










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkMotricidade
versão impressa ISSN 1646-107X
Motri. vol.13 no.1 Ribeira de Pena mar. 2017
ARTIGO DE REVISÃO
Imunometabolismo e Exercício Físico: Uma nova fronteira do conhecimento
Immunometabolism and Exercise: New avenues
Barbara de Moura Antunes1,*; Fabrício Eduardo Rossi1; Daniela Sayuri Inoue1; José Cesar Rosa Neto2; Fábio Santos Lira1
1 Grupo de ImunoMetabolismo e Exercício, Departamento de Educação física, Universidade Estadual Paulista, UNESP, Faculdade de Ciências e Tecnologia, Campus de Presidente Prudente, São Paulo, Brasil
2 Laboratório de ImunoMetabolismo, Departamento de Biologia Celular e do Desenvolvimento, Instituto de Ciências Biomédicas, Universidade de São Paulo, USP, São Paulo, SP, Brasil
RESUMO
Sistema imunológico e metabolismo celular interagem constantemente por meio de proteínas imuno-moduladoras, denominadas citocinas, a fim de manter a dinâmica e a comunicação entre os sistemas sendo, na atualidade, amplamente difundido nas diversas áreas do conhecimento, principalmente no campo da saúde, como Imunometabolismo. As principais doenças do século 21 caracterizam-se por apresentar processos inflamatórios bem definidos os quais favorecem o agravamento da doença e a instalação de co-morbidades associadas à alteração metabólica. Em linhas gerais, a resposta inflamatória está associada a dieta inadequada e sedentarismo, alterando a dinâmica metabólica entre citocinas, ácidos graxos e endotoxina e ativando, assim, fatores de transcrição gênica, como o NF-kB. Por outro lado, a prática regular de exercício físico é amplamente recomendada como potente ferramenta na prevenção e tratamento de distúrbios metabólicos em virtude de seu poder anti-inflamatório e anti-aterogênico por meio da produção de miocinas advindas do músculo-esquelético e com atuação anti-inflamatória, como a IL-6, provenientes dos estímulos gerados pela contração muscular, e ativação de proteínas e fatores de transcrição gênica, como o PPAR. Desta forma, o objetivo da presente revisão é contextualizar e difundir os principais conceitos de uma emergente área do conhecimento, o Imunometabolismo, caracterizando sua função e atuação nos campos da doença e da saúde por meio da prática do exercício físico.
Palavras-chave: Inflamação, Metabolismo, Imunometabolismo, Citocinas, Exercício Físico
ABSTRACT
Immune system and cell metabolism have been interacted constantly, by synthesis and release of immunomodulatory proteins, recognized as cytokines, in order to maintain the body homeostasis. The crosstalk between systems has been widely spread in several areas of knowledge, mainly in the health field, as Immunometabolism. Diseases of the 21st century, which affect world populations, are characterized by inflammatory processes favoring the worsening of the illness itself and the installation of other co-morbidities. In general, the inflammatory response is associated with poor diet and physical inactivity, which modify the metabolic dynamics among cytokines, fatty acids and endotoxin culminating in reaction cascades, activating gene transcription factors, such as NF-kB. Furthermore, regular physical exercise is widely recommended as a powerful tool at the prevention and treatment of several metabolic diseases in virtue of anti-inflammatory and anti-atherogenic capacity, through anti-inflammatory myokine production, such as IL-6, from the stimuli generated by muscle contraction and proteins and gene transcription factors activation, as PPAR, which increase the anti-inflammatory response. Thus, the purpose of this review is to contextualize and disseminate knowledge about an emerging area, the Immunometabolism, featuring its function and performance in the illness and health fields by physical exercise.
Keywords: Inflammation, Metabolism, Immunometabolism, Cytokines, Physical activity
INTRODUÇÃO
O comportamento sedentário e os maus hábitos alimentares estão associados com o desenvolvimento da inflamação crônica de baixo grau, a qual é um processo metabólico característico de diversas doenças crônicas degenerativas e/ou não transmissíveis (DCNTs), como obesidade (Ghigliotti et al., 2014), resistência insulínica e diabetes mellitus tipo 2 (Chen, Chen, Wang, & Liang, 2015), dislipidemias (Antunes et al., 2015), câncer (Seelaender, Batista, Lira, Silverio, & Rossi-Fanelli, 2012), aterosclerose (Husain, Hernandez, Ansai, & Ferder, 2015), entre outras, que modifica drasticamente o funcionamento das células imunológicas bem como o metabolismo energético do organismo.
