metil, THIF (Harmony 50 SX, DuPont™) e imazamox, IMA (Tuareg®, DuPont™) en dosis recomendadas en campo de 6 g a.i. ha-1 y 40 g a.i. ha-1, respectivamente. El diseño experimental fue completamente aleatorio con dos repeticiones (una bandeja, una repetición). Cuatro semanas después de la aplicación, se determinó el porcentaje de plantas sobrevivientes y se recolectó el material vegetal para los ensayos moleculares.
Tratamiento Inhibidores de la EPSPS
Las plantas se cultivaron según se describe en la sección anterior con algunas diferencias: durante la etapa de 4-5 hojas, las plantas fueron sometidas a tratamiento con glifosato (Roundup®, SC, 360 g i.a./L, Bayer Cropscience) con dosis de: 0, 33.7, 67.5, 135, 270, 540, 1080 g i.a./ha para WT, y 135, 270, 540, 1080, 2160, 4320 g i.a./ha para MB y TA. Los rango de dosis son diferentes porque el WT se controla completamente a 540 g i.a./ha, se ve inútil añadir más dosis. El diseño experimental fue completamente aleatorio con tres repeticiones (una bandeja, una repetición). Cuatro semanas después de la aplicación, se determinó el porcentaje de plantas sobrevivientes y se recolectó el material vegetal para los ensayos moleculares.
Extracción de ADN
Se recolectaron muestras frescas de hojas de plantas sobrevivientes a los diferentes herbicidas para cada población. Todas las muestras se almacenaron a -80°C hasta que se extrajo el ADN genómico y el extra cromosómico utilizando el protocolo de extracción con urea (Miriam Gil, UPNA, comunicación personal). La concentración y calidad del ADN se determinaron utilizando un espectrofotómetro NanoDrop 2000 (Thermo Fisher Scientific).
Mutaciones en el gen ALS
Las regiones del gen ALS CAD (desde los aminoácidos 124 a 205) y la región BE (desde los aminoácidos 574 a 653) fueron amplificadas con los primers Ap_3F, Ap_4R y Ap_2F, Ap_2R respectivamente (Scarabel et al., 2007). Después de la amplificación, los productos de PCR fueron purificados y secuenciados. Los resultados fueron visualizados y alineados utilizando Geneious Prime® 2023.2.1.
Mutaciones en el gen EPSPS
La región TAP del gen fue amplificada utilizando los primers EGF y EGR (Gaines et al., 2010) utilizando la misma reacción descrita anteriormente. Las condiciones de PCR fueron 95°C durante 5 minutos; 35 ciclos de 95°C durante 30 segundos, 61°C durante 30 segundos, 72°C durante 15 segundos, y una extensión final a 72°C durante 5 minutos. Después de la amplificación, los productos de PCR fueron purificados y secuenciados. Los resultados fueron visualizados y alineados utilizando Geneious Prime® 2023.2.1.
Número de copias del gen EPSPS
Se realizó una reacción de qPCR para evaluar el número relativo de copias del gen EPSPS y normalizarlo con β-tubulina (Koo et al., 2018) utilizando los primers: EPSPS_F, EPSPS_R3, β-tubulin_F y β-tubulin_R y el aumento relativo en el número de copias genómicas de EPSPS se expresó como 2ΔCt (Gaines et al., 2010). Cada muestra se evaluó por triplicado.
Resistencia múltiple a nivel de planta
Para caracterizar los mecanismos de resistencia múltiple a inhibidores de ALS y EPSPS a nivel de planta, se seleccionaron cinco biotipos de cada planta madre (total diez plantas por población) que sobrevivieron a glifosato y en los cuales se confirmó el mecanismo de resistencia. Estos biotipos seleccionados se utilizaron para la amplificación parcial del gen ALS siguiendo el protocolo descrito anteriormente.
