










Serviços Personalizados
Journal

Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
 Similares em
SciELO
Similares em
SciELO Compartilhar

 Permalink
PermalinkNascer e Crescer
versão impressa ISSN 0872-0754versão On-line ISSN 2183-9417
Nascer e Crescer vol.27 no.3 Porto set. 2018
https://doi.org/10.25753/BirthGrowthMJ.v27.i3.13431
REVIEW ARTICLES | ARTIGOS DE REVISÃO
Child maltreatment and mental disorders – the role of epigenetics
Maus tratos na infância e doença mental – o papel da epigenética
Marta QueirósI; João CaseiroII
I Hospital de Magalhães Lemos. 4149-003 Porto, Portugal. martinhaqueiros@hotmail.com
II Department of Child and Adolescent Psychiatry, Centro Materno Infantil do Norte, Centro Hospitalar do Porto. 4099-001 Porto, Portugal. joao_caseiro@hotmail.com
ABSTRACT
Introduction: Child maltreatment is associated with high risk for various physical and mental disorders and is associated with over 30% of adult psychopathology. Child maltreatment also relates with poor clinical outcomes, such as chronic disease, increased hospitalization, comorbidity and reduced response to treatment. Early adverse experiences can cause epigenetic changes, altering gene expression without changing DNA sequence. Epigenetic alterations in genes implicated in stress response and neurodevelopment might explain to some extent the impact of child maltreatment in mental health.
Objectives: Review available literature concerning the impact of child maltreatment in human epigenome, focusing on mental health related outcomes.
Development: DNA methylation induced by child maltreatment changes specific genes, in peripheral and brain tissues, producing major consequences in stress regulation, neural plasticity, and neurodevelopment. Methylation of the glucocorticoid receptor gene is one of the most studied epigenetic alterations that have been related to childhood maltreatment and seems to be responsible for an increased vulnerability to develop psychopathology. Epigenetic changes may not be permanent, and there are some interventions that seem to reduce DNA methylation. Therefore, in the future, DNA methylation may be used, not only in the diagnosis and prediction of treatment response, but also for therapeutic innovation.
Conclusions: Epigenetic changes can potentially explain pathophysiological changes related to child maltreatment exposure, may serve as biomarkers in stress-related disorders and provide targets for the development of new therapeutic and preventive interventions for individuals that were exposed to child abuse.
Keywords: Child abuse; domestic violence; epigenesis; genetic; mental disorders
RESUMO
Introdução: A exposição a maus tratos na infância está associada a risco acrescido de doença física e mental, relacionando-se com mais de 30% da psicopatologia e com maus resultados clínicos, nomeadamente formas de doença crónicas, aumento da hospitalização, comorbilidades e baixa resposta terapêutica. Experiências adversas na infância podem causar alterações epigenéticas, alterando a expressão genética sem modificar a sequência do ADN. Alterações na expressão dos genes implicados na resposta ao stress e no neuro-desenvolvimento poderão explicar o impacto dos maus tratos infantis na saúde mental.
Objetivos: Revisão da literatura acerca do impacto dos maus tratos na infância no epigenoma humano, salientando as consequências para a saúde mental.
Desenvolvimento: A metilação de ADN induzida por trauma na infância provoca alterações em genes específicos, em tecidos periféricos e cerebrais, alterando significativamente a regulação da resposta ao stress, plasticidade cerebral e neuro-desenvolvimento. A metilação do gene do recetor de glucocorticoides é uma das alterações epigenéticas mais estudadas e parece ser responsável por uma maior vulnerabilidade para psicopatologia em crianças expostas a maus tratos. As alterações epigenéticas não são necessariamente permanentes e algumas intervenções parecem diminuir a metilação de ADN. Futuramente, a metilação de ADN poderá utilizar-se no diagnóstico, predição da resposta terapêutica e desenvolvimento de tratamentos inovadores.
Conclusões: Algumas alterações patofisiológicas observadas em indivíduos expostos a maus tratos na infância poderão ser explicadas por mecanismos epigenéticos. As alterações epigenéticas poderão servir como biomarcadores de doenças associadas ao stress e possibilitar o desenvolvimento de tratamentos inovadores para indivíduos expostos a maus tratos na infância.
