











 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkO QUE SÃO
Os nemátodes entomopatogénicos (NEP) são nemátodes benéficos que causam doença e morte de insetos, sendo utilizados como agentes de proteção biológica em Portugal e no mundo (Figueiredo, 1997; Amaro, 2003; Dolinski, 2020; Koppenhöfer et al., 2020; Poinar & Grewal, 2012). Estes organismos têm sido isolados de solos de diferentes ecossistemas, desde o ártico até os trópicos (Dolinski, 2020). A existência de uma grande variedade de espécies e de linhagens geográficas de NEP confere a estes organismos um potencial elevado como agentes de proteção biológica, contra o ataque de populações de insetos fitófagos.
Os NEP são animais que pertencem ao filo Nematoda, têm corpo filiforme e dimensão reduzida, em regra cerca de 1 mm (Wood, 1988; Dolinski, 2020). Os NEP mais estudados pertencem às famílias Steinernematidae (géneros Steinernema Travassos 1927 e Neosteinernema Nguyen & Smart 1994) e Heterorhabditidae (género Heterorhabditis Poinar 1976) (Stock & Hunt, 2005; Poinar & Grewal, 2012).
A primeira espécie de NEP descrita por Steiner (1923), Aplectana kraussei Steiner, atualmente designada por Steinernema kraussei (Steiner, 1923), foi isolada a partir de um cadáver do escaravelho-japonês Popillia japonica Newman, 1838 (Coleoptera, Scarabaeidae) (Poinar & Grewal, 2012; Dolinski, 2020). Atualmente, existem mais de 100 espécies descritas e validadas, sendo que mais de 70% das espécies conhecidas foram descritas nos últimos 20 anos (Dolinski, 2020).
A primeira espécie de NEP assinalada em Portugal foi Steinernema carpocapsae (Weiser, 1955), em 2000, nos grupos de ilhas orientais e centrais do Arquipélago dos Açores (Rosa et al., 2000). Em 2006, foi iniciada a monitorização para estudar a presença e a distribuição de NEP em todo o território continental de Portugal; sobretudo em habitats considerados adequados para estes organismos. Steinernema feltiae (Filipjev, 1934) foi a primeira espécie de NEP a ser identificada em Portugal Continental, a partir de vários isolados recolhidos em diversos habitats do sul do país (Valadas et al., 2007). Posteriormente, foram identificadas outras espécies pertencentes às famílias Steinernematidae e Heterorhabditidae, assim como a predominância dessas espécies em Portugal Continental (e.g., Valadas et al. 2009, 2011, 2014).
CICLO DE VIDA
O desenvolvimento de NEP (Figura 1) inclui três estádios: ovo-juvenil-adulto. O estádio juvenil é composto por quatro estádios: J1, J2, J3 e J4 (Dillman & Sternberg, 2012; Trejo-Meléndez et al., 2024). O estádio utilizado nas formulações para proteção biológica, designado por juvenil infectante (JI), é o J3, mas ainda sem ter efetuado a ecdisis (ou muda), ou seja, ainda apresenta a cutícula de J2. O NEP penetra no hospedeiro ainda com a cutícula de J2 e só realiza a ecdisis no seu interior, para evitar o reconhecimento pelo hospedeiro (Figueiredo, 1997).
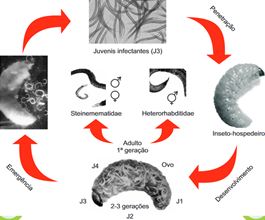
Figura 1 Ciclo de vida de nemátodes entomopatogénicos pertencentes aos géneros Steinernema e Heterorhabditis (Dolinski, 2020).
As fêmeas e os machos da primeira geração são formados a partir do J4, no caso das espécies que apresentam reprodução sexuada; as fêmeas da primeira geração colocam ovos que darão origem à segunda geração, ainda no interior do inseto-cadáver (Forst & Clarke, 2002; Dolinski, 2020). Os NEP podem ter duas ou três gerações no interior do inseto (1-2 semanas), dependendo da disponibilidade de alimento. Quando o alimento se esgota, o JI abandona o cadáver para procurar novo hospedeiro (pode durar dias ou meses) (Kaya, 1990; Dolinski, 2020, Trejo-Meléndez et al., 2024).
