










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkArquivos de Medicina
versão On-line ISSN 2183-2447
Arq Med vol.28 no.4 Porto ago. 2014
ARTIGO DE REVISÃO
A imunodeficiência e o sistema imunitário. O comportamento em portadores de HIV.
The immunodeficiency and the immune system. The role in HIV carriers.
Rui Soares1,2,3, Rui Duarte Armindo2, Graça Rocha1,2
1CHUC – Centro Hospitalar e Universitário de Coimbra
2Faculdade de Medicina da Universidade de Coimbra
3CnC – Centro de Neurociências e Biologia Celular de Coimbra
RESUMO
A SIDA é ainda, em pleno século XXI, responsável por centenas de milhar de mortes anualmente, um pouco por todo o mundo. Muitos estudos vão encarando a SIDA de diferentes pontos de vista: a SIDA como doença infecciosa, a SIDA como doença sexualmente transmissível, a SIDA como pandemia, mas é importante não esquecer que a SIDA é uma doença da imunidade, uma imunodeficiência. O objectivo principal desta revisão é precisamente relacionar a SIDA com os diferentes grupos de imunodeficiências, explicando também a interação do VIH com o sistema imunitário do doente. Recorrendo e citando a mais atual literatura nacional e internacional, publicada nas principais revistas do conhecimento médico, bem como a livros da especialidade, parte-se de uma breve explicação dos mecanismos básicos da imunidade humana para de uma forma mais estruturada se perceber a patologia que lhes está associada e culminando no assunto SIDA. Abordam-se imunidade inata e adquirida, bem como as principais imunodeficiências primárias e secundárias. Chega-se a uma caracterização semiológica da SIDA, sempre com as restantes imunodeficiências em mente e tendo a estrutura revista do Sistema Imunitário em conta. Os atuais critérios clínicos e laboratoriais do CDC (Center for Disease Control) dos EUA que definem um caso de SIDA permitem a esquematização de um algoritmo para seu diagnóstico diferencial com que se conclui este artigo.
Palavras-chave: sida, diagnóstico, imunidade, imunodeficiência, resposta celular
ABSTRACT
With the 21st century already here, AIDS is still responsible for thousands of deaths around the world. Many studies face AIDS from different points of view: AIDS as an infectious disease, AIDS as a sexually transmitted disease, AIDS as a pandemy, but its important not to forget that AIDS is an immune disease, an immunodeficiency. The main objective of this review is to relate AIDS with the different groups of immunodeficiencies, while also explaining the interaction of HIV with the patients immune system. Recurring and quoting the latest national and international literature published on the subject and some acclaimed text books, it is presented a brief explanation of the basic mechanisms of human immunity so that a structured comprehension of the related pathology is achieved and the AIDS subject reached. Natural and acquired immunity, primary and secondary immunodeficiencies are approached. Its presented a semiologic characterization of AIDS, always with the other immunodeficiencies in mind and having the reviewed structure of the immune system taken into account. The clinical and laboratory criteria from USAs CDC (Center for Disease Control) that define a case of AIDS allow the creation of a schematic algorithm for its differential diagnose with which this article is concluded.
Keywords: aids diagnosis, immunity, immunodeficiency, cellular response
Introdução
Por volta do século XIV, o termo immunis começou a ser relacionado com a saúde humana, indicando a capacidade de algumas pessoas escaparem a certas doenças, a que tantas outras não resistiam.1 Desde então, os conceitos foram-se desenvolvendo, até chegarmos à atual definição de sistema imunitário, uma complexa rede de células, tecidos e órgãos que trabalham, em conjunto, para defender o organismo contra os ataques de tudo quanto lhe é estranho.2
O sistema imunitário (SI) começa por excluir todos os invasores, à sua entrada, o que faz com extrema eficácia na maioria das vezes.
Como qualquer sistema desta complexidade, a imunidade humana pode apresentar defeitos, como a identificação errada do alvo, atacando aquilo que é próprio do organismo – a autoimunidade ou a falência da sua capacidade protetora – a imunodeficiência.3
No que toca às diferentes imunodeficiências, distinguem-se as primárias (IDP) defeitos genéticos que resultam na ineficácia de um ou mais dos componentes do sistema imunitário e que levam a um aumento da susceptibilidade às infecções,4 das secundárias (IDS), causadas por vários factores que afectam um sistema imunitário que, de outra forma, seria normal.5
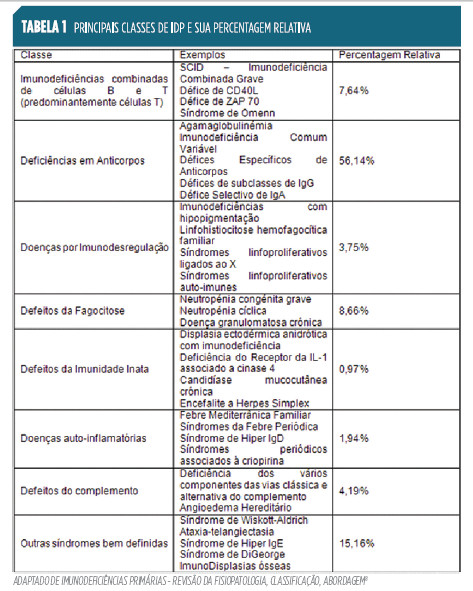
Ainda que seja considerado um campo em expansão, em que mesmo a definição pode vir a sofrer alterações6, a investigação na área das imunodeficiências primárias tem estado de acordo com a sua classificação a partir do componente do sistema imunitário afectado7 (Tabela 1).
