










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista Portuguesa de Imunoalergologia
versão impressa ISSN 0871-9721
Rev Port Imunoalergologia vol.24 no.3 Lisboa set. 2016
ARTIGO REVISÃO
A tropomiosina como um panalergénio: Revisão
Tropomyosin as a panallegen: Review
Joana Cosme1, Amélia Spínola Santos1, Manuel Pereira Barbosa1,2
1 Serviço de Imunoalergologia, Hospital de Santa Maria, Centro Hospitalar Lisboa Norte, Lisboa, Portugal
2 Clinica Universitária de Imunoalergologia, Faculdade de Medicina de Lisboa, Lisboa, Portugal
RESUMO
A tropomiosina é um panalergénio envolvido em reações de reatividade cruzada essencialmente entre espécies de invertebrados, sendo particularmente importante, em termos epidemiológicos, a reatividade cruzada entre os ácaros do pó doméstico e os crustáceos. Tendo por base uma revisão bibliográfica sobre o tema da hipersensibilidade à tropomiosina, apresenta‑se uma descrição da estrutura e da função desta proteína; de seguida enumeram‑se as várias tropomiosinas referidas na literatura e indica‑se o grau de homologia existente entre a tropomiosina do camarão e as tropomiosinas de outras fontes alergénicas. Neste trabalho faz‑se ainda uma revisão dos principais estudos sobre a prevalência de sensibilização à tropomiosina onde se verifica que a percentagem de sensibilização à tropomiosina varia de acordo com a fonte proteica e região geográfica consideradas. Por último, descrevem‑se os resultados de alguns trabalhos que destacam a importância da utilização da IgE específica para a tropomiosina como uma ferramenta diagnóstica adicional, in vitro, no diagnóstico de alergia ao camarão.
Palavras‑chave: Panalergénio, reatividade cruzada, sensibilização, tropomiosina, valor diagnóstico.
ABSTRACT
Tropomyosin is a panallergen involved in cross‑sensitivity reactions between, invertebrates, being of particular epidemiological importance, the cross‑reactivity between house dust mites and crustaceans. Based on a literature review about tropomyosin hypersensitivity, firstly there is made a description of the structure and function of this protein and afterwards the tropomyosin allergens described in the literature as well as the degree of homology between the shrimp tropomyosin and tropomyosin from other allergen sources are presented. The results of the main studies about tropomyosin sensitization prevalence which shows that tropomyosin sensitization varies according with the protein source and with the geographical area considered are also revised.
Finally, there are presented the results of some studies that emphasize the importance of using in vitro determination of tropomyosin specific IgE as an additional diagnostic tool in shrimp allergy diagnosis.
Key‑words: Cross‑reactivity, diagnosis tool, panallergen, sensitization, tropomyosin.
INTRODUÇÃO
O termo panalergénio é composto pelo prefixo grego pan, que significa todo, e pela palavra alergénio1. Assim, refere‑se a um grupo de proteínas, amplamente distribuídas pela natureza e que partilham, entre si, semelhanças estruturais e de função.
Os panalergénios são alergénios de reatividade cruzada, mediada pela IgE, entre uma grande variedade de fontes alergénicas relacionadas ou não entre si2,3,4,5.
Na base destes fenómenos de reatividade cruzada parece estar a homologia estrutural entre as moléculas2,3.
São exemplos de panalergénios as profilinas, as polcalcinas, as proteínas transportadoras de lípidos (LTP), as tropomiosinas2,3, entre outros.
As tropomiosinas são proteínas estruturais presentes em todas as células eucarióticas e que fazem parte da constituição dos ácaros, dos moluscos, dos insetos e dos nemátodos6.
A tropomiosina é reconhecida como alergénio major do camarão há mais de duas décadas, existindo trabalhos publicados desde 19817. Em 1994, Witteman et al8 descreveu a tropomiosina como um alergénio dos ácaros do pó doméstico. Atualmente sabe‑se que a tropomiosina faz parte da constituição de todos os invertebrados, sendo que as semelhanças aminoacídicas entre as tropomiosinas destes, associadas a idênticas conformações estruturais, parecem estar na base da reatividade cruzada entre os ácaros do pó doméstico e outros invertebrados, como o camarão, o caracol ou a barata, espécies taxonomicamente não relacionadas4,6.
TROPOMIOSINA – ESTRUTURA E FUNÇÃO
A tropomiosina pertence a uma família de proteínas altamente conservadas durante a evolução das espécies, com elevada estabilidade térmica, resistentes ao pH gástrico e às peptidases gastroentéricas e está presente quer nas células musculares quer nas não musculares dos vertebrados e dos invertebrados5,9.