Apesar da infeção ocorrer de forma similar nos géneros Steinernema e Heterorhabditis (Figura 1), neste último há o aparecimento de adultos hermafroditas de primeira geração, em vez de adultos anfimíticos (machos e fêmeas). Os hermafroditas autofecundam-se e produzem ovos que darão origem a juvenis que se desenvolverão, transformando-se em adultos anfimíticos de segunda geração. Após o acasalamento, surgem novamente ovos e juvenis que, depois de se alimentarem do que restou do cadáver, migram para o solo para procurarem novo hospedeiro (Stock & Blair, 2008).
MODO DE AÇÃO
De um modo geral, o JI penetra no hospedeiro pelos orifícios naturais do inseto (boca, ânus, espiráculos ou estigmas) ou por feridas. No caso de Heterorhabditis sp., pode ainda penetrar através da cutícula intacta nas zonas intersegmentares em que não se diferenciou a exocutícula (Figueiredo, 1997; Poinar & Grewal, 2012; Dolinski, 2020). No seu interior, o NEP atinge a hemolinfa (em resultado da libertação de enzimas proteolíticas por uma bactéria simbionte) e liberta a bactéria. Esta, pela ação de toxinas, causa septicemia nos hospedeiros infetados, que ocorrerá apenas em 24-48 h (Poinar & Grewal, 2012; Dolinski, 2020). As bactérias produzem também antibióticos para evitar ataques de organismos oportunistas e multiplicam-se rapidamente para servir de alimento aos estádios juvenis (Griffin et al., 2005; Poinar & Grewal, 2012).
Os NEP possuem uma associação simbionte com γ-proteobactérias da família Enterobacteriaceae, tendo sido encontrados dois géneros de bactérias, Xenorhabdus e Photorhabdus, associadas a Steinernema e a Heterorhabditis, respetivamente. Cada espécie de NEP possui a sua bactéria simbionte específica (Dolinski, 2020). As bactérias não só exibem extrema virulência, mas também: (1) auxiliam no desenvolvimento e adaptação do seu NEP; (2) defendem o inseto-cadáver contra predadores, necrófagos e competidores; (3) produzem nutrientes a partir do inseto-cadáver; (4) promovem o desenvolvimento e a reprodução do NEP e; (5) defendem os NEP do sistema imunitário do hospedeiro. Por sua vez, os NEP protegem a bactéria de riscos ambientais no solo e transportam-na de um inseto para a hemolinfa de outro, levando-as diretamente a um ambiente rico em nutrientes (Trejo-Meléndez et al., 2024). As bactérias simbiontes produzem também pigmentos que conferem ao cadáver cores características, permitindo a rápida identificação em campo. Enquanto o cadáver infetado pelo complexo Heterorhabditis-Photorhabdus adquire uma cor avermelhada ou alaranjada e é bioluminescente, o cadáver infetado pelo complexo Steinernema-Xenorhabdus adquire uma coloração que varia entre bege a pardo-escuro, sem apresentar bioluminescência (Dolinski, 2020).
Por sua vez, o inseto pode desencadear uma série de reações de defesa contra o NEP, como o encapsulamento (mediante o processo de melanização do NEP). Contudo, normalmente a bactéria mata o hospedeiro antes que seja desencadeada uma resposta letal (Poinar & Grewal, 2012). Assim, a relação nemátode-inseto resulta da capacidade do NEP escapar às defesas do inseto, permitindo falha no reconhecimento e/ou destruição das defesas.