Clinicamente, o quadro inflamatório relacionado com as DCNTs está associado com elevada síntese e liberação de marcadores pró-inflamatórios, como proteína C-reativa, fator de necrose tumoral-alfa (TNF-α) e interleucinas (IL-6, IL-1β) (Lira, Neto, & Seelaender, 2014), simultaneamente à diminuição nas concentrações circulantes de marcadores anti-inflamatórios como adiponectina, interleucina 10 (IL-10) e recetor antagonista da interleucinas (IL-1ra) (Ohashi, Shibata, Murohara, & Ouchi, 2014).
O comportamento sedentário é apontado como fator determinante para a instalação e desenvolvimento das doenças supracitadas e, frente ao exposto, há um consenso na área da saúde de que a prática regular de exercício físico possui efeitos protetivos à saúde por sua capacidade indutiva de efeitos anti-inflamatórios e anti-aterogênicos por prevenir ou diminuir o risco à formação de ateromas nas paredes arteriais (Nimmo, 2013; Petersen, 2005). Os benefícios do exercício físico estão diretamente associados com a produção de miocinas (Pedersen, 2011; Pedersen, 2012), advindas da contração muscular, que atuam de maneira autócrina e parácrina (Atuação autócrina=Sinalização celular na qual um fator liberado atua a função da própria célula produtora; Atuação parácrina= Sinalização celular na qual um fator liberado pela célula secretora atua em outras estruturas do mesmo ambiente local), bem como potencializam a síntese e liberação de outros marcadores anti-inflamatórios como a IL-10 e IL-1ra (Steensberg, Fischer, Keller, Møller, & Pedersen, 2003).
No âmbito da saúde ou da doença é evidente a constante e direta interação entre células imunológicas, marcadores inflamatórios e metabolismo, originando uma emergente área de investigação denominada de Imunometabolismo, e frente a este panorama o objetivo desta revisão é contextualizar, mediante a busca e utilização principais estudos indexados em bases de dados nacionais e internacionais, e difundir os conhecimentos sobre a área de Imunometabolismo caracterizando sua função e atuação no campo da doença e saúde por meio da prática do exercício físico.
Imunometabolismo: da origem a atualidade
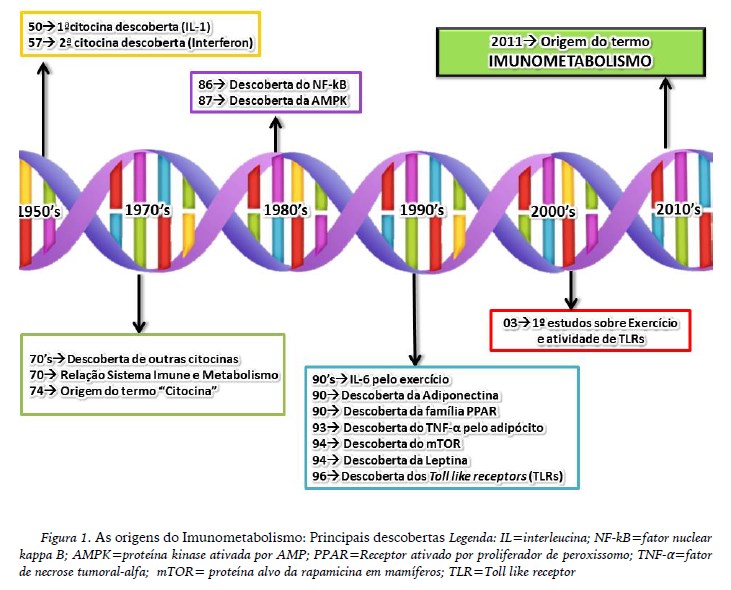
A saúde do organismo é constantemente assegurada pela fundamental interação entre sistema imunológico e processos metabólicos, onde as células imunológicas compõem uma importante linha de defesa contra agentes estressores e, frente à resposta imunológica, significantes alterações no metabolismo celular ocorrem para o reestabelecimento da homeostase corporal. A partir destas relações, deu-se o termo Imunometabolismo, que pode ser definido como um crosstalk constante entre o sistema imunológico e a atuação do metabolismo celular.