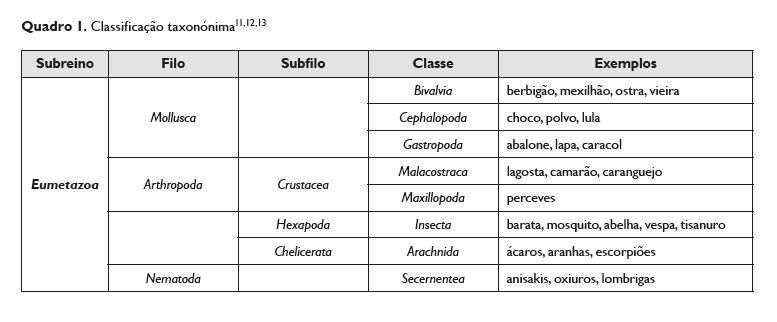
A tropomiosina dos vertebrados não é alergénica, pelo contrário, nos invertebrados, como nos crustáceos (como o camarão, o caranguejo e a lagosta), nos nemátodos (como no Anisakis simplex ou no Ascaris lumbricoides), nos aracnídeos (como os ácaros), nos insetos (como na barata) ou nos moluscos (como a lula, o caracol, o mexilhão e a ostra) é alergénica4,9,10. Estes invertebrados pertencem ao reino Animalia e ao subreino Eumetazoa.
O Quadro 1 apresenta, de forma breve, uma classificação taxonómica onde os invertebrados referidos se enquadram.
Estruturalmente a tropomiosina consiste numa molécula composta por duas cadeias paralelas, em alfa hélice, enroladas em torno uma da outra, formando um dímero em espiral6,10, sendo o seu peso molecular médio de 37 kDa5.
Nas células musculares, a tropomiosina, juntamente com a actina e a miosina, participa na atividade contráctil das células. A função da tropomiosina nas células não musculares não é totalmente conhecida, embora se acredite que participe na regulação da morfologia e motilidade celulares10.
Apesar das tropomiosinas serem estruturalmente homólogas entre si, foram identificadas diferentes isoformas desta em diferentes espécies, tecidos e células6,10.
Por exemplo, no que se refere à tropomiosina do camarão, foram identificadas 8 epitopos em 5 partes distintas da molécula14.
TROPOMIOSINA COMO ALERGÉNIO
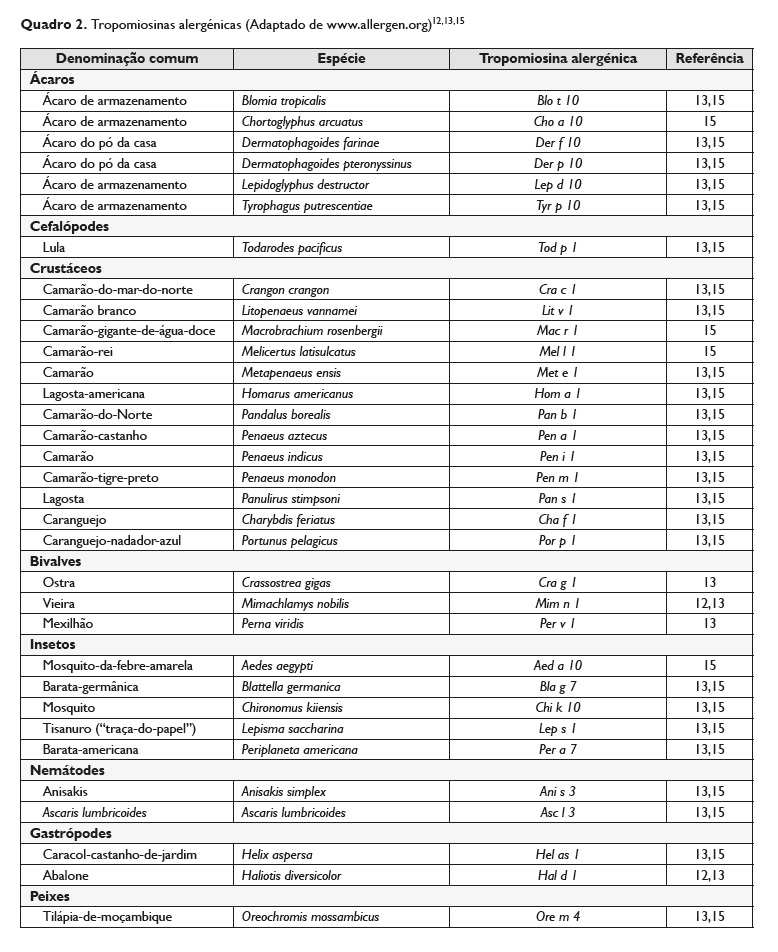
Na lista de nomenclatura alergénica da OMS/IUIS15 (WHO/IUIS Allergen Nomenclature Sub‑Committee, disponível em http://www.allergen.org) existem, à data da elaboração da presente revisão, 29 referências para a tropomiosina.
Para além destas existem, ainda, descritas na literatura, outras tropomiosinas (Quadro 2)12,13.
A primeira descrição da tropomiosina como alergénio major do camarão data de 1981 e é feita por Hoffman et al7. Em 1989, foi identificado na espécie Paneus indicus um alergénio correspondente a uma proteína resistente ao calor designada por Sa‑II, mais tarde renomeada Pen i 116 de acordo com a nomenclatura standard dos alergénios. Em 1993, Daul et al demonstraram que o alergénio major do camarão é uma proteína com 36 kDa e que está presente quer no corpo do camarão (cru e cozido) quer no líquido de cozedura do camarão, quando este é cozinhado17. Estes achados são corroborados pelos trabalhos de Shanti et al (1993)18 e de Leung et al (1994)19.
A tropomiosina (Pen a 1) é o alergénio major do camarão- castanho (Penaeus aztecus), estando presente quer nas formas cruas quer nas cozidas do camarão17 ou no vapor libertado da sua cozedura, estando sensibilizados a esta proteína cerca de 82% dos doentes com alergia ao camarão10,21,22.