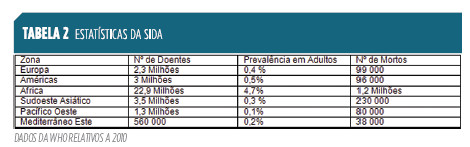
As imuno deficiências secundárias, por outro lado, são normalmente divididas em função da causa que a elas se associa (Idade Extrema, Doenças Metabólicas, Doenças genéticas, tratamento farmacológico, Cirurgia e trauma, Condições Ambientais, Doenças Infecciosas9). A mais conhecida destas é a SIDA, Síndrome da Imunodeficiência Adquirida, pelas suas características epidémicas e alta mortalidade (Tabela 2), em resultado da infecção por VIH, vírus da imunodeficiência humana.
O conhecimento dos mecanismos básicos inerentes às principais imunodeficiências, em geral, e à decorrente da SIDA, em particular, é de extrema importância para qualquer profissional de saúde.
Sistema imunitário – principais agentes de defesa
A compreensão da patologia do sistema imunitário humano implica, como em qualquer outro sistema de qualquer outro organismo, o conhecimento prévio da sua fisiologia.
A imunidade inata responde de forma não específica e indiscriminada, através de várias barreiras, constituindo a primeira linha de defesa do organismo. A característica que a distingue satisfatoriamente da restante imunidade é o facto de basear a sua atuação em informação codificada a nível germinal, em oposição aos receptores gerados por mecanismos somáticos, ao longo do desenvolvimento do indivíduo, nos quais se baseia a imunidade adquirida.10 Com esta diferença vêm outros dados importantes, invariavelmente ligados à imunidade inata–uma resposta não específica, imediata e não formadora dememória.11
Podemos considerar, dentro deste grupo de imunidade, seguindo os critérios acima referidos, quatro tipos de barreiras, com formas de funcionamento bastante distintas e dignas de um olhar mais aprofundado: anatómicas, fisiológicas, fagocíticas e inflamatórias.3
Barreiras anatómicas
Podemos considerar a pele como a principal barreira a uma eventual infecção.12 A sua espessa camada de queratina impede a penetração da maioria dos agentes com que contactamos.
O funcionamento destas primeiras barreiras do sistema imunitário envolve vários aspectos. Como interface com o exterior, impedem fisicamente a passagem do que vem de fora. Algumas células, especializadas nas diferentes barreiras da pele, são capazes de detectar e destruir material invasor. A ligeira acidez do tecido que reveste o corpo é capaz de atrasar o desenvolvimento de algumas bactérias e a secreção de sebo acaba por adicionar mais uma camada protetora. A vertente química deste nível de proteção baseia-se especialmente na secreção de lisozimas e fosfolipases (capazes de desestabilizar a paredebacteriana) e defensinas, comatividade antimicrobiana. Também a manutenção de uma flora normal na pele e tracto gastrointestinal pode prevenir a colonização por agentes patogénicos.
Quando a integridade estrutural destas superfícies é comprometida, como num eczema ou em queimaduras graves, são as infecções bacterianas as primeiras a aparecer. O Staphylococcus aureus, por exemplo, é capaz de aproveitar estas falhas para iniciar um círculo vicioso de destruição que acaba por resultar em infecções graves.13
Barreiras fisiológicas
Um conjunto de outros factores, distribuídos pelo organismo, é capaz de barrar a infecção impedindo o desenvolvimento de uma grande parte dos potenciais agentes infecciosos. Maioritariamente inespecíficos, estes vão da temperatura e do pH até várias moléculas com função imunitária.
Começando pela temperatura corporal, podemos considerar, quanto a esta, dois importantes níveis de atuação inatos. Por um lado, temos a temperatura corporal normal do organismo que, com a capacidade de se manter constante, consegue limitar a possibilidade de infecção ao restrito universo dos organismos capazes de sobreviver por volta dos37ºC.
Como a gama de agentes já adaptados a esta temperatura normal vai aumentando,14 há ainda outra resposta imunitária ativa e não específica, relacionada com a temperatura – a febre.15 A subida da temperatura na febre resulta diretamente de uma série de alterações na produção e reabsorção de calor, como vasoconstrição e contração muscular, entre outras. Estes mecanismos, por sua vez, são ativados pela libertação de citoquinas por células imunes agredidas, capazes de atuar a nível do hipotálamo. Como consequência, para além do aumento da dificuldade de desenvolvimento do invasor, temos a importante ativação de outros níveis do sistema imunitário, especialmente macrófagos,17 e alguns linfócitost.18
Curioso, neste caso, é o facto de, mesmo em situações de clara imunodeficiência (mesmo por imunossupressão), a febre se tornar a principal, e muitas vezes a única, manifestação de infecções graves.16
A acidez gástrica é fundamental para evitar a entrada definitiva de organismos capazes de chegar ao estômago, pois muito poucos conseguem sobreviver com um pH tão baixo. Dificuldades nesta capacidade de acidificar o meio, como no caso de recém nascidos, tornam também mais fáceis alguns tipos de infecção.3
Outro importante grupo de moléculas da imunidade inata está ligado à identificação em caso de infecção. A sua função vai para além da interface desta imunidade com a adquirida, dado que ativam outras respostas inespecíficas. Tomemos como exemplo uma intrusão viral num organismo humano. As grandes diferenças existentes entre metabolismos fazem com que algumas moléculas do vírus não estejam, em condições normais, num humano. O SI inato consegue detectar algumas dessas moléculas, através de receptores e, ainda que não distinga diferentes tipos de vírus para a sua resposta, consegue perceber que está perante algo que lhe é estranho. Entre estes receptores adquirem especial importância as colectinas, solúveis, e os Receptores TL, na membrana das células.
O funcionamento das colectinas baseia-se na presença dos seus domínios lectina e colagénio. O domínio lectina liga-se a açúcares dos agentes patogénicos enquanto o domínio tipo colagénio trata de se ligar a outros componentes do SI inato.3
Os receptores TL (Toll-Like Receptors) são moléculas relacionadas com uma família inicialmente descoberta no SI da mosca da fruta. Assim que um TLR reconhece e se liga a um tipo específico de alvo, activa rapidamente células efetoras condizentes.