Em 2011, em um artigo de revisão, teve-se a primeira menção do termo Imunometabolismo por Mathis e Shoelson, evidenciando a necessidade de investigações acerca da interação entre imunologia e metabolismo no contexto, neste caso, da obesidade, tendo em vista que esta doença afeta a resposta imunológica em virtude do desenvolvimento e instalação da inflamação crônica de baixo grau provocando significantes alterações metabólicas. Conceitualmente, a inflamação crônica de baixo grau corresponde ao aumento de duas a quatro vezes das concentrações séricas basais de mediadores inflamatórios que atuam como preditores de mortalidade (Candore, Caruso, & Colonna-Romano, 2010).
Atualmente, diversos estudos caracterizam os padrões morfofuncionais do tecido adiposo e elucidam a presença de inúmeras células imunológicas residentes (Mraz & Haluzik, 2014; Schipper, Prakken, Kalkhoven, & Boes, 2012), como macrófagos e linfócitos, que sofrem alterações metabólicas e funcionais em resposta à obesidade (Lee & Lee 2014; Lumeng, Bodzin, & Saltiel, 2007). Por outro lado, outras doenças advindas do comportamento sedentário, como dislipidemia, diabetes mellitus tipo 2, hipertensão arterial, câncer, entre outras, também apresentam impactantes modificações metabólicas, reconhecidas atualmente como doençoma (do inglês diseasome) da inatividade física (Pedersen, 2009), e um quadro inflamatório crônico de baixo grau com elevada atividade das células imunes (McNelis & Olefscky, 2014) e produção de citocinas inflamatórias como TNF-α, IL-6, IL-1β e proteínas quimioatraentes de células imunológicas, como a proteína quimiotáctica de monócitos-1 (MCP-1) (Olefscky & Glass, 2010).
Um dos principais elos de comunicação entre as células imunológicas e o metabolismo são as citocinas, as quais podem ser caracterizadas como proteínas sinalizadoras e/ou imuno-moduladoras com atuação autócrina, parácrina e endócrina, sintetizadas principalmente por linfócitos, monócitos e macrófagos, e que são capazes de receber e enviar estímulos para os mais diversos tecidos e órgãos (Pillon, Bilan, Fink, & Klip, 2013).
As citocinas, na década de 70, eram intituladas como linfocinas, pois se acreditava que eram sintetizadas exclusivamente por linfócitos, posteriormente, foram renomeadas para monocinas ao verificar-se que os monócitos e macrófagos também possuíam a capacidade de produção e liberação; entretanto, após a verificação que tecidos e outras estruturas celulares também apresentavam esta capacidade, adotou-se a nomenclatura citocina (Abbas, Lichtman, & Pober, 1998; Cordova-Martinez & Alvarez-Mon, 1999). Neste contexto, é relevante ressaltar que o local de origem de sua produção é determinante para a nomenclatura, por exemplo, as proteínas sinalizadoras quando sintetizadas e liberadas por miócitos são denominadas de miocinas (Pedersen, Akerström, Nielsen, & Fisher, 2007; Pedersen 2011) e pelos adipócitos como adipocinas (Trayhurn, Drevonm, & Eckel, 2011; Trayhurn & Wood, 2004).
As miocinas são sintetizadas e liberadas para a corrente sanguínea por estímulo da contração muscular (Iizuka, Machida, & Hirafuji, 2014) e, sob esta perspetiva, a prática regular de exercício físico possui suma importância na regulação do Imunometabolismo, uma vez que o mesmo é capaz de induzir significantes alterações metabólicas, pois durante a execução dos exercícios há aumento da demanda energética e modificações hormonais para que haja a regulação e manutenção da atividade muscular e, consequentemente, pelo aumento do aporte energético para as células musculares. Frente a este panorama, uma das pesquisas pioneiras que demonstraram a interação entre o sistema imune e metabolismo durante exercício foi conduzida por Bjorn Ahlborg e Gunvor Ahlborg (1970); estes autores associaram a elevação das quantidades das células imunes no sangue durante a sessão de exercício físico com o aumento da resposta adrenérgica, via estímulo simpático do sistema nervoso central, resultando na síntese e liberação de catecolaminas, estas capazes de estimular e disponibilizar substratos para a musculatura esquelética em contração, levando também a uma modulação imunológica local e sistêmica.