Outros estudos indicam, ainda, a tropomiosina como um alergénio presente nos crustáceos e nos moluscos23,24.
Por exemplo, em 1998 o grupo de Leung et al publica alguns trabalhos onde a tropomiosina é descrita como sendo o alergénio major da lagosta (Pan s 1)25 e do caranguejo (Cha f 1)26. No caso do caranguejo, de acordo com os autores, a tropomiosina é uma proteína de 34 kDa com elevada homologia com a tropomiosina do camarão Metapenaeus ensis (Met e 1)25,26. No que se refere aos moluscos, o papel da tropomiosina também é reconhecido como alergénio, por exemplo na lula (Miyazawa et al1996)27.
Em relação aos cefalópodes, por exemplo, em 2006 Motoyama et al25 demonstraram que a tropomiosina é, também, o alergénio major deste grupo. Para além disto, evidenciaram que as tropomiosinas dos cefalópodes apresentam elevada identidade de sequência (mais de 92 %) entre si, podendo este facto estar na base da reatividade cruzada entre os cefalópodes28.
Também Emoto et al (2009)29 descreveram que a tropomiosina é o alergénio major de uma grande variedade de gastrópodes e de bivalves. Para além disto, estes autores corroboraram ainda as conclusões de Lehrer & McCants (1987)30, Leung et al (1996)31 e Motoyama et al (2006)28 ao defenderem que a tropomiosina participa nos fenómenos de reatividade cruzada entre estes e entre os cefalópodes e os crustáceos.
Moreno‑Escobosa et al (2002)32 e Marinho et al (2005)33 descrevem a tropomiosina como o alergénio implicado provavelmente na alergia aos perceves e na reatividade cruzada entre estes crustáceos com os ácaros.
Há, ainda, relatos do papel da tropomiosina na alergia aos caracóis34,35 e na alergia a parasitas, como é o caso do Anisakis simplex36,37.
Para a classe dos insetos, sabe‑se que, por exemplo na barata-americana (Periplaneta americana) e na barata-germânica (Blattella germanica), as tropomiosinas, respectivamente Per a 7 e Bla g 7, são alergénios importantes na reatividade cruzada com outras espécies38. Por exemplo, Per a 7 com os ácaros39 e Per a 7 com o camarão40.
Os autores descrevem, também, uma homologia na sequência aminoacídica 80%, 81% e 82%, respectivamente entre a tropomiosina Per a 7 e as tropomiosinas do Dermatophagoides pteronyssinus, do Dermatophagoides farinae e do camarão Metapenaeus ensis (Met e 1)39.
Nos ácaros do pó doméstico, o grupo 10 foi atribuído ao grupo das tropomiosinas alergénicas. Assim, por exemplo, a tropomiosina do Dermatophagoides pteronyssinus é designada Der p 1041,42. A tropomiosina (Der p 10) é um alergénio minor dos ácaros embora se pense que o elevado grau de homologia entre esta e as tropomiosinas do marisco ou dos caracóis possa estar na base de reacções alérgicas sistémicas graves43.
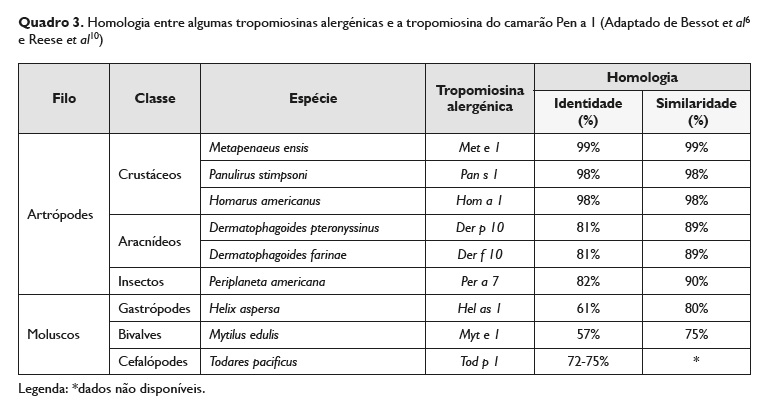
No Quadro 3 está indicada, de forma resumida, a homologia entre a tropomiosina de algumas espécies com a tropomiosina do camarão (Pen a 1). Neste Quadro os autores consideram a identidade (comparação da percentagem de aminoácidos idênticos em ambas as sequências) e a similaridade (comparação da percentagem de aminoácidos que são idênticos ou que pertencem ao mesmo grupo de aminoácidos em ambas as sequências) entre as tropomiosinas6,10.
SENSIBILIZAÇÃO A TROPOMIOSINA
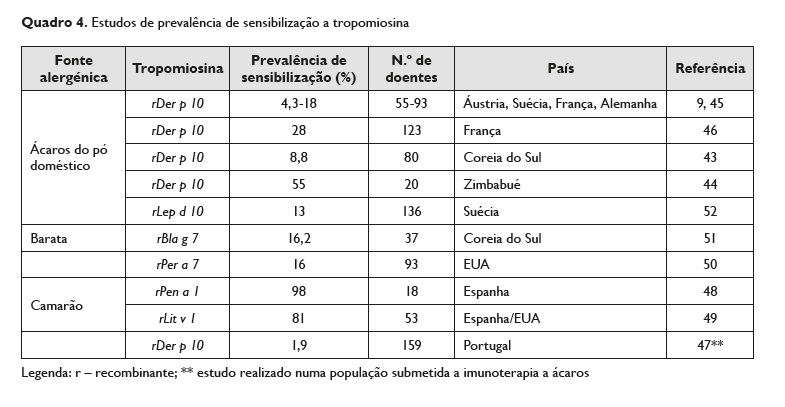
A prevalência de sensibilização à tropomiosina apresenta valores muito distintos na literatura.