Estudos recentes de patologia experimental em mamíferos têm mostrado que défices primários em algumas colectinas causam dificuldades ao desenrolar do processo inflamatório19 e agravam consequências de infecções como a aspergilose pulmonar invasiva.20
Há também já casos descritos de imunodeficiências primárias por mutações em genes codificantes de receptores TL ou de intervenientes nas vias que lhes estão diretamente associadas. Nestes casos, as consequências vão ainda mais além, pela sua interferência com o nível seguinte de imunidade, a adquirida.21 Assim, podem ir desde encefalites,22 no caso de alguns destes receptores que são naturalmente expressos no sistema nervoso central, a infecções bacterianas generalizadas, se for toda a via a estar afectada.23
O sistema complemento constitui mais uma parte fulcral da imunidade inata por mediadores químicos envolve cerca de 20 proteínas circulantes, ativadas por 3 vias distintas – clássica, alternativa e da lectina. Enquanto a via da lectina está relacionada com o funcionamento das colectinas, as vias clássica e alternativa começam, respectivamente, por contacto com anticorpos e com a superfície de alguns agentes patogénicos.
Qualquer que seja a forma de ativação da cascata a restante sequência de reações será a mesma e irá, progressivamente, ativar todas as moléculas do sistema, rápida e eficientemente. As três principais consequências da ativação do complemento são a formação de um complexo de ataque à membrana, um polímero que se liga e compromete a membrana do agente invasor; o aumento da permeabilidade vascular por alguns fragmentos moleculares de baixo peso; a ligação de complexos complemento agente a células efetoras com receptores próprios.
Vários tipos de deficiências podem afectar cada um dos componentes do Complemento, com diferentes repercussões clínicas. Pelo elevado número de proteínas envolvidas nesta cascata, são também muitas as imunodeficiências primárias que lhe estão associadas, sendo tanto maior o dano quanto mais cedo fosse intervir a proteína em falta. O angioedema hereditário adquire uma especial importância entre estes casos, já que a sua transmissão se faz com dominância do alelo mutado, aumentando riscos de recorrência. Resulta da deficiência da proteína inibidora de C1 e causa inchaço de tecidos subcutâneos ou membranas mucosas, podendo ser especialmente perigoso no caso de ocorrer nas vias aéreas.24
Barreiras fagocíticas
A ingestão e subsequente destruição de produtos prejudiciais é outra importante forma de defesa. Como imunidade inata, esta está a cargo especialmente de monócitos circulantes, neutrófilos e macrófagos tecidulares.
O funcionamento deste processo pode resumir-se à formação de uma vesícula com o alvo a fagocitar, o fagossoma, a que se segue a sua fusão com o lisos- soma onde estão as enzimas capazes de o digerirem produtos que podem depois ser exocitados. Várias imunodeficiências descritas resultam de interferências com o funcionamento normal destes agentes fagocitários.3
Resposta inflamatória
Podemos considerar, como mecanismo que coordena e permite todas estas formas de resposta inespecífica, a inflamação.
É uma reação do sistema imunitário inato, em que podemos encontrar cinco sinais cardinais: rubor, calor, tumor, dor e perda de função.25
A imunidade adquirida tem como marca de qualidade e unicidade a memória imunológica, propriedade que permite relembrar uma resposta imunológica específica, tornando-a ainda mais rápida e eficiente caso um novo encontro com o mesmo agente aconteça.
A resposta imunitária adaptada pode dividir-se em três fases de desenvolvimento: a fase dos linfócitos naïve, em que as células desta imunidade ainda não tiveram encontros prévios com agentes infecciosos e, portanto, manifestam simplesmente o que está codificado de base no seu genoma; a fase da resposta primária, quando há uma resposta específica, durante um primeiro contacto com o agente estranho, havendo expansão numérica (em que é dada primazia às combinações corretas de material genético para o combate específico a esse agente) e diferenciação em células efetoras de um grande número de linfócitos – deve eliminar o alvo e gerar células de memória, prontas a responder de forma rápida e específica a uma reinfecção; e uma resposta imunitária secundária, baseada nessa memória produzida no primeiro encontro, ainda mais eficaz. Esta habilidade de relembrar uma resposta prévia tem tanto de rara como de importante e, por isso, os organismos que combinam as imunidades inata e adquirida são aqueles que realmente podem sobreviver a uma longa vida num ambiente repleto de agentes patogénicos.26 O ponto de união entre os agentes mais importantes da imunidade adquirida está em células estaminais da medula óssea.25
Considera-se haver dois grandes braços de atuação através de imunidade adquirida: a imunidade humoral (com produção de anticorpos) e a mediada por células.
Imunidade Humoral
A resposta imunitária humoral, mediada por anticorpos, começa por ser induzida pela exposição a material estranho – o antigénio – que pode ser prejudicial ao organismo e que já passou as primeiras linhas de defesa deste. Reconhecido, dá-se a apresentação do antigénio, neste caso a células que irão produzir as imunoglobulinas/anticorpos, capazes de responder de forma específica à origem do antigénio em causa.
Segundo o modelo bifásico atualmente mais aceite,27 a apresentação do antigénio para resposta humoral requer quase sempre uma interação entre linfócitos B e t (especialmente os Thelper/CD4+). para além desta, a exposição fica a cargo de células apresentadoras de antigénios (APCs), na sua maoria linfócitos B, macrófagos e células dendríticas.
Apresentado o antigénio, inicia-se a última parte da maturação do linfócito B, aquela que depende da presença do antigénio e o torna num plasmócito, produtor de anticorpos. Um grupo mais reduzido de linfócitos B desenvolve-se por uma via alternativa e torna-se uma célula de memória, pronta para um segundo ataque do mesmo agente, que permite a resposta secundária.
Voltando então aos plasmócitos, estes vão ser estimulados e tornam-se efetores, libertando o anticorpo adequado na corrente sanguínea.