A localização do inseto-hospedeiro pelos NEP é altamente especializada e pode envolver três tipos de estratégias de procura: movimento tipo “procura ativa” (cruiser); por “emboscada” (ambusher); e estratégia mista (Lewis et al., 2006). Os estímulos utilizados pelos NEP podem ser de natureza física e química, como vibrações, níveis de CO2, produtos de excreção, gradientes de temperatura (Dolinski, 2020). Os NEP, que apresentam comportamento tipo “procura ativa”, buscam ativamente os seus hospedeiros no solo e respondem positivamente aos seus sinais voláteis, deslocando-se até encontrarem o hospedeiro, como é o caso das espécies Heterorhabditis bacteriophora (Poinar, 1976) e Steinernema glaseri (Steiner, 1929). As espécies que utilizam a estratégia “emboscada” apresentam uma movimentação designada por nictação, que consiste na suspensão do corpo que fica apoiado apenas na ponta posterior. Desta forma, a parte anterior fica livre a aguardar a passagem de um hospedeiro para então “saltar” sobre ele. Os NEP S. carpocapsae e S. scapterisci Nguyen e Smart 1990 são exemplos de espécies que fazem nictação (Ishibashi & Kondo, 1990). Ainda existem espécies que, de acordo com a distância a que estão do hospedeiro, apresentam ambos os comportamentos (“procura ativa” e “emboscada”) (Grewal et al., 1994a).
APLICAÇÃO
Os NEP podem ser aplicados com quase todos os equipamentos de pulverização, terrestres ou aéreos. Qualquer que seja o sistema de pulverização utilizado, deve garantir-se a agitação adequada (com baixo estresse mecânico) durante a preparação e a aplicação (Shapiro-Ilan et al., 2012).
Diversos fatores abióticos, como por exemplo, radiação ultravioleta (UV), temperatura, humidade, textura do solo, pH, fertilizantes e produtos fitofarmacêuticos podem influenciar negativamente a sobrevivência do JI, pelo que o seu conhecimento é fundamental para definir qual a melhor estratégia de aplicação do NEP (Grewal et al., 1994a; Koppenhöfer et al., 1995; Shapiro-Ilan et al., 2012; Dolinski, 2020). Entre os fatores mais importantes, constam os efeitos negativos da radiação UV, recomendando-se a aplicação ao final da tarde ou de manhã cedo. No caso de aplicações foliares, é aconselhado o uso de molhantes para preservar a integridade do JI até que penetre no hospedeiro (Dolinski, 2020). Quanto à humidade do solo, pH e temperatura, existem valores ótimos em função da espécie. Por exemplo, algumas espécies, como Heterorhabditis indica Poinar, Karunakar & David, 1992, Steinernema glaseri e S. riobrave Cabanilla, Poinar & Rauston, 1994, são relativamente tolerantes ao calor, enquanto H. megidis Poinar, Jackson e Klein, 1987, S. feltiae (Filipjev, 1934) e H. marelatus Liu & Berry, 1996 são mais tolerantes a temperaturas baixas (Grewal et al., 1994b).
Organismos como fungos, bactérias e nemátodes predadores fazem parte dos fatores bióticos que também podem afetar a eficácia dos NEP em campo (Dolinski, 2020).
É necessário respeitar as recomendações do fabricante em relação às condições de armazenamento e aplicação de NEP, assim como as contraindicações, em particular as incompatibilidades com fertilizantes e produtos fitofarmacêuticos.
VANTAGENS E LIMITAÇÕES À UTILIZAÇÃO
Atualmente, são vários os atributos que fazem dos NEP potenciais agentes de proteção biológica (por exemplo, Dolinski, 2020; Koppert, 2024): (1) são produzidos com facilidade, seja em insetos-hospedeiros, ou meios artificiais; (2) podem ser armazenados; (3) são facilmente aplicados em campo, na água de rega ou por pulverização; (4) possuem a habilidade de procurar ativamente o hospedeiro; (5) são compatíveis com a maioria dos produtos fitofarmacêuticos; (6) são seguros para organismos não-alvo incluindo humanos, dado que possuem um estreito espectro de hospedeiros, sendo muito específicos (não há risco de causarem mortalidade indiscriminada de organismos não alvo); (7) reproduzem-se no hospedeiro produzindo novas gerações; e (8) originam zero-resíduos com impacto para o solo, águas subterrâneas e cadeias alimentares. As bactérias simbiontes associadas aos NEP não são consideradas perigosas para o ambiente, pois a sua permanência no solo é mínima e não possuem formas de sobrevivência (Akhurst & Smith, 2002). A escolha do NEP deve, no entanto, ser feita de forma cuidadosa, privilegiando a utilização de NEP nativos, relativamente a espécies exóticas, uma vez que os NEP nativos estarão adaptados às condições climáticas e à entomofauna local (Lacey & Georgis, 2012).