Palavras-chave: abuso infantil; doença mental; epigenética; violência doméstica
INTRODUCTION
Child maltreatment (CM) is a public health problem that affects up to one in four children of all ages, races, cultures and socio-economic backgrounds worldwide.1-4 CM comprises psychological, physical and sexual abuse, as well as caregiver neglect.4-6
Early life adversities have been associated with various physical and mental disorders through the lifespan and with poor clinical outcomes.2,5,7-10 There is evidence indicating that over 30% of adult psychopathology is directly related to CM, namely depression, anxiety, post-traumatic stress disorder (PTSD), personality disorders, substance misuse and aggressive and suicidal behaviours.1,4-6,11 Individuals exposed to child abuse have a greater incidence of comorbidities and chronic disease, are more likely to be hospitalized, and show a reduced response to treatment.4,5,12 CM can be particularly harmful in early childhood since this is a sensitive period characterized by a rapid neuroendocrinological development, greater vulnerability to stress and environmental factors and increased brain plasticity.6,13 Children exposed to CM experience difficulties in emotion regulation and in maintaining healthy relationships, and may develop mood, anxiety, or behavioural disorders.9
Recent studies have examined epigenetic mechanisms that are responsible for altered expression of genes related to stress response, neurodevelopment and mood regulation. These epigenetic changes seem to mediate the association between CM and some psychiatric disorders.2,10
Epigenetic changes (EC) are the process by which environmental factors alter gene expression without changing the DNAs nucleotide sequence.4,10,11,14 Epigenetic alterations of DNA mainly occur in three different ways: histone modification, non-coding RNA associated gene silencing and DNA methylation.4,11
DNA methylation is the most studied mechanism in epigenetic research regarding the impact of CM in mental health.4,5 DNA methylation is a dynamic mechanism that can rapidly respond to environmental factors such as CM, resulting in physiological as well as in pathological processes, through the lifespan.4,12,15 DNA methylation occurs mainly at cytosine–guanine nucleotide (CpG) sites, which are overrepresented in promoter regions of genes.4,10,16 The promoter region of the gene is the sequence needed to turn the gene on and off, it is generally found near the transcription start site, and has binding sites for transcription factors and enzymes that produce