











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Las especies invasoras son uno de los principales problemas medioambientales, suponiendo un reto para la conservación de la biodiversidad y la gestión sostenible de los recursos naturales (Rai et al., 2020). Además, previsiblemente, el cambio climático aumentará la posibilidad de invasión favoreciendo a ciertas especies (Hulme, 2017). Carpobrotus sp.pl. destaca como una de las más peligrosas debido a su capacidad invasora (Battisti et al., 2021). Es originaria de zonas costeras de Sudáfrica (Campoy et al., 2018) y fue introducida fuera de su área de distribución a principios del siglo XX como medida para la estabilización de dunas, así como planta ornamental (Preston & Sell, 1988). Actualmente, se ha naturalizado ampliamente en numerosos hábitats costeros fuera de su área nativa (Campoy et al., 2018).
Un estudio reciente (Novoa et al., 2023), identificó tres clústeres genéticos distintos del género Carpobrotus sp.pl., presentes en áreas nativas y no nativas, los cuales no se pueden distinguir por sus características morfológicas. Así, sabemos que en la Península Ibérica habitan dos clústeres, el clúster A, originario de la provincia del Cabo Occidental (Sudáfrica) y presente en Punta de Rons (Pontevedra), A Lanzada (Pontevedra) y Cádiz (Cádiz) y el clúster B, poblaciones originarias de Mdumbi (Sudáfrica) y con presencia en La Marina (Alicante) y Samil (Pontevedra). Debido a la amenaza de invasión, es fundamental analizar y comprender los mecanismos de adaptación de Carpobrotus (Drenovsky et al., 2012). En este contexto, la metabolómica ofrece una valiosa información sobre la influencia de la genética y del entorno en la producción de determinados compuestos en respuesta a condiciones ambientales cambiantes. Por esta razón, se examinaron los perfiles metabolómicos de plántulas de Carpobrotus sp.pl. de cuatro poblaciones de España. La cromatografía de gases acoplada a espectrofotometría de masas (GC-MS) se utilizó para llevar a cabo el análisis. Este método muestra cómo los factores genéticos y ambientales afectan la adaptación de las plantas de este género invasor.
MATERIALES Y MÉTODOS
Dos poblaciones de cada clúster genético presente en España, pero procedentes de diferentes regiones climáticas, fueron elegidas: A Lanzada (Pontevedra) y Cádiz (Cádiz) para el clúster A, y La Marina (Alicante) y Samil (Pontevedra) para el clúster B. Se recolectaron las plántulas, se tomaron los datos morfológicos iniciales y se cultivaron en invernadero durante cincuenta días con riego semanal. Después de los tratamientos, se evaluaron diversos parámetros de crecimiento.
Tabla 1 Poblaciones incluidas en el estudio, indicando el clúster genético y la región climática
| Origen | Clúster genético | Región climática |
|---|---|---|
| A Lanzada | A | Atlántica |
| Samil | B | Atlántica |
| Cádiz | A | Mediterránea |
| La Marina | B | Mediterránea |
En el material vegetal de la parte aérea, se llevó a cabo un análisis metabolómico no dirigido. La identificación y la cuantificación se realizaron mediante cromatografía de gases acoplada a espectrofotometría de masas (GC-MS). Para encontrar diferencias significativas entre poblaciones, se utilizaron pruebas estadísticas como ANOVA, Kruskal-Wallis y análisis univariante (dependiendo de la normalidad y homogeneidad de los datos de origen). Por último, se empleó un análisis multivariante (PCA, PLS-DA) para mejorar la comprensión de la variación metabolómica y sus implicaciones biológicas.
RESULTADOS Y DISCUSIÓN
Parámetros morfológicos iniciales y de desarrollo
Las poblaciones de la región climática mediterránea (Cádiz y La Marina), presentaron un peso y longitud inicial de la parte aérea significativamente más bajos en comparación con las poblaciones del área atlántica, especialmente en contraste con las plántulas de Samil (Figura 1A y 1B). Estas diferencias podrían atribuirse a la presencia de limitaciones hídricas en el área mediterránea. Mientras en las poblaciones atlánticas las precipitaciones superan los 1100 mm anuales (https://www.meteogalicia.gal), en la región mediterránea no llegan a la mitad con 492 mm en Cádiz y 360 mm en La Marina durante el año 2021 (año previo al ensayo). Estas alteraciones en los rasgos de respuesta a las condiciones ambientales permiten que las plantas puedan responder de forma adaptativa a una variedad de entornos más amplia que si los rasgos fueran “inalterables” (Sultan, 1995), lo que se conoce como plasticidad fenotípica. No obstante, las plántulas de Cádiz mostraron un número inicial de verticilos notablemente mayor en comparación con el resto de las poblaciones. En todos los parámetros iniciales, las plantas de La Marina mostraron los valores más bajos.
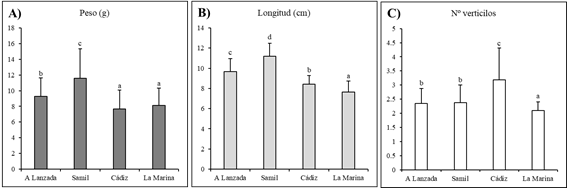
Figura 1 Peso (g), longitud (cm) y número de verticilos al inicio (t=0), antes del trasplante, de las plántulas de las cuatro poblaciones de Carpobrotus sp.pl. Diferentes letras indican diferencias estadísticamente significativas entre poblaciones (p < 0.05). N=112.