Os anticorpos vão, depois, se tudo correr bem, ligarse ao seu alvo, destruindo-o ou incapacitando-o.
O anticorpo é uma proteína única cuja molécula apresenta uma forma aproximada de y (molécula bifuncional) e que contém uma ponta, o paratopo, no qual podem encaixar epítopos, parte reconhecida do antigénio. Como se veio a perceber ao longo dos anos, a complexidade da interação antigénio anticorpo permite englobar mais de um modelo explicativo ao mesmo tempo, sendo aceites atualmente tanto o modelo chave-/fechadura, como o do encaixe induzido (em que as partes envolvidas se consideram bem menos rígidas do que no primeiro, ajustando-se uma à outra) e até um recente modelo de equilíbrio (que encontra um equilíbrio base entre duas formas de um mesmo anticorpo, com diferentes para topos e que evolui depois em função do antigénio apresentado28).
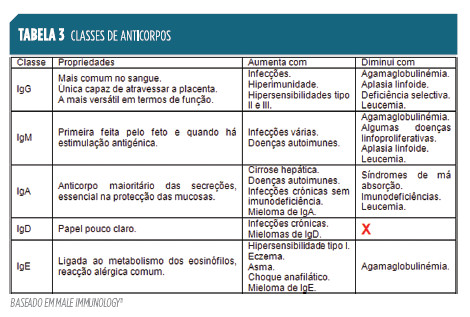
Pequenas diferenças, na sequência de aminoácidos da região constante das cadeias pesadas dos anticorpos, permitem a divisão dos mesmos em 5 classes principais: IgG, IgM, IgA, IgD, IgE11. A Tabela 3
resume as suas principais características.
O grande número de tipos de células e moléculas envolvidas aumenta também a probabilidade de ocorrer um erro algures na complexa resposta humoral. As principais imunodeficiências primárias relacionadas com a resposta humoral são divididas pela União Internacional de Sociedades Imunológicas nos seguintes grupos:29
• Ausência de linfócitos B com consequente redução de todos os tipos de anticorpos;
• Diminuição de linfócitos B e ausência de 2 ou mais tipos de anticorpos;
• Valores de linfócitos B normais com diminuição de valores de IgG e IgA e aumento de IgM;
• Valores de linfócitos B normais, mas erros estruturais nos anticorpos;
• Deficiência apenas na resposta, relacionada com grupo específico de antigénios.
A estas acrescentam-se mielomas, leucemias linfóides e a própria SIDA, todas causas secundárias que põem em risco este tipo de imunidade.
Imunidade celular
A imunidade celular, por outro lado, tem uma abordagem diferente do problema: destruir células infectadas e eliminar os seus invasores intracelulares. É que, enquanto um invasor extracelular pode, depois de sinalizado e isolado, entrar nos habituais processos de endocitose e degradação subsequente, tudo se complica quando se trata de um invasor intracelular, muitas vezes preparado para entrar precisamente por vias fagocíticas. Neste caso, a primeira forma de resposta apenas contribui para o sucesso do invasor e foi precisa, por isso, uma forma ainda mais extrema de imunidade – a imunidade mediada por células.
Para que esta ação nunca seja descoordenada, a imunidade mediada por células está diretamente relacionada com a mediada por anticorpos, nomeadamente por comunicação intercelular através de citoquinas, pequenas moléculas proteicas, a cargo das quais está uma boa parte da imunomodulação,3 aquando de uma infecção (o que por vezes acaba por envolvê-las também em processos de autoimunidade31).
Através destes mensageiros, a resposta pode ser encaminhada num sentido humoral ou celular, consoante o que se vai percebendo acerca do invasor e sua forma de atuação.
Quando esta balança pende para o lado da respostacelular,as consequências diretas estão, invariavelmente, relacionadas com:32
• Ativação de linfócitos tcitotóxicos, especificamente desenhados para atuar contra o antigénio a quese está a responder ecapazes de ativara apoptose de células infectadas, que expressam o antigénio na sua membrana;
• Ativação de linfócitos Thelper que vão:
• Ativar macrófagos e células NK que, apesar de mais comuns na resposta inespecífica, acabam aqui por ter um importante papel na destruição do invasor intracelular (macrófagos) ou da célula em condições indesejadas (célula NK). Estes dois tipos de células aparecem juntos nesta resposta, pela sua capacidade de cooperação33 e regulação recíproca;34
• Libertação de uma variedade de citoquinas, capazes de influenciar as respostas das outras células envolvidas na resposta imunitária adquirida.
Muitos factores de risco ambientais ou de ação de agentes patogénicos podem pôr em causa as células envolvidas na teia da imunidade adquirida, mediada por células. Algumas das primeiras a serem descritas relacionam-se com malnutrição,35 em especial com défices próteo-calóricos36 e stress, nos primeiros meses de vida.37
Também infecções do timo,38 órgão onde ocorre uma parte importante da maturação dos linfócitos t, podem causar imunodeficiências desta ordem, tal como os tumores a ele relacionados, entre os quais se destaca o linfoma de Hodgkin.39
As interleucinas são proteínas que podem ser produzidas por células t e por macrófagos, dependendo do estímulo que o origine. Das diversas funções das interleucinas, realçam-se as de ativação de outros linfócitos e as de indução de divisão celular. Dos estudos recentes, é importante realçar que a interleucina 17 (Il17) poderá funcionar como um marcador da expressão de CD4, isto porque esta proteína é produzida pelos linfócitos t CD4 ativados.64
Maturação das células da imunidade adquirida e problemas associados
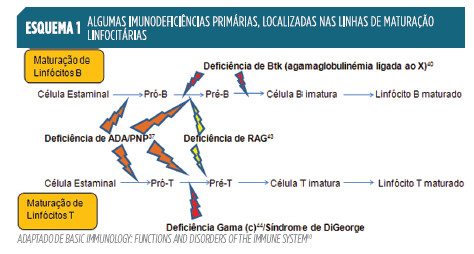
As complexas linhas de desenvolvimento associadas à imunidade adquirida, muitas das quais mantidas ativas no adulto, estão sujeitas à ocorrência de erros. O Esquema 1
mostra alguns desses problemas e a sua localização nas linhas de desenvolvimento.