RNA.9 These regions usually have low levels of DNA methylation leaving the genes accessible for transcription.4,17 When methylation occurs, methyl groups bind to CpG sites on the promoter region of a gene and the access of transcription factors to regulatory elements is blocked.4,5,11,14,16 Therefore, hypermethylation has generally been associated with gene silencing and hypomethylation with increased gene expression.16
The brain is one of the organs that is most prone to epigenetic plasticity, so it may be more susceptible to EC related to CM exposure, compared to other organs.4
OBJECTIVES
The aim of this article is to perform a literature review concerning the impact of CM in human epigenome and mental health related outcomes and to discuss possible implications for treatment.
DEVELOPMENT
We performed a non-systematic review using the PubMed database.
With the purpose of reviewing the various EC that have been associated to CM and mental health related outcomes, we exclusively analysed studies performed with human samples. We selected fourteen articles that focused on mental health related outcomes and eight that focused on physiological and neurobiological outcomes.
Regarding the possible implications for treatment, and given the paucity of data, we also reviewed studies with animal samples and those that did not relate specifically to CM, selecting ten articles.
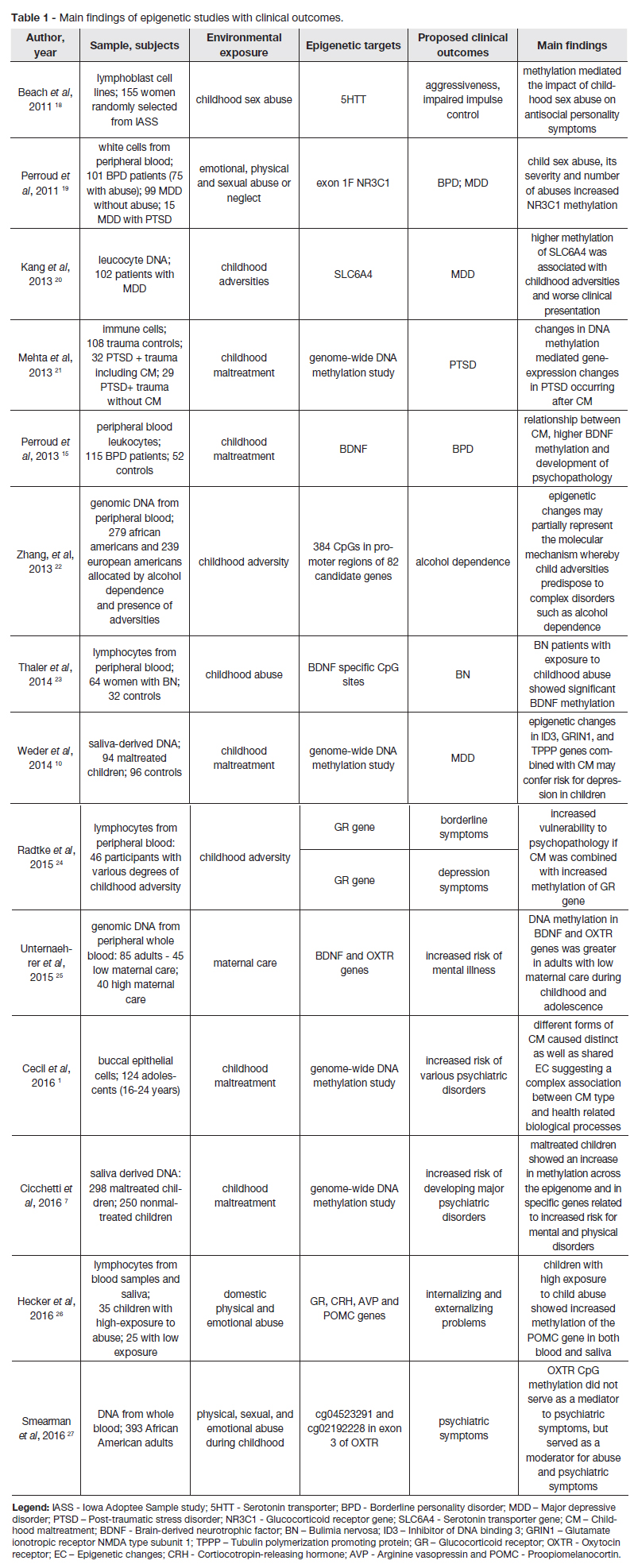
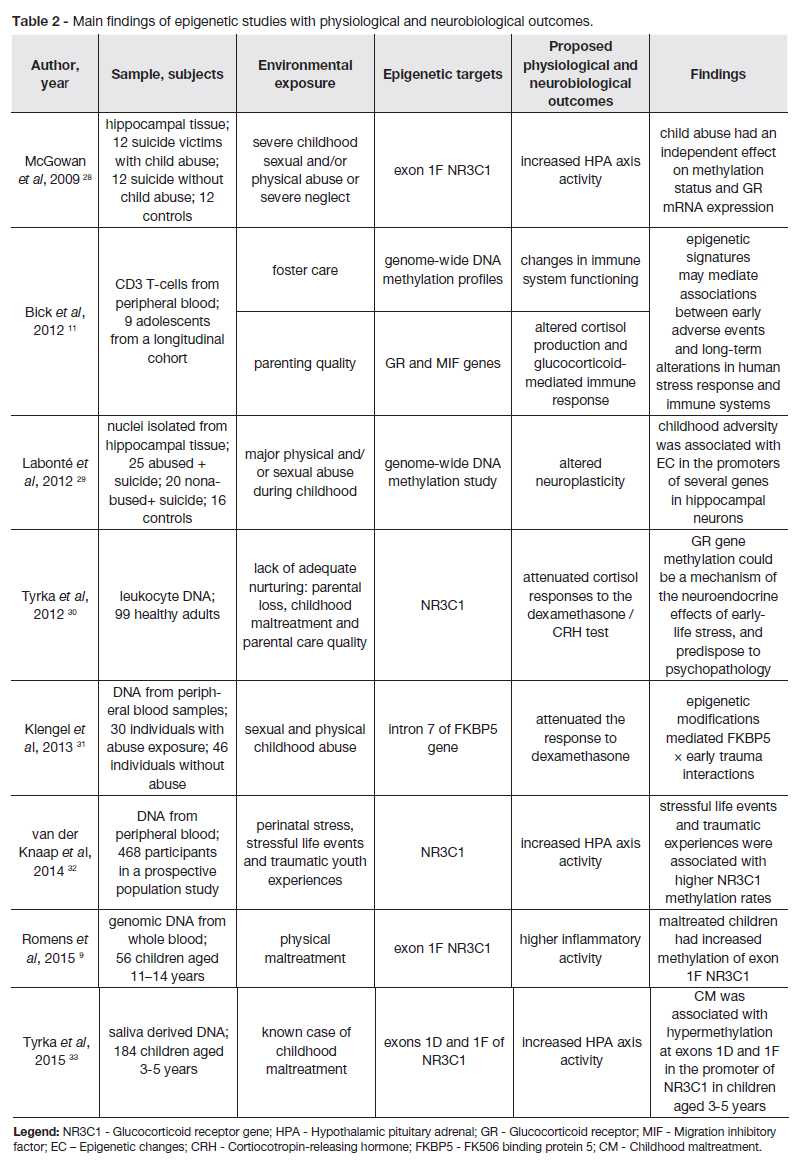
Table 1 and table 2 summarize the main findings of epigenetic studies in individuals exposed to CM. There is a great methodological variation between studies in the definition of maltreatment, measures of exposure, type of samples used and population characteristics. We chose to separate the data according to the type of outcomes. In table 1 we present studies with clinical outcomes and in table 2 studies with physiological and neurobiological outcomes. The data is organized by date of publishing.
We focus our review on those studies that examined the relationship between CM and mental health. We present the main findings according to the affected physiological pathway.
Hypothalamic pituitary adrenal axis
The hypothalamic pituitary adrenal (HPA) axis has a major role in the regulation of stress response and other biological functions such as growth, reproduction, metabolism and circadian rhythm, and is particularly affected by early life stress.11,19,24 Recent research has highlighted the effect of CM in the expression of genes related to the HPA axis, regulated through epigenetic mechanisms.