No que se refere aos ácaros, as taxas de sensibilização à tropomiosina dos ácaros são, geralmente, baixas, com valores que variam ligeiramente de acordo com a distribuição geográfica. Por exemplo, em 2015, Kim et al43 demonstrou uma prevalência de 8,8% de sensibilização a rDer p 10 em doentes coreanos alérgicos a ácaros do pó doméstico43. Em África as taxas de sensibilização publicadas são, geralmente, mais elevadas.
Por exemplo, Westritschnig et al, determinaram uma taxa de sensibilização de 55% em doentes do Zimbabué44.
Na Europa, as taxas de sensibilização in vitro são geralmente mais baixas, com valores, por exemplo, de 18% em doentes da Áustria e na Suécia, de 9% e 10%, respetivamente, na França e na Itália45, ou de 4,3% na Alemanha9. Um trabalho francês, da região de Marselha (Sul de França), de Bronnert et al, mostra uma percentagem geral de sensibilização a rDer p 10 ligeiramente mais elevada, com valor de sensibilização a Der p 10 de 28% na população estudada (n=123). Estes autores determinaram percentagens relativas de 30% e 25% respetivamente para as crianças e adultos e demonstraram que, na sua população de doentes com alergia a ácaros, os doentes com níveis mais elevados de IgE específica para rDer p 10 apresentavam clínica de alergia aos mariscos, ao contrário dos alérgicos a ácaros, com níveis mais baixos de IgE específica para rDer p 1046.
Em Portugal, Pereira dos Reis et al identificaram uma prevalência de sensibilização à tropomiosina do camarão numa amostra de 159 doentes alérgicos a Dermatophagoides pteronyssinus e/ou Dermatophagoides farinae de 1,9%, de acordo com os valores de IgE específica para tropomiosina (método Immunocap Pharmacia Diagnostics®) em doentes submetidos a imunoterapia a ácaros47.
No que se refere ao camarão Penaeus aztecus, o único alergénio major identificado é a tropomiosina Pen a 1, presente em pelos menos 82% dos doentes com alergia ao camarão21,22. Gámez et al (2011) confirmaram, num grupo de 18 doentes alérgicos ao camarão, que todos (100%) apresentavam testes cutâneos em picada para camarão e que 98% destes doentes apresentavam IgE específica positiva para rPen a 148.
Ayuso et al, em 2010, também determinaram uma frequência de sensibilização à tropomiosina do camarão Litopenaeus vannamei (rLit v 1) de 81%, sendo que a taxa de sensibilização era superior (94 %) no grupo das crianças ao dos adultos (61%) com alergia a camarão49.
No que se refere a barata, a tropomiosina é considerada um alergénio minor em países como EUA50 ou Coreia do Sul (taxa de sensibilização a rBla g 7 de 16,2%)51.
No Quadro 4 estão indicados, de forma resumida, os dos estudos referidos, onde se determinou a prevalência de sensibilização a tropomiosina.
Analisando os valores de sensibilização apresentados no Quadro 4 e no que se refere, por exemplo, à fonte alergénica, constata‑se que a prevalência de sensibilização a Der p 10 apresentada nos estudos apresenta valores muito próximos, com exceção do estudo realizado no Zimbabué onde se identificou uma frequência de sensibilização mais elevada (55%). Esta diferença pode ser justificada não só pelo reduzido tamanho da amostra estudada, como também pelas diferenças geográficas. Já no que se refere aos estudos que consideraram como fonte alergénica o camarão, tanto Gámez et al48 como Ayuso et al49 descrevem nos seus trabalhos frequências de sensibilização similares. Por outro lado, Pereira dos Reis et al47 identifica uma prevalência de apenas 1,9% na população portuguesa. Esta diferença pode dever‑se, contudo, ao facto de a amostra considerada neste trabalho ser uma população de doentes com alergia a ácaros submetida a imunoterapia. Importa realçar, todavia, que os autores portugueses demonstraram que a realização de imunoterapia a ácaros não foi fator de risco para aumento da sensibilização a tropomiosina.
Nos estudos que consideraram a barata como fonte alergénica, apesar das metodologias utilizadas nos estudos terem sido distintas, já que enquanto Jeong et al51 determina uma sensibilização a rBla g 7 através do método ELISA, Satinover et al50 fazem‑no através do método InmunoCAP® de estreptavidina, as frequências encontradas são semelhantes.
PODE A TROPOMIOSINA SER UM MARCADOR DIAGNÓSTICO ADICIONAL NA ALERGIA AO CAMARÃO?
A investigação diagnóstica da alergia ao camarão deve incluir uma história clínica, realização de testes cutâneos em picada, o doseamento de IgE específicas séricas e a realização de uma prova de provocação oral.