VIH, SI e SIDA
A compreensão tão completa quanto possível da complexa interação do VIH com o sistema imunitário do hospedeiro é a base indispensável para uma intervenção eficaz contra a SIDA. É seguindo este mote do Doutor João Castro e Melo que passamos à segunda parte do artigo.
Vírus da imunodeficiência humana – breve descrição
O VIH (ver Figura 1
) é um vírus da família Retroviridae e do género lentivirus. É um vírus que transporta a sua informação genética em duas cadeias de RNA, de polaridade positiva, e protegido de dentro para fora por uma nucleocápside, a matriz viral, e uma cápsula maioritariamente lipídica. tudo isto acaba por resultar no tamanho de, aproximadamente, 0,1 μM, o que a torna 20 vezes menor do que uma bactéria E. coli, por exemplo.45
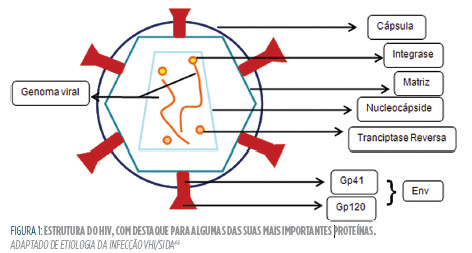
Os 9 genes do genoma do HIV-1 codificam três proteínas estruturais (gag, pol e Env), duas regulatórias (Tat e Rev) e quatro acessórias (Nef, Vif, Vpr e Vpu).46
Como entra?
Calcula-se que a origem do VIH tenha acontecido por recombinação de uma versão original que infectava símios e que terá entrado no organismo de humanos caçadores de chimpanzés. Apesar de já ter sido isolado a partir de quase todos os fluídos corporais, apenas sangue, secreções genitais e, mais raramente leite materno, estão implicados na sua transmissão.47
Não há casos descritos de transmissão via vector artrópode ou por contacto casual.
Assim, podemos considerar, como portas de entrada do HIV, todos os locais que permitam o contacto de um destes fluídos de um indivíduo infectado com a corrente sanguínea de um saudável. O VIH passa de um humano para outro por transmissão sexual, através de agulhas/seringas infectadas, da mãe para o filho (in útero, durante o parto ou por aleitamento), ou através de uma transfusão sanguínea, por exemplo. Ainda que a pele seja normalmente uma boa barreira protetora do sangue, a sua lesão pode levar ao contágio. Ainda menos eficazes costumam ser as mucosas, como a genital, oral ou mesmo nasal e do olho, e que são também, por isso, importantes portas de entrada do vírus.48
Acredita-se que, em contacto com mucosas do organismo a infectar, o vírus se aproveita de algumas células apresentadoras de antigénios, muito abundantes e distribuídas, para chegar àquelas que vai de facto infectar e usar para se reproduzir.49
Que alvos? Como infecta?
As designações CD (cluster of differentiation) indicam um conjunto de moléculas de superfície celular que diferenciam os diferentes linfócitos.
No caso do VIH ganham especial importância as células que expressam CD4, sendo as mais importantes alguns linfócitos Thelper, monócitos, macrófagos e células dendríticas. Tradicionalmente, considera-se que a entrada do vírus nacélula se dá por fusão direta, começando com a ligação do CD4 à glicoproteína gp120 (de fixação), com a ajuda de um co-receptor próximo, normalmente CXCR4 (especialmente em macrófagos) ou CCR5 (característico dos linfócitos Thelper50). Esta ligação desencadeia então uma série de acontecimentos sequenciais, incluindo algumas mudanças de conformação, que permitem o envolvimento da gp,41 a glicoproteína de fusão propriamente dita, no processo.51 Dá-se a fusão das duas membranas e a entrada da nucleocápside do vírus.
Dentro da célula-alvo, vão ser as proteínas acompanhantes do RNA a ganhar destaque. A transcriptase Reversa sintetiza uma primeira cadeia de DNA, degrada o RNA e completa o DNA, formando a cadeia dupla que o caracteriza. Com ajuda da proteína acessória Vpr, o complexo conjunto é transportado para dentro do núcleo da célula, onde a integrase faz o seu trabalho, juntando genoma viral ao original. Este, quando expresso, resulta na multiplicação do vírus, que é depois libertado por gemulação.
Como escapa?
Como foi visto na primeira parte deste artigo, são muitas as formas de o organismo se defender de agressores como o VIH. A verdade é que o vírus consegue escapar a um sistema imunitário competente, a maioria das vezes, deixando-o incapacitado. Se falarmos de respostas a infecções virais em geral, a resposta humoral tem, normalmente, por objectivo, o controlo da disseminação e o evitar da reinfecção, enquanto a celular é importante na indução da morte de células infectadas e, consequentemente, do vírus que as infecta.52
O caso do VIH, contudo, é paradigmático, por conseguir uma prolongada replicação viral, mesmo na presença de repostas específicas, inicialmente vigorosas. Pensa-se que contribuem, de forma determinante, para esta fuga, factores como a destruição dos linfócitos Thelper, especificamente infectados pelo vírus, o aparecimento de algumas variantes antigénicas53 e a existência de uma cápsula capaz de manter, estruturalmente minimizado, o acesso aos seus epítopos mais importantes.54
Mais recentemente, alguns mecanismos adicionais têm sido mais bem percebidos. Uma proteína viral, que se tem demonstrado fulcral, logo no primeiro momento de evasão, interrompendo a apresentação de antigénios, é a nef.55,56,57 por outro lado, a proteína Tat acaba por ser extremamente importante contra a ação citotóxica do sistema imunitário, por alterar proteínas de membrana dos linfócitos T CD8+, fazendo que sejam estes a ficar marcados para degradação.58 Alguns polimorfismos do genoma viral são ainda capazes de tornar as células infectadas incapazes de expressar receptores ativadores da função citotóxica das células NK, que de outra forma poderiam ser importantes, ajuda contra a disseminação do vírus.59
Que consequências? – A SIDA
A consequência direta para a maior parte dos infectados pelo VIH é, assim, a Síndrome de Imunodeficiência Adquirida.