HPA axis activity is regulated by negative feedback.11,12 After an acute stress response, glucocorticoids bind to glucocorticoid receptor (GR) in components of the HPA axis, but also in the hippocampus and other brain structures, inhibiting further activation and restoring homeostasis.12 The disruption of this regulatory mechanism is known to be associated with various psychiatric disorders.12,24
The most extensively investigated and reviewed gene in epigenetic studies of childhood maltreatment is the NR3C1 gene which encodes the GR.
McGowan et al performed the first study translating findings from animal studies to humans. They examined hippocampal samples obtained from suicide victims, either exposed or not exposed to CM, and control subjects who died suddenly and had no history of CM. The authors found hypermethylation of the exon 1F of NR3C1 and decreased hippocampal GR expression in suicide victims with a history of CM compared with controls and with suicide victims without history of CM. This study suggests that early life events can induce EC of relevant genomic regions responsible for HPA axis regulation, which could contribute to an increased risk for psychopathology.28
Perroud et al, compared a sample of patients with borderline personality disorder (BPD) with various degrees of CM exposure, against samples of patients with major depressive disorder (MDD) and patients with MDD and co-morbid PTSD. In the BPD sample there was a significant association between childhood sexual abuse and physical neglect and hypermethylation of NR3C1. Other types of abuse were not significantly associated with methylation status. Repeated abuses, number of types of abuse and neglect and sexual abuse with penetration correlated with a higher methylation percentage in BDP patients. When considering the whole sample, all types of CM were associated with NR3C1 methylation. The authors hypothesise that EC in the NR3C1 gene may link CM to the development of BPD and that NR3C1 methylation may be a marker of CM severity.19 These results were obtained using a peripheral blood sample, indicating that what was observed in the brain by McGowan et al was true for the peripheral blood.19,28
Bick et al examined peripheral blood samples from nine adolescents in order to study DNA methylation patterns of GR and macrophage migration inhibitory factor (MIF) genes, according to parenting quality assessed five to ten years before. MIF controls the glucocorticoid-mediated suppression of inflammation and cytokine release and is associated with GR expression and immune system activity. Bad parenting, measured by the Parent–Adolescent Relationship Questionnaire, was associated with hypermethylation of both GR and MIF genes, suggesting that inferior caregiving quality may induce long term EC.11
Several other studies have supported the role of different types of CM in inducing hypermethylation of the GR gene, low GR expression, disrupted negative feedback inhibition of the HPA axis with altered basal cortisol secretion and HPA axis reactivity.9,24,30,32,33 A study by Radtke et al demonstrated an association between CM induced EC and the development of psychopathologic symptoms.24
Although epigenetic changes may be tissue specific, they can occasionally be observed in DNA from peripheral as well as brain samples.16,26 By using different DNA sources, studies have demonstrated that DNA methylation of GR gene occurs at overlapping CpG sites in brain tissue, peripheral blood and saliva samples.16,19,28,33 The consistent results in both brain and peripheral samples make the GR gene a promising candidate for use as a biomarker.
FKBP5, a functional regulator of the GR complex, has also been studied as a target for CM induced epigenetic changes. Klengel et al found that a FKBP5 polymorphism when associated with CM induced DNA demethylation of glucocorticoid response elements in the FKBP5 locus, increased FKBP5 responsiveness and GR resistance. This would lead to altered stress hormone regulation, changes in neuronal circuits and in other GR responsive systems, increasing the risk of developing stress-related psychopathology.31
Proopiomelanocortin (POMC) is a precursor of the adrenocorticotropic hormone (ACTH), one of the main hormones involved in the regulation of the HPA axis. Hecker et al, in a study performed with Tanzanian children, found a strong association between high-exposure to child abuse and hypermethylation of the POMC gene in both blood and saliva samples. There was a high concordance between childrens self-report and methylation status, suggesting that POMC methylation may serve as a biomarker of child abuse in this population.26
Serotonin
Serotonin is a neurotransmitter implicated in several physiological processes, such as mood regulation, emotional processing, memory, social interaction and HPA axis regulation.34 Altered serotonin neurotransmission is thought to be a risk factor for psychopathology and occurs more frequently when there is an exposure to environmental stressors.35 For instance, disfunction of the serotonergic system is known to play a role in the increased risk of depression in individuals exposed to CM.18,20 The serotonin transporter gene (SLC6A4) regulates serotonergic neurotransmission and seems to be implicated in the aforementioned gene-environment association.Hypermethylation of SLC6A4 promoter is reported to be associated with decreased brain serotonin synthesis and with the consequent development of psychopathology.20,34
A study by Beach et al concluded that a history of childhood sexual abuse was associated with methylation of the promoter region of SLC6A4, in a sample of 155 women. Also, SLC6A4 mediated the link between sexual abuse and antisocial personality disorder symptoms, namely aggressiveness and impaired impulse control.18
In a sample of depressed patients, Kang et al found that higher methylation of SLC6A4 was associated with childhood adversities and worse clinical presentation. The authors proposed that, in the future, SLC6A4 methylation status could be used as a biomarker for childhood adversities and for certain presentations of depression.20
Brain-derived neurotrophic factor