Después de un período de crecimiento de cincuenta días se midieron los parámetros de crecimiento de las réplicas destinadas a metabolómica, revelando resultados similares, con excepción del peso y contenido de agua de las raíces (Figura 2). En particular, A Lanzada mostró los valores más altos de peso en raíces, mientras que las poblaciones mediterráneas mostraron generalmente valores más bajos en línea con los parámetros inicialmente registrados. En Cádiz y La Marina, las poblaciones mediterráneas, el peso de la raíz fue un 50% menor que en A Lanzada, mientras que en Samil fue un 17% menor en comparación con A Lanzada. Las raíces de Cádiz mostraron una disminución del 10% en el contenido de agua en comparación con las otras tres poblaciones.
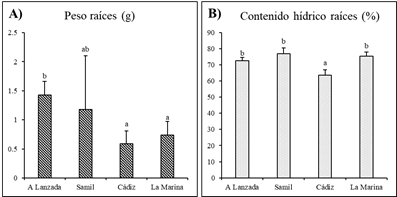
Figura 2 Peso (g) contenido hídrico (%) de las raíces al final del periodo de cincuenta días de crecimiento de las plántulas de las cuatro poblaciones de Carpobrotus sp.pl. Diferentes letras indican diferencias estadísticamente significativas entre poblaciones (p < 0.05). N=7.
Análisis comparativo del perfil metabolómico
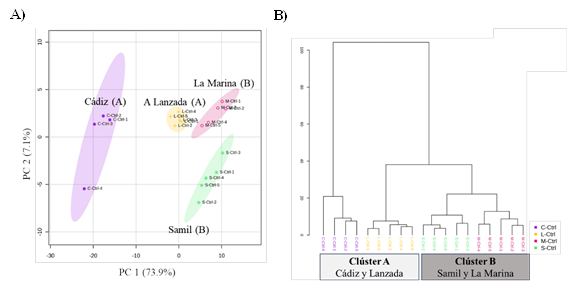
El software MS-DIAL identificó un total de 578 compuestos, de los cuales 389 eran desconocidos y 189 fueron anotados según la iniciativa de la Sociedad de Metabolómica, de estos, 158 metabolitos fueron finalmente identificados. Para evaluar la influencia de las poblaciones en los metabolitos en general se analizaron los datos mediante un análisis de componentes principales (PCA) (Figura 3a).
Los dos primeros componentes describieron el 81% de la variabilidad total, mostrando una clara distinción entre las cuatro poblaciones. El primer componente (CP1) representó la variación más alta con un 73,9% separando la población de Cádiz del resto, y agrupando las poblaciones del clúster B. En este componente los metabolitos que representaron un mayor peso fueron D-Arabitol, D-(+)-Ácido galacturónico, ácido hexanoico, 2-Hidroxipiridina y ácido nicotínico, que se acumularon de forma más significativa en las plantas de Cádiz seguidas de las de A Lanzada (perteneciente al mismo clúster genético).
Por otro lado, el componente 2 (CP2) representó un 7,1% de la variabilidad total, y está definido principalmente por el ácido glucónico, glicerol-3-galactósido-2, fosfato, lactulosa y glucoheptulosa, donde los cuatro últimos compuestos se acumulan de forma más significativa exclusivamente en plantas originarias de Samil y el ácido glucónico tanto en las de Samil como en las de Cádiz. CP2 separó claramente a Samil de las poblaciones de A Lanzada y Marina, mientras que las muestras de Cádiz mostraron una tendencia intermedia.
La agrupación observada durante el análisis de PCA se confirmó aún más mediante el análisis clúster (Figura 3b), que confirmó la formación de cuatro grupos principales separados correspondientes a las cuatro poblaciones. El análisis agrupó en un mismo conjunto a las poblaciones por clúster, por un lado, Samil y La Marina, y en una unidad separada Cádiz y A Lanzada.
Además, el análisis de la importancia variable en las puntuaciones de proyección (VIP) (basado en los 15 metabolitos con la puntuación VIP más alta, superior a 1,4) señaló que varios ácidos orgánicos, en concreto ocho de los quince encontrados, correspondieron a este grupo, siendo las clases de compuestos que contribuían principalmente a la discriminación de grupos con especial influencia del D- Arabitol, el ácido quínico y el ácido shikímico.
CONCLUSIONES
Este enfoque integral permite el estudio de las variaciones a nivel morfológico y metabolómico en especies del género Carpobrotus sp.pl. atendiendo a diferentes condiciones climáticas, atlánticas o mediterráneas, y genéticas, clúster A o B. Por una parte, a nivel morfológico, las plántulas de regiones mediterráneas presentaron un tamaño inicial inferior, probablemente debido a adaptaciones a condiciones ambientales adversas como la sequía. Por otra, a nivel metabolómico las cuatro poblaciones son distintas, con diferencias más notables entre las poblaciones del clúster A que entre las del clúster B. Estas variaciones están marcadas principalmente por la acumulación de metabolitos, en su mayor parte ácidos orgánicos, en las plántulas de la población de Cádiz seguido de las de A Lanzada, ambas pertenecientes al mismo clúster genético. Los resultados muestran cómo el factor climático tiene un menor peso en el clúster B, ya que las diferencias entre poblaciones no son tan marcadas entre poblaciones. Finalmente, el factor genético tiene un mayor peso en el perfil metabolómico de Carpobrotus que el climático, a diferencia de los parámetros morfológicos analizados, dónde el entorno mostró una mayor influencia.