O tempo que uma pessoa infectada demora a desenvolver SIDA varia, e depende de muitos factores. Certo é que, na maioria dos casos, o sistema imunitário do hospedeiro acaba por atingir um ponto de degradação tal que permite que as portas fiquem abertas quase permanentemente, para inúmeros agentes oportunistas.
A progressiva perda de capacidade imunitária (avaliada pela contagem de linfócitos CD4+ viáveis) relaciona-se com o aumento da probabilidade de aparecimento de certos agentes infecciosos e permite-nos considerar normal a seguinte sequência: Herpes Zoster (dor e eczema), Herpes simplex (vesículas), tuberculose (febre, cansaço, hemoptises), Candidíase Oral (Odinofagia, Disfagia), Sarcoma de Kaposi (lesão cutânea e queixas gastrointestinais), pneumocistose (dispneia, dor no peito, tosse seca), toxoplasmose (enxaqueca, confusão, convulsões), Mycobacterium avium (febre, suores noturnos, diarreia), Criptosporidíase (perda ponderal, Vómitos, Diarreia).60,61
Para além das infecções que lhe podem estar associadas, a SIDA é caracterizada pelo aumento de falhas noutra ação fundamental do sistema imunitário – a luta contra o cancro. Ainda que a relação não seja direta com todos os tipos de cancro, a verdade é que a imunodeficiência causada pelo VIH já se provou associada a vários tipos de linfoma, cancro do lábio, testículo, entre outros.62
Conclusão
O complexo sistema imunitário humano protege-nos, em condições normais, de um sem-número de situações que, em última instância, nos levariam à morte. A sua evolução foi, provavelmente. Um dos mais importantes acontecimentos da história filogenética, mas trouxe também, claro, maior probabilidade de ocorrência de erros. Por todas as características mencionadas anteriormente, é fundamental estar atento a uma infecção como a do VIH, e saber distingui-la de outras que a esta se assemelhem.
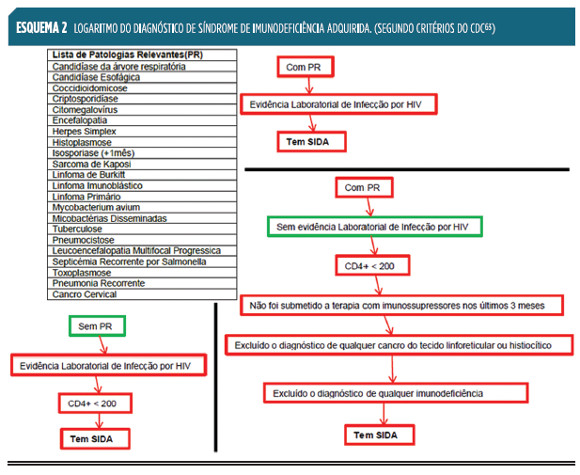
Os atuais critérios clínicos e laboratoriais do CDC (Center for Disease Control) dos EUA, que definem um caso de SIDA,63 permitem a esquematização de um diagnóstico diferencial, partindo de dados clínicos como o que está apresentado sob a forma de algoritmo no Esquema 2
.
As páginas deste artigo não são, certamente, suficientes para dizer tudo o que há de importante sobre estes vastos assuntos, mas esperamos que esta revisão possa ter relacionado um conjunto de conceitos essenciais, de forma clara e útil, a qualquer profissional de saúde.
Agradecimentos
À Dr.ª Sónia lemos, do Hospital pediátrico, do Centro Hospitalar e Universitário de Coimbra, pelo seu apoio e colaboração na revisão deste artigo.