A principal limitação à utilização de NEP é ainda o custo da sua produção, em particular a multiplicação de NEP in vitro com recurso a meio líquido (Dolinski, 2020). Outra das limitações que tem sido associada à utilização de agentes de proteção biológica (NEP ou outro) é a necessidade de supervisão especializada (por exemplo, Devi et al., 2023). Todavia, cada vez mais, a atividade agrícola para ter sucesso terá que incorporar novos conhecimentos e o uso de NEP continuará a crescer à medida que os agricultores e os consumidores se tornarem mais conscientes dos seus benefícios.
EXEMPLOS DE UTILIZAÇÃO
A comercialização de NEP foi iniciada em 1976 nos EUA (Poinar & Grewal, 2012), mas na época não teve êxito, devido ao elevado custo de produção e à competição com a utilização de Bacillus thuringiensis (Bt). Os progressos nos métodos de produção proporcionaram novas possibilidades de êxito na comercialização, considerando-se, atualmente, que pertencem ao grupo dos bioinsecticidas mais importantes, logo a seguir a Bt (Amaro, 2003).
Em alguns países, como na União Europeia, a comercialização de NEP é permitida sem exigência prévia de homologação (Amaro, 2003), porque são considerados macrorganismos. Em muitos países têm sido em grande parte excluídos dos requisitos de registo de produtos fitofarmacêuticos, dado o seu elevado nível de segurança para humanos, organismos não-alvo e meio ambiente, características que facilitaram a comercialização, originando o desenvolvimento comercial de pelo menos cinco espécies de Heterorhabditis e oito espécies de Steinernema (Quadro 1) (Koppenhöfer et al., 2020).
Quadro 1 Nemátodes entomopatogénicos pertencentes aos géneros Steinernema e Heterorhabditis comercializados por região do mundo (Af- África, As- Ásia, Au- Austrália, AN- América do Norte, AS- América do Sul, Eu- Europa) (adaptado de Koppenhöfer et al., 2020)
| Género | Espécie | Região | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Heterorhabditis | H. bacteriophora | AN, Eu | H. indica | AN | H. marelata | AN | H. megidis | AN, Eu | H. zealandica | AN, Au | ||||||
| Steinernema | S. carpocapsae | Af, AN, AS, As, Au, Eu | S. feltiae | AN, Eu | S. kraussei | AN, Eu | S. kushidai | As | S. longicaudum | As | S. riobrave | AN | S. svapyerisci | AN | S. scarabaei | AN |
Recentemente, Koppenhöfer et al. (2020) compilaram os NEP que têm sido usados comercialmente no mundo (ou se mostraram promissores), por espécie de inseto fitófago e por cultura (Quadro 2).
Quadro 2 Lista de espécies de nemátodes entomopatogénicos (NEP) (Hb- Heterorhabditis bacteriophora, Hd- H. downesi, Hm- H. marelata, Hmeg- H. megidis, Hz- H. zealandica, Sc- Steinernema carpocapsae, Sf- S. feltiae, Sg- S. glaseri, Sk- S. kushidai, Sr- S. riobrave, Ss- S. scapterisci, Ssc- S. scarabaei) comercializados, com indicação da cultura afetada e da espécie de inseto fitófago (adaptado de Koppenhöfer et al., 2020)1
| Cultura | Ordem inseto | Família inseto | Nome comum | Nome científico | Espécie primária de NEP usada |
|---|---|---|---|---|---|
| Alcachofra | Lepidoptera | Pterophoridae | Platyptilia carduidactyla | Sc | |
| Banana | Coleoptera | Curculionidae | Gorgulho-da-bananeira | Cosmopolites sordidus | Hb, Sc, Sf |
| Batata-doce | Coleoptera | Curculionidae | Gorgulho-da-batata-doce | Cylas formicarius | Hb, Sc, Sf |
| Cana-do-açúcar | Coleoptera | Curculionidae | Gorgulho-da-beterraba | Temnorhinus mendicus | Hb, Sc |