As provas de provocação oral continuam a ser o Gold standard diagnóstico53,54.
Apesar disto e, mesmo adotando metodologias e protocolos de atuação estandardizados, as provas de provocação oral são um procedimento diagnóstico dispendioso, com uma realização morosa e não isentas de induzir sintomas clínicos graves48,54.
Assim, nesta perspetiva, o doseamento de anticorpos IgE específicos para componentes de proteínas de determinados alimentos tem mostrado ser uma ferramenta diagnóstica alternativa promissora, cada vez mais utilizada na prática clínica51. No que se refere, por exemplo, à alergia ao camarão, vários são os alergénios identificados.
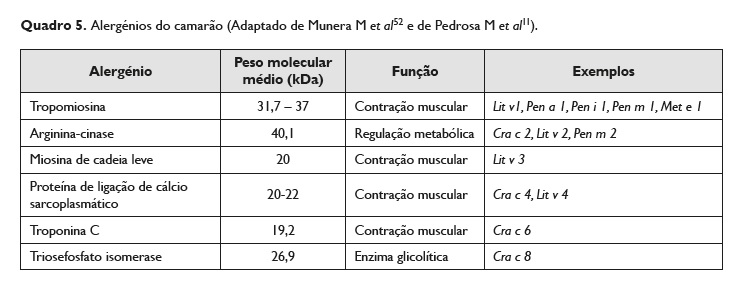
No Quadro 5 estão indicados os alergénios mais bem caracterizados na literatura, sendo que, para além destes, existem outros menos frequentemente descritos, como a homocianina, as proteínas de união aos ácidos gordos, a miosina de cadeia pesada, a α‑actina, entre outros 55,11.
Com o avanço científico dos últimos anos nos métodos de diagnóstico in vitro, a utilização da tropomiosina recombinante do camarão é defendida por alguns autores como teste diagnóstico alternativo na alergia ao camarão6 com maior sensibilidade e especificidade do que os testes cutâneos em picada com extrato total de camarão na deteção de alergia ao marisco56.
Antes do advento do estudo molecular, realizavam‑se sobretudo testes cutâneos prick‑prick. Utilizando este teste, Jirapongsananuruk et al57 descreveram que pápulas com um diâmetro entre 20 a 30 mm nos testes cutâneos em picada realizados com extratos comerciais e naturais de camarão, juntamente com a utilização de testes cutâneos em picada com alimento em natureza (prick‑prick), têm 95% de probabilidade de prever um resultado positivo numa prova de provocação oral em doentes tailandeses com alergia ao camarão-tigre-castanho (Penaeus monodon) e ao camarão-gigante-de-água-doce (Macrobrachium rosenbergii). Para além disto, estes autores determinaram que o teste cutâneo em picada utilizando extrato de camarão (Center Laboratory, Port Washington, NY) apresenta uma sensibilidade de 88,33%, uma especificidade de 37,50%, com valores preditivos positivo e negativo, respetivamente, de 91,38% e de 30%, no diagnóstico de alergia a estas espécies de camarão57.
Gámez et al48 determinaram num grupo de doentes espanhóis, que o doseamento da IgE específica para rPen a 1 apresenta uma sensibilidade de 88%, uma especificidade de 77% com valores preditivos positivo de 72% e negativo de 91%. Perante estes valores, os autores advogam que a determinação da IgE específica a rPen a 1 fornece um valor diagnóstico adicional aos TCP e à determinação da IgE específica para o camarão na confirmação diagnóstica de alergia ao camarão em doentes europeus.
Contudo, e tendo em conta que o valor de especificidade não é muito elevado, na opinião destes autores a prova de provocação oral deva ser considerada em alguns casos48.
Yang et al58 também compararam a sensibilidade e especificidade diagnóstica da determinação da IgE específica para rPen a 1 com a da IgE específica para extrato completo de camarão e com a utilização de testes cutâneos em picada utilizando extrato de camarão (Hollister‑Stier Laboratories, Spokane, Wash).
De acordo com estes autores, os três métodos têm valores de sensibilidade diagnóstica semelhantes, mas com valores de especificidade distintos (especificidade da IgE para tropomiosina de camarão de 92,8%, especificidade da IgE específica para o camarão de 75% e especificidade dos testes cutâneos em picada de 64,2%). No mesmo estudo, os autores determinaram a eficiência diagnóstica dos testes definida pelos mesmos como a proporção de verdadeiros positivos e verdadeiros negativos detetados pelos estes, sendo o valor de eficiência diagnóstica da determinação da IgE específica para a tropomiosina superior à determinação da IgE específica para o camarão e à dos testes cutâneos em picada (88,5%, 74,2%, e 65,7%, respetivamente)58.
Assim, à semelhança de Gámez et al, estes autores defendem que a determinação da IgE específica para a tropomiosina do camarão tem um valor diagnóstico adicional na alergia ao camarão48,58.
Não se conhecem estudos que indiquem valores de cut‑off precisos para a IgE específica da tropomiosina do camarão no diagnóstico de alergia ao camarão ou aos crustáceos, como está descrito para o leite de vaca, ovo, trigo ou amendoim48,58.