Referências
1. Silverstein AM. A History of Immunology. 2nd ed. New York. Academic Press/Elsevier; 2009. [ Links ]
2. National Institute of Allergy and Infectious Diseases. Immune System. Available at: http://www.niaid.nih.gov/topics/immunesystem. Accessed March 6; 2012. [ Links ]
3. Kindt TJ, Osborne BA, Goldsby RA. Kuby Immunology. 6th ed. New York. W. H. Freeman & Company: 2006. [ Links ]
4. Blázquez OS, García PR, Falcón FA. Inmunodeficiencias primarias. In: Fernandez JG, Dominguez JAR, Suso JJM, Tascón AB. Manual Diagnóstico y Terapéutica en Pediatria, 5thed. Madrid. Cofás; 2009; 201-9. [ Links ]
5. Chinen J, Shearer WT. Secondary immunodeficiencies, including HIV infection. J Allergy Clin Immunol 2010;125(suppl 2):S195-S203. [ Links ]
6. Casanova JL, Abel L. Primary immunodeficiencies: a field in its infancy. Science 2007;317(5838):617-9. [ Links ]
7. Notarangelo LD. Primary immunodeficiencies. J Allergy Clin Immunol 2010;125(2 Suppl 2):S182-94. [ Links ]
8. Batalha S, Russo T, Almeida M et al. Imunodeficiências primárias – Revisão da Fisiopatologia, classificação, abordagem. Revista Saúde Infantil 2011;33(3):97-102. [ Links ]
9. Chinen J, Shearer WT. Secondary immunodeficiencies, including HIV infection. J Allergy Clin Immunol 2010;125(2 Suppl 2):S195-203. [ Links ]
10. Medzhitov R, Janeway C. Innate Immunity: The Virtues of a Nonclonal System of Recognition Cell 1997;91(3):295-8. [ Links ]
11. Male D, Brostoff J, Roth DB, Roitt I. Immunology. 7th ed. Philadelphia. Mosby Elsevier: 2006. [ Links ]
12. Helbert, M. Flesh and Bones of Immunology. 1st ed. London. Mosby Elsevier: 2006. [ Links ]
13. Errante PR, Frazão JB, Condino-Neto A. Displasia ectodérmica anidrótica com imunodeficiência. Rev Bras Alerg Imunopatol 2010; 33: 215-219 [ Links ]
14. Snyder O. Hospital Institute of Technology and Management (website). Available at: http://www.hi-tm.com/1908/SECTION-5-D-1908.pdf. Accessed May 3; 2012. [ Links ]
15. Becker JH, Wu SC. Fever – an update. J Am Podiatr Med Assoc 2010; 100(4): 281-90. [ Links ]
16. Pizzo PA. Fever in Immunocompromised Patients. N Engl J Med 1999; 341: 893-900. [ Links ]
17. Kashio M, Sokabe T, Shintaku et al. Redox signal-mediated sensitization of transient receptor potential melastatin 2 (TRPM2) to temperature affects macrophage functions. Proc Natl Acad Sci USA 2012; 109(17): 6745-50. [ Links ]
18. Mace TA, Zhong L, Kilpatrick C et al. Differentiation of CD8+ T cells into effector cells is enhanced by physiological range hyperthermia. J Leukoc Biol 2011; 90 (5): 951-62. [ Links ]
19. Gram K, Yang S, Steiner M et al. Simultaneous absence of surfactant proteins A and D increases lung inflammation and injury after allogeneic HSCT in mice. Am J Physiol Lung Cell Mol Physiol 2009; 296(2): L167-75. [ Links ]
20. Madan T, Reid KB, Clark H et al. Susceptibility of mice genetically deficient in SP-A or SP-D gene to invasive pulmonary aspergillosis. Mol Immunol 2010; 47(10): 1923-30. [ Links ]
21. Hoebe K, Janssen E, Beutler B. The interface between innate and adaptive immunity. Nat Immunol 2004; 5(10): 971-4 [ Links ]
22. Zhang SY, Jouanguy E, Ugolini S et al. TL R3 deficiency in patients with herpes simplex encephalitis. Science 2007; 317(5844): 1522-7. [ Links ]
23. Chapel H. An unusual presentation of a human TL R pathway deficiency: lessons (online presentation). Available at: http://imunologie.lf2.cuni.cz/soubory_esidworkshop/Chapel2004.pdf. Accessed May 13; 2012. [ Links ]
24. Blaese RM, Winkelstein JA, eds. Patient & Family Handbook for Primary Immunodeficiency Diseases. 4th ed. Towson. Immune Deficiency Foundation: 2007. [ Links ]
25. Chandrasoma P, Taylor C. Concise Pathology. 3rd ed. London. Appelton & Lange: 1991. [ Links ]
26. Janeway CA, Travers P, Walport M et al. Immunobiology: The Immune System in Health and Disease. 5th ed. New York. Garland Science: 2001. [ Links ]
27. Baumgarth N. A teo-phase model of B-cell activation. Immunol Rev 2000; 176: 171-80. [ Links ]
28. Pier GB, Lyczak JB, Wetzler LM. Immunology, Infection and Immunity. 1st ed. Washington DC. ASM Press; 2004. [ Links ]
29. Notarangelo L, Casanova JL, Conley ME et al. Primary immunodeficiency diseases: An update from the International Union of Immunological Societies Primary Immunodeficiency Diseases Classification Committee Meeting in Budapest, 2005. J Allergy Clin Immunol 2006; 117(4): 883-96. [ Links ]
30. Cameron MJ, Kelvin DJ. Cytokines, Chemokines and Their Receptors. In: Madame Curie Bioscience Database [Internet]. Austin (TX): Landes Bioscience; 2000-. Available at: http://www.ncbi.nlm.nih.gov/books/NBK6294. Accessed August 24; 2012. [ Links ]
31. OShea JJ, Ma A, Lipsky P. Cytokines and autoimmunity. Nat Rev Immunol 2002; 2(1): 37-45. [ Links ]
32. Kaiser, GE. The adaptive Immune System - Cell-Mediated Immunity (website). Available at: http://www.faculty.ccbcmd.edu/courses/bio141/lecguide/unit5/cellular/cmioverview/cmioverview.html. Accessed May 13, 2012. [ Links ]
33. Baratin M, Roetynk Sophie, Léopolard C et al. Natural killer cell and macrophage cooperation in MyD88-dependent innate responses to Plasmodium falciparum. Proc Natl Acad Sci USA.2005; 102(41): 14747-52. [ Links ]
34. Nedvetzki S, Sowinski S, Eagle RA. Reciprocal regulation of human natural killer cells and macrophages associated with distinct immune synapses. Blood 2007; 109(9): 3776-85. [ Links ]