| Citrinos | Coleoptera | Curculionidae | Pachnaeus spp. | Hb, S | |
| Citrinos | Coleoptera | Curculionidae | Diaprepes abbreviatus | Hb, Hi, Sr | |
| Citrinos | Lepidoptera | Tortricidae | Thaumatotibia leucotreta | Hb | |
| Cogumelos | Diptera | Sciaridae | Moscas-do-terriço | (várias espécies) | Sf,Hb, H |
| Cogumelos | Diptera | Sciaridae | Moscas-do-terriço | (várias espécies) | Sf, Hb, Hi |
| Culturas protegidas | Thysanoptera | Thripidae | Tripe-da-California | Frankliniella occidentalis | Sc, S |
| Fruteiras | Coleoptera | Bupestridae | Carocho-negro | Capnodis tenebrionis | Sf |
| Fruteiras | Diptera | Tephritidae | Moscas-da-fruta | (várias espécies) | Hi, Sc |
| Fruteiras | Lepidoptera | Pyralidae | Amylois transitella | Sc | |
| Fruteiras | Lepidoptera | Sesiidae | Sésia | Synanthedon spp. (várias espécies) | Hb, Sc, Sf |
| Hortícolas | Diptera | Agromyzidae | Mineiras de hortícolas | Liriomyza spp. | Sc, S |
| Hortícolas | Lepidoptera | Noctuidae | (várias espécies) | Sc, Sf, Sr | |
| Hortícolas | Lepidoptera | Noctuidae | Rosca ou nóctua | Agrotis ipsilon | Sc |
| Hortícolas | Lepidoptera | Noctuidae | Lagarta-da-espiga-do-milho | Helicoverpa zea | Sc, Sf, Sr |
| Hortícolas | Lepidoptera | Noctuidae | Rosca ou nóctua | Agrotis segetum | Sc, S |
| Milho | Coleoptera | Chrysomelidae | Diabrotica spp. | Hb, Sc | |
| Palmeira | Coleoptera | Curculionidae | Escaravelho-da-palmeira | Rhynchophorus ferrugineus | Sc |
| Pequenos frutos | Coleoptera | Curculionidae | Gorgulho-da-videira | Otiorhynchus sulcatus | Hb, Hd, Hmeg, Hm, Sc, Sg, Sf |
| Pequenos frutos | Coleoptera | Curculionidae | Gorgulho-do-morangueiro | Otiorhynchus ovatus | Hm, Sc |
| Pequenos frutos | Coleoptera | Curculionidae | Rhadopterus picipes | Hb | |
| Pequenos frutos | Coleoptera | Scarabaeidae | Escaravelhos | (várias espécies) | Ssc |
| Pequenos frutos | Lepidoptera | Crambidae | Chrysoteuchia topiaria | Sc | |
| Nogueira Pecan | Coleoptera | Curculionidae | Curculio caryae | Sc | |
| Pomóideas | Lepidoptera | Tortricidae | Bichado-da-macieira | Cydia pomonella | Hz, Sc, Sf |
| Prunóideas | Coleoptera | Curculionidae | Gorgulho-da-ameixeira | Conotrachelusnenuphar | S |
| Tomateiro | Lepidoptera | Geleichiidae | Traça-do-tomateiro | Phthorimaea absoluta | Hb, Sc, S |
1 Pelo menos um artigo científico relatou ≥70% de supressão do inseto fitófago no campo.
Alguns dos exemplos de NEP comercializados em Portugal pertencem às espécies: S. carpocapsae (de uso mais generalista; para lepidópteros, coleópteros e heterópteros); S. feltiae (de uso em climas mais frios; para dípteros e coleópteros); e H. bacteriophora (para coleópteros).
CONCLUSÕES
A utilização de NEP constitui uma alternativa segura e eficaz para a redução das populações de insetos fitófagos. Os produtos à base destes organismos são excecionalmente seguros para humanos, organismos não-alvo e meio ambiente, estando por isso isentos de homologação na maioria dos países europeus, incluindo Portugal. Atualmente, existem várias opções de produtos comercializados à base de NEP no mercado, direcionados a diferentes espécies de insetos fitófagos de várias culturas agrícolas. A escolha do NEP a utilizar deve ter em conta a espécie de inseto alvo e requer algum grau de supervisão, uma vez que o sucesso na aplicação de NEP depende de um conjunto de fatores abióticos e bióticos que podem condicionar a eficácia destes organismos.