RESULTADOS Y DISCUSIÓN
Los bioensayos con herbicidas junto con estudios moleculares y secuenciación confirmaron la resistencia múltiple a los inhibidores de ALS y EPSPS en dos poblaciones españolas de A. palmeri. Los patrones de resistencia a THIF e IMA fueron similares en ambas poblaciones, con mayor supervivencia al primero y muy baja al segundo, indicando la ausencia de mecanismos de resistencia cruzada. Se encontraron distintas mutaciones en los codones: Pro-197-Thr, Asp-376-Glu y Trp-574-Leu (Figura 1). Estudios de dosis-respuesta con glifosato mostraron que biotipos resistentes de ambas poblaciones sobrevivieron a 540 g i. a/ha del producto comercial, mientras que el WT fue controlado completamente. No se encontraron mutaciones en la secuencia génica de EPSPS. Las pruebas de qPCR confirmaron que la sobreexpresión del este gen es el mecanismo primario de resistencia en ambas poblaciones, con un número mayor de copias entre 39 y 120 respecto a las plantas WT. La sobreexpresión del gen EPSPS es un mecanismo común de resistencia al glifosato en A. palmeri (Gaines et al., 2010; Küpper et al., 2017, 2018) lo que podría justificar su supervivencia al glifosato. Diez plantas por población supervivientes a glifosato se han ensayado para la confirmar la acumulación de mecanismo de resistencia múltiple a nivel de planta buscando mutaciones en el gen ALS. Se encontraron tres mutaciones en posiciones Pro-197, Asp-376 y Trp-574. Sorprendentemente, en Pro-197 se confirmaron cuatro sustituciones aminoacídicas: Pro-197-Thr, Pro-197-Ser, Pro-197-Ala y Pro-197-Ile. Cuatro plantas presentaban sustitución de glutámico por aspártico en el codón Asp-376 y tres con mutación en Trp-574 (leucina por triptófano). En tres biotipos se encontraron acumulaciones de alelos mutados, dos con Pro-197-Thr junto con Asp-376-Glu y una con Pro-197-Thr y Trp-574-Leu. Aunque estudios previos han reportado la coexistencia de diversas mutaciones alélicas dentro de una misma población (Küpper et al., 2018; Singh et al., 2019; Manicardi et al., 2023) nunca se había reportado antes la acumulación de mutaciones ampliamente reconocidas que confieren resistencia a inhibidores de ALS a nivel de planta en esta especie. Se necesitan más estudios para descartar la presencia de eventuales mecanismo de resistencia non-target site.
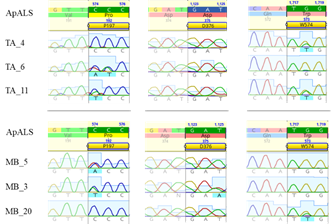
Figura 1 Detalles del cromatograma de las mutaciones de ALS encontradas en este estudio. Se muestran tres secuencias de las poblaciones TA (TA) y MB (MB) alineadas al genoma de referencia (ApALS). Las mutaciones en los codones Pro-197, Asp-376 y Trp-574 están resaltadas. Los cambios de aminoácidos son: CCC a prolina (197), ACC a serina, ATC a isoleucina y ACC a treonina; GAT a ácido aspártico y GAA a ácido glutámico; TGG a triptófano y TTG a leucina. El doble pico indica heterocigosis.
CONCLUSIONES
En este estudio se confirma por primera vez la presencia en España de poblaciones de A. palmeri con resistencia múltiple a los inhibidores de la ALS y la EPSPS, siendo además el primer caso en Europa para esta especie. También se identificó la presencia de biotipos con múltiples mecanismos de resistencia target-site a ambos MoA.
Aunque faltan estudios genéticos, estos datos sugieren el desarrollo inicial de estos mecanismos en el país de origen y su posterior selección in situ. Esta conclusión se respalda con la alta variabilidad de mutaciones alélicas en el gen ALS.
Dada la presencia de poblaciones resistentes a múltiples mecanismos de acción y sus continuas introducciones, es fundamental aplicar estrategias de manejo integrado para contener su expansión tanto en áreas agrícolas como en márgenes de carreteras. Reducir la presión de selección de herbicidas, utilizando medios de control alternativo, es crucial para limitar su propagación en cultivos y garantizar la eficacia de los productos herbicidas.