CONCLUSÃO
A tropomiosina é um panalergénio amplamente distribuído pela natureza e importante na reatividade cruzada entre alimentos e aeroalergénios de origem animal, tais como camarão e ácaros. É, por exemplo, um alergénio major do camarão e minor nos ácaros do pó doméstico.
A percentagem de sensibilização à tropomiosina varia de acordo com a fonte proteica considerada e de acordo com a região geográfica. Para além disso, as metodologias utilizadas, os critérios de inclusão considerados e as populações incluídas em cada estudo podem também interferir nas diferenças encontradas entre as prevalências de sensibilização. Assim sendo, o conhecimento das suas características estruturais revela ser de extrema importância para uma melhor compreensão da sua alergenicidade. Poucos são, todavia, os estudos levados a cabo com o objetivo de determinar a sensibilização às tropomiosinas de diferentes fontes alergénicas.
A produção de tropomiosina recombinante tem revelado ser uma ferramenta diagnóstica importante, com valor adicional no diagnóstico de alergia ao camarão.
REFERÊNCIAS
1. Hauser M, Roulias A, Ferreira F, Egger M. Panallergens and their impact on the allergic patient. Allergy Asthma Clin Immunol 2010;6:1‑14. [ Links ]
2. Migueres M, Dávila I, Frati F, Azpeitia A, Jeanpetit Y, Lheritier‑Barrand M, Incorvaia C, et al. Types of sensitization to aeroallergens: Definitions, prevalences and impact on the diagnosis and treatment of allergic respiratory disease. Clin Transl Allergy 2014; 4:1‑8. [ Links ]
3. Moreno‑Aguilar C. Improving pollen immunotherapy: minor allergens and panallergens. Allergol Immunopathol (Madr) 2008; 36:26‑30. [ Links ]
4. Werfel T, Asero R, Ballmer‑Weber BK, Beyer K, Enrique E, Knulst AC, et al. Position paper of the EAACI: food allergy due to immunological cross‑reactions with common inhalant allergens. Allergy 2015; 70:1079‑90. [ Links ]
5. Popescu FD. Cross‑reactivity between aeroallergens and food allergens. World J Methodol 2015; 5:31‑50. [ Links ]
6. Bessot JC, Metz‑Favre C, Rame JM, De Blay F, Pauli G. Tropomyosin or not tropomyosin, what is the relevant allergen in house dust mite and snail cross allergies?. Eur Ann Allergy Clin Immunol 2010; 42:3‑10. [ Links ]
7. Hoffman DR, Day ED, Miller JS. The major heat stable allergen of shrimp. Ann Allergy 1981; 47:17-22. [ Links ]
8. Witteman AM, Akkerdaas JH, van Leeuwen J, van der Zee JS, Aalberse RC. Identification of a cross reactive allergen (presumably tropomyosin) in shrimp, mite and insects. Int Arch Allergy Immunol 1994; 105: 56‑61. [ Links ]
9. Becker S, Gröger M, Canis M, Pfrogner E, Kramer MF. Tropomyosin sensitization in house dust mite allergic patients. Eur Arch Otorhinolaryngol 2012; 269:1291‑6. [ Links ]
10. Reese G, Ayuso R, Lehrer SB. Tropomyosin: an invertebrate pan‑allergen. Int Arch Allergy Immunol 1999; 119:247‑58. [ Links ]
11. Pedrosa M, Boyano‑Martinez T, Gracia‑Ara C, Quirce S. Shellfish Allergy: A comprehensive review. Clinic Rev Allerg Immunol 2015; 49:203‑16. [ Links ]
12. Woo CK, Bahna SL. Not all shellfish allergy is allergy! Clin Transl Allergy 2011;10;1‑3. [ Links ]
13. Gonzalez‑Fernandez J, Rodero M, Daschner A, Cuéllar C. New insights into the allergenicity of tropomyosin: a bioinformatics approach. Mol Biol Rep 2014; 41:6509‑17. [ Links ]
14. Reese G, Schicktanz S, Lauer I, et al. Structural, immunological and functional properties of natural recombinant Pen a 1, the major allergen of Brown Shrimp, Penaeus aztecus. Clin Exp Allergy 2006; 36: 517‑23. [ Links ]
15. Allergen.org [homepage on the Internet]. University of Nebraska‑Lincoln, USA: WHO/IUIS Allergen Nomenclature Sub‑committee [updated on 2015‑07‑13; cited 2015‑12‑02]. Available from: http://www.allergen.org/. [ Links ]
16. Nagpal S, Rajappa L, Metcalfe DD, Rao PV. Isolation and characterization of heat‑stable alergens from shrimp (Penaeus indicus). J Allergy Clin Immunol 1989; 83:26-36. [ Links ]
17. Daul, CB, Morgan, JE, Lehrer, SB. Hypersensitivity reactions to crustacea and mollusks. Clin Rev Allergy 1993a; 11:201-22. [ Links ]
18. Shanti KN, Martin BM, Nagpal S, Metcalfe DD, Subba Rao PV. Identification of tropomyosin as the major shrimp allergen and characterization of its IgE‑binding epitopes. J Immunol 1993; 151: 5354‑63. [ Links ]