Brain-derived neurotrophic factor (BDNF) is a neurotrophin implicated in cellular proliferation and survival, synaptic activity and neural plasticity.23 BDNF seems to be associated with the interaction between CM and neurobehavioral outcomes.13
In a sample of 115 patients with BPD and 52 controls, Perroud et al found a relationship between CM and higher DNA methylation of BDNF. This study suggests that CM induces EC in the BDNF gene, which may contribute to the development of psychopathology.15
Thaler et al investigated the link between bulimia nervosa (BN), BDNF methylation and child abuse. They compared 64 women with BN diagnosis and 32 controls and found an increased methylation of specific CpG sites in BN patients, particularly in those exposed to CM.23
Unternaehrer et al analysed DNA methylation in a sample of 85 adults and screened them for maternal care quality using the Parental Bonding Instrument. They found a greater BDNFTS methylation in the low maternal care group compared to the high maternal care group.25
Oxytocin
Oxytocin is a neurohormone that increases the salience of social cues and has been associated with socialization and development of affective bonds. Oxytocin has a role in the pathophysiology of some psychiatric disorders, particularly those characterized by impairments in social functioning. CM seems to influence the oxytocin system by decreasing oxytocin levels and the expression of oxytocin receptors (OXTR). One of the proposed mechanisms for this association is induction of EC in the OXTR gene.27
In a sample of 85 adults screened for maternal care quality, Unternaehrer et al found a greater OXTRTS2 methylation in the low maternal care group compared to the high maternal care group. The authors suggest that EC in the OXTR gene induced by low maternal care might contribute to changes in stress-reactivity and consequently increase the risk for psychopathology.25
A study by Smearman et al, found that history of abuse during childhood predicted higher methylation of CpG sites of OXTR in a sample of 393 African American adults. OXTR methylation did not mediate the association between abuse and psychiatric symptoms, but interacted with abuse to predict psychiatric outcomes such as depression and anxiety.27
Genome-wide studies
Early adverse events have been found to affect DNA methylation profiles of candidate genes and across the entire genome.11 Genome-wide studies provide an overview of differential DNA methylation profiles across the genome and help to generate hypothesis for candidate-gene studies. Genome-wide approaches are particularly useful when studying complex disorders such as mental disorders.5,11
Bick et al used a genome-wide approach to analyse changes in methylation patterns according to foster care exposure, in a sample of nine adolescents. Adolescents with history of foster care showed hypermethylation in 72 genes, and hypomethylation in 101 genes. The most relevant genes that were differentially methylated were related with immune system functioning, suggesting that exposure to foster care may alter immune response.11
In a sample of 41 suicide victims with a history of childhood abuse, Labonté et al found 248 hypermethylated and 114 hypomethylated CpG sites, when compared with non-abused suicide victims. In this study, the most significantly differentially methylated genes were involved in neuronal plasticity. The authors suggest that these findings may be relevant to understand the mechanisms underlying suicide risk in individuals exposed to CM.29
In a sample of PTSD patients, Mehta et al found a significantly higher number of EC in the childhood trauma exposed group, suggesting a greater role of epigenetic mechanisms in the pathophysiology of this type of PTSD. They propose that considering the exposure to CM in patients with PTSD, it may be useful to analyse the potential application of EC as biomarkers for PTSD and as therapeutic targets.21
Other genome-wide studies have consistently found an increased differential methylation across the genome in individuals exposed to CM and support the possible role of CM induced EC in the pathophysiology of stress-related psychiatric disorders.1,7,10
Cecil et al found maltreatment-specific methylation patterns in individuals exposed to various forms of CM. These findings indicate that different types of CM may induce EC in genes related to distinct biological pathways and therefore explain different clinical presentations.1
Implications for treatment
Although DNA methylation is thought to be the most stable form of epigenetic modification, there is evidence that gene silencing induced by promoter methylation is reversible.2,36 Consequently, by reverting DNA methylation, it may be possible to prevent or treat certain psychiatric symptoms in individuals that were exposed to CM.37
DNA methylation can be reversed using several drugs, namely histone deacetylase inhibitors, such as trichostatin A, and DNA methylation inhibitors, such as zebularine and 5-aza-2-deoxycytidine.2,37 We will focus on drugs that are commonly used in psychiatry.
In a study using rodent brain samples, Dong et al found that clozapine and sulpiride induced demethylation in cortical and striatal regions with previously hypermethylated GABAergic gene promoters. This effect was potentiated by associating valproate. The authors propose that, in schizophrenic and bipolar patients, some of the therapeutic effect of the co-administration of atypical antipsychotics with valproate may be due to DNA demethylation.38
A later study by the same investigators, found that valproate alone was effective in inducing promoter demethylation of two cortical GABAergic genes in extracts of mouse frontal cortex.36
In a naturalistic sample of depressed patients treated with several antidepressants, Tadic et al found a lower methylation of the promoter region of the BDNF gene in the patients that clinically improved, regardless of the class of antidepressant used, suggesting that antidepressants may promote DNA demethylation.39
The presented findings suggest that understanding how drugs induce DNA demethylation may lead to the development of innovative treatments for psychiatric disorders such as schizophrenia, bipolar disorder and depression.