35. McMurray DN. Cell-mediated immunity in nutritional deficiency. Prog Food Nutr Sci 1984; 8(3-4): 193-228. [ Links ]
36. Smythe PM, Brereton-Stiles GG , Grace HJ. Thymolymphatic deficiency and depression of cell-mediated immunity in protein-calorie malnutrition. The Lancet 1971; 298: 939-944. [ Links ]
37. Dutz W, Rossipal E, Ghavami H et al. Persistent cell mediated immune-deficiency following infantile stress during the first 6 months of life. Eur J Pediatr 1976; 122(2): 117-30. [ Links ]
38. Kumar RK, Penny R. Cell-mediated immune deficiency in Hodgkins disease. Immunology Today 1982; 3: 269-73. [ Links ]
39. Savino W, Dardenne M. Nutricional imbalances and infections affect the thymus: consequences on T-cell-mediated immune responses. Proc Nutr Soc 2010; 69(4): 636-43. [ Links ]
40. Abbas AK, Lichtman AH. Basic Immunology: Functions and Disorders of the Immune System. 2nd ed. Philadelphia. Saunders: 2006. [ Links ]
41. Feldhahn N, Río P, Soh BNB et al. Deficiency of Brutons tyrosine in B cell precursor leukemia cells. Proc Natl Acad Sci USA 2005; 102(37): 13266-71. [ Links ]
42. Fleischman A, Hershfield MS, Toutain S et al. Adenosine Deaminase Deficiency and Purine Nucleoside Phosphorylase Deficiency in Common Variable Immunodeficiency. Clin Diagn Lab Immunol 1998; 5(3): 399-400 [ Links ]
43. Mombaerts P, Iacomini J, Johnson RS et al. RAG-1-deficient mice have no mature B and T lymphocytes. Cell 1992; 68(5): 869-77. [ Links ]
44. Decaluwe H, Taillardet M, Corcuff E et al. Gamma(c) deficiency precludes CD8+ T cell memory despite formation of potent T cell effectors. Proc Natl Acad Sci U S A 2010; 107(20): 9311-6. [ Links ]
45. National Institute of Allergy and Infectious Diseases. Stucture of HIV. Available at: http://www.niaid.nih.gov/topics/HIVAIDS/Understanding/Biology/Pages/structure. Accessed May 6; 2012. [ Links ]
46. Taveira N. Etiologia da infecção VIH/SIDA. In: Infecção VIH/SIDA. 1st ed. V. N. Famalicão; 2004; 39-52. [ Links ]
47. Otero JJ. SIDA: mecanismos de transmisión y su prevención. In: Infecção VIH/SIDA. 1st ed. V. N. Famalicão; 2004; 83-100. [ Links ]
48. Toledo Jr AC, Ribeiro FA, Ferreira FG et al. Conhecimento, atitudes e comportamentos frente ao risco ocupacional de exposição ao HIV entre estudantes de Medicina da Faculdade de Medicina da Universidade Federal de Minas Gerais. Rev Soc Bras Med. 1999; 32(5). [ Links ]
49. Geijtenbeek TB, Kwon DS, Torensma R et al. DC-SIGN , a Dendritic Cell-Specific HIV-1-Binding Protein that Enhances trans-Infection of T Cells. Cell 2000; 100: 587-97. [ Links ]
50. Garg H, Blumenthal R. The Role of HIV Gp41 Mediated Fusion/Hemifusion in Bystander Apoptosis. Cell Mol Life Sci 2008; 68(20): 3134-44. [ Links ]
51. Levy JA. Infection by Human Imunodeficiency Virus – CD4 is not enough. N Engl J Med 1996; 335(20): 1528-30. [ Links ]
52. Caetano JA. Imunopatologia da infecção pelo VIH. In: Infecção VIH/SIDA. 1st ed. V. N. Famalicão; 2004; 53-66. [ Links ]
53. Piquet V, Trono D. Living in oblivion: HIV immune evasion. Semin Immunol 2011; 13(1): 51-7. [ Links ]
54. Johnson WE, Desroisiers RC. Viral Persistence: HIVs Strategies of Immune System Evasion. Annu Rev Med 2002; 53: 499-518. [ Links ]
55. Kasper MR, Roeth JF, Williams M et al. HIV-1 Nef disrupts antigen presentation early in the secretory pathway. J Biol Chem 2005; 280(13): 12840-8. [ Links ]
56. Yi L, Rosales T, Rose JJ et al. HIV-1 Nef Binds a Subpopulation of MHC-I throughout Its Trafficking Itinerary and Down-regulates MHC-I by perturbing both Anterograde and Retrograde Trafficking. J Biol Chem 2010; 285(40):30884-905. [ Links ]
57. Xu W, Santini PA, Sullivan JS et al. HIV-1 evades virus-specific IgG2 and IgA class switching by targeting systemic and intestinal B cells via long-range intercellular conduits. Nat Immunol 2009; 10(9): 1008-17. [ Links ]
58. Faller EM, Sugden SM, McVey MJ et al. Soluble HIV Tat Protein Removes the IL-7 Receptor a-Chain from the Surface of Resting CD8 T Cells and Targets It for Degradation. J Immunol. 2010; 185(5):2854-66. [ Links ]
59. Alter G, Heckerman D, Schneidewind A et al. HIV-1 adaptation to NK cell mediated immune pressure. Nature 2011; 476(7358):96-100. [ Links ]
60. AIDS.gov. Opportunistic infections. Available at: http://aids.gov/hiv-aids-basics/staying-healthy-with-hiv-aids/potential-related-health-problems/opportunistic-infections. Accessed May 20; 2012. [ Links ]
61. Ong EL. Common AIDS-associated opportunistic infections. Clin Med 2008; 8(5): 539-43. [ Links ]
62. Frisch M, Biggar RJ, Engels EA et al. Association of cancer with AIDS-related immunosuppression in adults. JAMA 2001; 285(13): 1736-45. [ Links ]
63. CDC. 1993 revised classification system for HIV infection and expanded surveillance case definition for AIDS among adolescents and adults. MMWR Recomm Rep 1992; 41: 1-19. [ Links ]
64. Maek-A-Nantawat W, Buranapraditkun S et al. Increased interleukin-17 production both in helper T cell subset Th17 and CD4-negative T cells in human immunodeficiency virus infection. Viral Immunology 2007; 20(1): 66-75. [ Links ]
Rui Soares
Faculdade de Medicina da Universidade de Coimbra Microbiologia, sala 19 Rua larga, 3004-504 Coimbra. E-mail: rui.soares@uc.pt
Data de recepção / reception date: 05/08/2013
Data de aprovação / approval date: 03/12/2013