19. Leung, PSC, Chu, KH, Chow, WK, Aftab, A, Bandea, CI, Kwan, HS, Nagy, SM, Gershwin, ME. Cloning, expression, and primary structure of Metapenaeus ensis tropomyosin, the major heat‑stable shrimp allergen. J Allergy Clin Immunol 1994; 92:837‑845. [ Links ]
20. Reese G, Tracey D, Daul CB, Lehrer SB. IgE and monoclonal antibody reactivities to the major shrimp allergen Pen a 1 (tropomyosin) and vertebrate tropomyosins. Adv Exp Med Biol 1996; 409:225‑30 [ Links ]
21. Daul, CB, Slattery, M, Reese, G, Lehrer, SB. Identification of the major brown shrimp (Penaeus aztecus) as the muscle protein tropomyosin. Int Arch Allergy Clin Immunol 1994; 105:49-55. [ Links ]
22. Reese G, Jeoung BJ, Daul CB, Lehrer SB. Characterization of recombinant shrimp allergen Pen a 1 (tropomyosin). Int Arch Allergy Immunol 1997; 113:240‑2. [ Links ]
23. Taylor SL. Molluscan shellfish allergy. Adv Food Nutr Res 2008; 54:139‑77. [ Links ]
24. Chu KH, Wong SH, Leung PS. Tropomyosin is the major mollusk allergen: reverse transcriptase polymerase chain reaction, expression and IgE reactivity. Mar Biotechnol 2000; 2:499-509. [ Links ]
25. Leung PS, Chen Y, Gershwin MR, Wong, SH, Kwan, HS, Chu, KH. Identification and molecular characterization of Charybdis feriatus tropomyosin, the major crab allergen. J Allergy Clin Immunol 1998; 102:847-52. [ Links ]
26. Leung PS, Chen YC, Mykles DL, Chow WK, Li CP, Chu KH. Molecular identification of the lobster muscle protein tropomyosin as a seafood allergen. Mol Mar Biol Biotechnol 1998, 7:12-20. [ Links ]
27. Miyazawa H, Fukamachi H, Inagaki Y, Reese G, Daul CB, Lehrer SB, Inouye S, Sakaguchi M. Identification of the first major allergen of a squid (Todarodes pacificus). J Allergy Clin Immunol 1996; 98:948-53. [ Links ]
28. Motoyama K, Ishizaki S, Nagashima Y, Shiomi K. Cephalopod tropomyosins: identification as major allergens and molecular cloning. Food Chem Toxicol 2006; 44:1997‑02. [ Links ]
29. Emoto A, Ishizaki S, Shiomi K. Tropomyosins in gastropods and bivalves: Identification as major allergens and amino acid sequence features. Food Chemistry 2009; 114: 634-41. [ Links ]
30. Lehrer SB, McCants ML. Reactivity of IgE antibodies with crustacean and oyster allergens. Evidence for common antigenic structures. Journal of Allergy and Clinical Immunology 1987; 80: 133-139. [ Links ]
31. Leung PS, Chow WK, Duffey S, Kwan HS, Gershwin ME, Chu K. IgE reactivity against a cross‑reactive allergen in Crustacea and Mollusca: Evidence for tropomyosin as the common allergen. Journal of Allergy and Clinical Immunology 1996; 98: 954-961 [ Links ]
32. Moreno Escobosa MC, Alonso LE, Sanchez AA, et al. Barnacle hypersensitivity. Allergol Immunopathol (Madr) 2002; 30:100‑3. [ Links ]
33. Marinho S, Gaspar A, Morais Almeida M, Postigo I, Guisantes J, Martínez J et al. Alergia a perceves no contexto da síndrome acaros‑crustaceos‑moluscos‑baratas. Rev Port Imunoalergologia 2005; 13:187‑93. [ Links ]
34. Ishikawa M, Ishida M, Shimakura K, Nagashima Y, Shiomi K. Purification and IgE‑binding epitopes of a major allergen in the gastropod Turbo cornutus. Biosci Biotechnol Biochem 1998; 62(7):1337‑43. [ Links ]
35. Asturias JA1, Eraso E, Arilla MC, Gomez‑Bayon N, Inácio F, Martínez A. Cloning, isolation, and IgE‑binding properties of Helix aspersa (brown garden snail) tropomyosin. Int Arch Allergy Immunol 2002 Jun;128(2):90‑6. [ Links ]
36. Guarneri F, Guarneri C, Benvenga S. Cross‑reactivity of Anisakis simplex: possible role of Ani s 2 and Ani s 3. Int J Dermatol 2007; 46:146‑50. [ Links ]
37. Asturias JA, Eraso E, Moneo I, Martinez A. Is tropomyosin na allergen in Anisakis? Allergy 2000; 55:898‑9. [ Links ]
38. Pomés A, Arruda LK. Investigating cockroach allergens: aiming to improve diagnosis and treatment of cockroach allergic patients Methods 2014; 66:75‑85. [ Links ]
39. Santos ABR, Tobias KR, Ferriani VPL, Rizzo MC, Naspitz CK, Pomes A, et al. Identification of tropomyosin from Periplaneta americana as a major cockroach allergen. J Allergy Clin Immunol 1999;103(1):S122. [ Links ]
40. Asturias JA, Gomez‑Bayon N, Arilla MC, Martinez A, Palacios R, Sanchez‑Gascon F, Martinez J. Molecular characterization of American cockroach tropomyosin (Periplaneta Americana allergen 7), a cross‑reactive allergen. J Immunol 1999;162(7): 4342‑8. [ Links ]