Psychotherapy also seems to influence DNA methylation.15,40,41 Even though two of the following studies, by Yehuda et al and by Roberts et al, didn´t use samples with a history of CM, we included them in this review attending to their relevance and focus on EC in response to psychotherapy.
Perroud et al evaluated 115 subjects with BPD that underwent a four week course of intensive dialectical behaviour therapy (DBT) and 52 controls. BDP patients that were exposed to CM had higher BDNF methylation at the beginning of the study. BPD patients that responded to DBT treatment showed a decrease in BDNF methylation status after the intervention that related with improvement in depression scores, hopelessness scores and impulsivity. The authors suggest that BDNF methylation levels may serve as a biomarker of psychopathology in patients exposed to CM and may be used to predict the response to psychotherapeutic interventions.15
A study by Yehuda et al examined the methylation patterns of DNA extracted from lymphocytes of combat veterans with PTSD that underwent a prolonged exposure therapy. Responders had a higher pre-treatment methylation of the exon 1F NR3C1 promoter compared to non-responders. Responders presented a decrease in FKBP5 promoter methylation associated with recovery, whereas non-responders showed an increase in FKBP5 promoter methylation. The authors conclude that exon 1F NR3C1 methylation may be used as a prognosis biomarker, while FKBP5 gene methylation may relate to symptom severity, and that successful psychotherapy may constitute a form of environmental regulation that alters the epigenome.41
Roberts et al analysed a large sample of children with anxiety disorders that were offered cognitive behavioural therapy (CBT). They found an association between changes in FKBP5 gene methylation and CBT treatment response.40
The aforementioned evidence indicates that psychotherapy may be an effective way of reducing demethylation in stress related disorders with a correspondent improvement in clinical outcome.
Studies in rodent samples suggest that physical exercise may modulate epigenetic changes induced by repeated stress by increasing the global DNA methylation profile in the rat hypothalamus.42
Although most of the interventions mentioned above have not been investigated in human samples with CM exposure, current knowledge may useful for the future development of DNA demethylation methods to prevent or treat trauma related psychopathology.
CONCLUSIONS
Early life adversities have been associated with an increased risk of several mental disorders. Recent research has tried to clarify the mechanisms that underlie this association. Epigenetics may help to explain some interactions between early environmental exposures and changes in gene expression.
Epigenetic studies performed in individuals exposed to CM have proposed various epigenetic pathways through which CM may induce pathophysiological changes. This knowledge will hopefully help to clarify disease mechanisms of some complex disorders, namely stress-related psychopathology.
EC are promising candidates for use as biomarkers in stress-related disorders, contributing to an improvement in clinical diagnosis and to the prediction of treatment response.
Epigenetic research related to CM may contribute to the validation of current treatments and lead to the development of new therapeutic and preventive interventions that, by changing methylation levels of specific genes, could improve mental health outcomes in individuals exposed to CM.
This review has several limitations. Generalization of these results is limited by great methodological variations between studies in the definition of maltreatment, measures of exposure, type of samples used and population characteristics. We found no studies concerning histone modification and non-coding RNA associated gene silencing, as such, we can only consider the mechanism of DNA methylation when discussing our results. Moreover, most studies use retrospective self-report data which can lead to unreliable estimates of CM exposure.
Future research should aim to further elucidate the association between different forms of maltreatment, EC across the developmental stages and the impact on adult psychopathology. There is a need for prospective studies with larger samples of maltreatment-exposed children to confirm former findings and to clarify the complex relationship between CM and adverse mental health outcomes.
REFERENCES
1. Cecil CA, Smith RG, Walton E, Mill J, McCrory EJ, Viding E. Epigenetic signatures of childhood abuse and neglect: Implications for psychiatric vulnerability. J Psychiatr Res. 2016; 83:184-94. [ Links ]
2. Tyrka AR, Ridout KK, Parade SH. Childhood adversity and epigenetic regulation of glucocorticoid signaling genes: Associations in children and adults. Dev Psychopathol. 2016; 28:1319-31. [ Links ]
3. Pollak SD. Multilevel developmental approaches to understanding the effects of child maltreatment: Recent advances and future challenges. Dev Psychopathol. 2015; 27:1387-97. [ Links ]
4. Lutz PE, Turecki G. DNA methylation and childhood maltreatment: from animal models to human studies. Neuroscience. 2014; 264:142-56. [ Links ]
5. Lutz PE, Almeida D, Fiori LM, Turecki G. Childhood maltreatment and stress-related psychopathology: the epigenetic memory hypothesis. Curr Pharm Des. 2015; 21:1413-7. [ Links ]
6. Schury K, Kolassa IT. Biological memory of childhood maltreatment: current knowledge and recommendations for future research. Ann N Y Acad Sci. 2012; 1262:93-100. [ Links ]
7. Cicchetti D, Hetzel S, Rogosch FA, Handley ED, Toth SL. An investigation of child maltreatment and epigenetic mechanisms of mental and physical health risk. Dev Psychopathol. 2016; 28:1305-17. [ Links ]
8. Nugent NR, Goldberg A, Uddin M. Topical Review: The Emerging Field of Epigenetics: Informing Models of Pediatric Trauma and Physical Health. J Pediatr Psychol. 2016; 41:55-64. [ Links ]
9. Romens SE, McDonald J, Svaren J, Pollak SD. Associations between early life stress and gene methylation in children. Child Dev. 2015; 86:303-9. [ Links ]
10. Weder N, Zhang H, Jensen K, Yang BZ, Simen A, Jackowski A, et al. Child abuse, depression, and methylation in genes involved with stress, neural plasticity, and brain circuitry. J Am Acad Child Adolesc Psychiatry. 2014; 53:417-24.e5. [ Links ]
11. Bick J, Naumova O, Hunter S, Barbot B, Lee M, Luthar SS, et al. Childhood adversity and DNA methylation of genes involved in the hypothalamus-pituitary-adrenal axis and immune system: whole-genome and candidate-gene associations. Dev Psychopathol. 2012; 24:1417-25. [ Links ]
12. Anacker C, O’Donnell KJ, Meaney MJ. Early life adversity and the epigenetic programming of hypothalamic-pituitary-adrenal function. Dialogues Clin Neurosci. 2014; 16:321-33.
13. Roth TL, Sweatt JD. Epigenetic marking of the BDNF gene by early-life adverse experiences. Horm Behav. 2011; 59:315-20. [ Links ]
14. De Bellis MD, Zisk A. The biological effects of childhood trauma. Child Adolesc Psychiatr Clin N Am. 2014; 23:185-222. [ Links ]
15. Perroud N, Salzmann A, Prada P, Nicastro R, Hoeppli ME, Furrer S, et al. Response to psychotherapy in borderline personality disorder and methylation status of the BDNF gene. Transl Psychiatry. 2013;3:e207. [ Links ]
16. Menke A, Binder EB. Epigenetic alterations in depression and antidepressant treatment. Dialogues Clin Neurosci. 2014; 16:395-404. [ Links ]
17. Smart C, Strathdee G, Watson S, Murgatroyd C, McAllister-Williams RH. Early life trauma, depression and the glucocorticoid receptor gene-an epigenetic perspective. Psychol Med. 2015; 45:3393-410. [ Links ]
18. Beach SR, Brody GH, Todorov AA, Gunter TD, Philibert RA. Methylation at 5HTT mediates the impact of child sex abuse on women’s antisocial behavior: an examination of the Iowa adoptee sample. Psychosom Med. 2011; 73:83-7.
19. Perroud N, Paoloni-Giacobino A, Prada P, Olie E, Salzmann A, Nicastro R, et al. Increased methylation of glucocorticoid receptor gene (NR3C1) in adults with a history of childhood maltreatment: a link with the severity and type of trauma. Transl Psychiatry. 2011; 1:e59. [ Links ]
20. Kang HJ, Kim JM, Stewart R, Kim SY, Bae KY, Kim SW, et al. Association of SLC6A4 methylation with early adversity, characteristics and outcomes in depression. Prog Neuropsychopharmacol Biol Psychiatry. 2013; 44:23-8. [ Links ]
21. Mehta D, Klengel T, Conneely KN, Smith AK, Altmann A, Pace TW, et al. Childhood maltreatment is associated with distinct genomic and epigenetic profiles in posttraumatic stress disorder. Proc Natl Acad Sci U S A. 2013; 110:8302-7. [ Links ]
22. Zhang H, Wang F, Kranzler HR, Zhao H, Gelernter J. Profiling of childhood adversity-associated DNA methylation changes in alcoholic patients and healthy controls. PLoS One. 2013; 8:e65648. [ Links ]
23. Thaler L, Gauvin L, Joober R, Groleau P, de Guzman R, Ambalavanan A, et al. Methylation of BDNF in women with bulimic eating syndromes: associations with childhood abuse and borderline personality disorder. Prog Neuropsychopharmacol Biol Psychiatry. 2014; 54:43-9. [ Links ]
24. Radtke KM, Schauer M, Gunter HM, Ruf-Leuschner M, Sill J, Meyer A, et al. Epigenetic modifications of the glucocorticoid receptor gene are associated with the vulnerability to psychopathology in childhood maltreatment. Transl Psychiatry. 2015; 5:e571. [ Links ]
25. Unternaehrer E, Meyer AH, Burkhardt SC, Dempster E, Staehli S, Theill N, et al. Childhood maternal care is associated with DNA methylation of the genes for brain-derived neurotrophic factor (BDNF) and oxytocin receptor (OXTR) in peripheral blood cells in adult men and women.Stress. 2015; 18:451-61. [ Links ]
26. Hecker T, Radtke KM, Hermenau K, Papassotiropoulos A, Elbert T. Associations among child abuse, mental health, and epigenetic modifications in the proopiomelanocortin gene (POMC): A study with children in Tanzania. Dev Psychopathol. 2016; 28:1401-12. [ Links ]
27. Smearman EL, Almli LM, Conneely KN, Brody GH, Sales JM, Bradley B, et al. Oxytocin Receptor Genetic and Epigenetic Variations: Association With Child Abuse and Adult Psychiatric Symptoms. Child Dev. 2016; 87:122-34. [ Links ]
28. McGowan PO, Sasaki A, D’Alessio AC, Dymov S, Labonte B, Szyf M, et al. Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nat Neurosci. 2009; 12:342-8.