41. Aki T, Kodama T, Fujikawa A, Miura K, Shigeta S, Wada T, et al. Immunochemical characterization of recombinant and native tropomyosins as a new allergen from the house dust mite, Dermatophagoides farinae. J Allergy Clin Immunol 1995; 96:74-83. [ Links ]
42. Shafique RH, Inam M, Ismail M, Chaudhary FR. Group 10 allergens (tropomyosins) from house‑dust mites may cause covariation of sensitization to allergens from other invertebrates. Allergy Rhinol (Providence) 2012;3: e74‑90. [ Links ]
43. Kim HS, Kang SH, Won S, Lee EK, Chun YH, Yoon JS, et al. Immunoglobulin E to allergen components of house dust mite in Korean children with allergic disease. Asia Pac Allergy 2015; 5:156‑62. [ Links ]
44. Westritschnig K, Sibanda E, Thomas W, Auer H, Aspöck H, Pittner G, et al. Analysis of the sensitization profile towards allergens in central Africa. Clin Exp Allergy 2003; 33:22‑7. [ Links ]
45. Weghofer M, Thomas WR, Kronqvist M, Mari A, Purohit A, Pauli G, et al Variability of IgE reactivity profiles among European mite allergic patients. Eur J Clin Invest 2008;38:959‑65. [ Links ]
46. Bronnert M, Mancini J, Birnbaum J, Agabriel C, Liabeuf V, Porri F, et al. Component‑resolved diagnosis with commercially available D. pteronyssinus Der p 1, Der p 2 and Der p 10: relevant markers for house dust mite allergy. Clin Exp Allergy 2012; 42:1406‑15. [ Links ]
47. Pereira dos Reis R, Pires AP, Tomaz E, Inácio F. Sensibilização a tropomiosina em doentes alérgicos a ácaros. Rev Por Imunoalergologia 2007; 15: 251‑58. [ Links ]
48. Gámez C, Sanchez‑Garcia S, Ibáñez MD, López R, Aguado E, López E, et al. Tropomyosin IgE‑positive results are a good predictor of shrimp allergy. Allergy 2011; 66:1375‑83. [ Links ]
49. Ayuso R, Sanchez‑Garcia S, Lin J, Fu Z, Ibáñez MD, Carrillo T, et al. Greater epitope recognition of shrimp allergens by children than by adults suggests that shrimp sensitization decreases with age. J Allergy Clin Immunol 2010; 125:1286‑93. [ Links ]
50. Satinover SM, Reefer AJ, Pomes A, Chapman MD, Platts‑Mills TA, Woodfolk JA. Specific IgE and IgG antibody‑binding patterns to recombinant cockroach allergens. J Allergy Clin Immunol 2005; 115:803‑9. [ Links ]
51. Jeong KY, Lee J, Lee IY, Ree HI, Hong CS, Yong TS. Allergenicity of recombinant Bla g 7, German cockroach tropomyosin. Allergy 2003; 58:1059‑63. [ Links ]
52. Saarne T, Kaiser L, Rasool O, Huecas S, van Hage‑Hamsten M, Gafvelin G. Cloning and characterisation of two IgE‑binding proteins, homologous to tropomyosin and alpha‑tubulin, from the mite Lepidoglyphus destructor. Int Arch Allergy Immunol 2003; 130:258‑65. [ Links ]
53. Muraro A, Werfel T, Hoffmann‑Sommergruber K, Roberts G, Beyer K, et al. EAACI food allergy and anaphylaxis guidelines: diagnosis and management of food allergy. Allergy 2014; 69:1008‑25. [ Links ]
54. Kattan JD, Sicherer SH. Optimizing the diagnosis of food allergy. Immunol Allergy Clin North Am 2015; 35:61‑76. [ Links ]
55. Munera M, Gómez L, Puerta L. El camarón como una fuente de alérgenos. Biomédica 2013; 33:306‑18. [ Links ]
56. Wong L, Huang CH, Lee BW. Shellfish and house dust mite allergies: Is the link tropomyosin? Allergy Asthma Immunol Res 2016; 8:101‑6. [ Links ]
57. Jirapongsananuruk O, Sripramong C, Pacharn P, Udomputunurak S, Chinratanapisit S, Piboonpocanun S, et al. Specific allergy to Penaeus monodon (seawater shrimp) or Macrobrachium rosenbergii (fresh water shrimp) in shrimp‑allergic children. Clin Exp Allergy 2008; 38:1038‑47. [ Links ]
58. Yang AC, Arruda LK, Santos AB, Barbosa MC, Chapman MD, Galvão CE, et al. Measurement of IgE antibodies to shrimp tropomyosin is superior to skin prick testing with commercial extract and measurement of IgE to shrimp for predicting clinically relevant allergic reactions after shrimp ingestion. J Allergy Clin Immunol 2010; 125:872‑8. [ Links ]
Joana Cosme
Serviço de Imunoalergologia, Hospital de Santa Maria, Centro Hospitalar de Lisboa Norte
Av. Prof. Egas Moniz,
1649‑035 Lisboa
Telefone: 217805000
Financiamento: Nenhum.
Declaração de conflitos de interesse: Nenhum.
Data de receção / Received in: 10/03/2016
Data de aceitação / Accepted for publication in: 08/06/2016


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}