29. Labonte B, Suderman M, Maussion G, Navaro L, Yerko V, Mahar I, et al. Genome-wide epigenetic regulation by early-life trauma. Arch Gen Psychiatry. 2012; 69:722-31. [ Links ]
30. Tyrka AR, Price LH, Marsit C, Walters OC, Carpenter LL. Childhood adversity and epigenetic modulation of the leukocyte glucocorticoid receptor: preliminary findings in healthy adults. PLoS One. 2012; 7:e30148. [ Links ]
31. Klengel T, Mehta D, Anacker C, Rex-Haffner M, Pruessner JC, Pariante CM, et al. Allele-specific FKBP5 DNA demethylation mediates gene-childhood trauma interactions. Nat Neurosci. 2013; 16:33-41. [ Links ]
32. van der Knaap LJ, Riese H, Hudziak JJ, Verbiest MM, Verhulst FC, Oldehinkel AJ, et al. Glucocorticoid receptor gene (NR3C1) methylation following stressful events between birth and adolescence. The TRAILS study. Transl Psychiatry. 2014; 4:e381. [ Links ]
33. Tyrka AR, Parade SH, Eslinger NM, Marsit CJ, Lesseur C, Armstrong DA, et al. Methylation of exons 1D, 1F, and 1H of the glucocorticoid receptor gene promoter and exposure to adversity in preschool-aged children. Dev Psychopathol. 2015; 27:577-85. [ Links ]
34. Provenzi L, Giorda R, Beri S, Montirosso R. SLC6A4 methylation as an epigenetic marker of life adversity exposures in humans: A systematic review of literature. Neurosci Biobehav Rev. 2016; 71:7-20. [ Links ]
35. Booij L, Tremblay RE, Szyf M, Benkelfat C. Genetic and early environmental influences on the serotonin system: consequences for brain development and risk for psychopathology. J Psychiatry Neurosci. 2015; 40:5-18. [ Links ]
36. Dong E, Chen Y, Gavin DP, Grayson DR, Guidotti A. Valproate induces DNA demethylation in nuclear extracts from adult mouse brain. Epigenetics. 2010; 5:730-5. [ Links ]
37. Szyf M. The genome-and system-wide response of DNA methylation to early life adversity and its implication on mental health. Can J Psychiatry. 2013; 58:697-704. [ Links ]
38. Dong E, Nelson M, Grayson DR, Costa E, Guidotti A. Clozapine and sulpiride but not haloperidol or olanzapine activate brain DNA demethylation. Proc Natl Acad Sci USA. 2008; 105:13614-9. [ Links ]
39. Tadic A, Muller-Engling L, Schlicht KF, Kotsiari A, Dreimuller N, Kleimann A, et al. Methylation of the promoter of brain-derived neurotrophic factor exon IV and antidepressant response in major depression. Mol Psychiatry. 2014; 19:281-3. [ Links ]
40. Roberts S, Keers R, Lester KJ, Coleman JR, Breen G, Arendt K, et al. HPA axis related genes and response to psychological therapies: genetics and epigenetics. Depress Anxiety. 2015; 32:861-70. [ Links ]
41. Yehuda R, Daskalakis NP, Desarnaud F, Makotkine I, Lehrner AL, Koch E, et al. Epigenetic Biomarkers as Predictors and Correlates of Symptom Improvement Following Psychotherapy in Combat Veterans with PTSD. Front Psychiatry. 2013; 4:118. [ Links ]
42. Kashimoto RK, Toffoli LV, Manfredo MH, Volpini VL, Martins-Pinge MC, Pelosi GG, et al. Physical exercise affects the epigenetic programming of rat brain and modulates the adaptive response evoked by repeated restraint stress. Behav Brain Res. 2016; 296:286-9. [ Links ]
Marta Queirós
Hospital de Magalhães Lemos
R. Prof. Álvaro Rodrigues
4149-003 Porto
Email: martinhaqueiros@hotmail.com
Received for publication: 31.10.2017
Accepted in revised form: 05-07-2018


{kind=link}
{kind=link}