Desta forma, fica evidente que em distintos campos, seja na doença ou prática de exercício físico, há íntima comunicação entre sistema imunológico e metabolismo celular, tornando a vertente do Imunometabolismo uma promissora linha de investigação no âmbito do entendimento dos fenômenos que compõe o organismo humano frente a diferentes condições de estímulos estressores. Na figura 1 estão ilustradas as principais descobertas no campo do imunometabolismo que fortaleceram e corroboraram para a formação e consolidação desta área de pesquisa em expansão.
Interação entre Metabolismo, Células Imunológicas e Vias Energéticas
As células do sistema imunológico desempenham papel fundamental na proteção do organismo e manutenção da homeostase corporal compondo uma linha de defesa amplamente mobilizada frente alterações metabólicas, tais como em situações de doença e frente à realização de exercício físico, sendo este último estímulo um agente estressor dependente de sua intensidade, volume e duração. Neste contexto, em meados dos anos 90 foi observado que o exercício físico quando realizado de forma intensa contribuía no aumento da incidência de infeção do trato respiratório superior, enquanto que o exercício de intensidade moderada teria um efeito protetor ao risco de infeções (Nieman, 1994).
Mediante a suscetibilidade às infeções e processos inflamatórios associado com a prática de exercício físico, buscou-se compreender quais possíveis mecanismos modulavam a resposta imunológica, e segundo Costa Rosa e Vaisberge (2002) são mobilizadas do sistema imunológico componentes celulares e humorais em resposta as alterações mecânicas (hipóxia, hipertermia e lesão muscular), metabólicas (glutamina) e hormonais (adrenalina, cortisol, entre outros) impostas pelo exercício.
Frente às alterações impostas pelo esforço físico, diversos ajustes imuno-metabólicos ocorrem a fim de manter o equilíbrio dos sistemas e a execução dos esforços, uma vez que, durante a contração muscular há aumento da demanda energética fazendo-se necessário o aumento da disponibilidade de substratos energéticos, tais como a glutamina, para as células musculares, entretanto, a captação e utilização da glutamina como aporte energético gerou a hipótese de uma competição de substrato entre o músculo esquelético e o sistema imune. De acordo com Pedersen et al. (1998) o músculo esquelético, assim como as células imunes, também utiliza da glutamina como fonte de energia, e em situações nas quais há necessidade de maior substrato energético, como em quadros de sepsemia e exercício físico extenuante em alta intensidade, ocorrerá redução na função de algumas células do sistema imunológico, discutiremos este quadro nos tópicos a seguir.
Metabolismo Energético das Células Imunes
O sistema imunológico possui importância fundamental, não somente para prevenir ou combater uma infeção, mas também na reparação de lesões, proliferação e biossíntese de células e atividade secretória (Calder, Dimitriads, & Newsholme, 2007). Neutrófilos, linfócitos e monócito/macrófagos possuem importantes papeis na resposta imune e inflamatória aguda e crônica.
Os linfócitos são ativados em quadros crônicos de infeção, sendo que os linfócitos T são requeridos para ativar macrófagos, enquanto linfócitos B produzem e secretam anticorpos em resposta ao estímulo antigênico. Já os neutrófilos, constituem 60% dos leucócitos circulantes e atua na fase aguda da infeção, sendo considerados como linha de frente na defesa do organismo por meio da fagocitose e degradação de patógenos. Os macrófagos, assim como os neutrófilos, realizam a fagocitose de patógenos, porém são apresentadores de antígeno por meio do complexo de histocompatibilidade maior II (MHC II) (Newsholme, 2001).
O metabolismo energético das células do sistema imunológico é extremamente refinando, utilizando principalmente glicose, glutamina, ácidos graxos e corpos cetónicos. A glutamina, embora seja um aminoácido não-essencial, exibe fundamental importância nas condições inflamatórias, especialmente para a proliferação e estimulação das funções celulares, inclusive aumentando da produção de citocinas de acordo com o fenótipo de cada célula (Newsholme, 2001).
O linfócito, em um estado quiescente, utiliza o metabolismo oxidativo pela via do ciclo do ácido tri-carboxílico (Ardawi & Newshole, 1984; Norata et al., 2016), entretanto, sob uma determinada ativação, o seu metabolismo sofre mudança brusca, por necessitar de grande demanda energética imediata, ao sair do seu estado de repouso para um estado de proliferação, e passam a produzir energia por meio da glicólise mesmo na presença de oxigênio, o que é chamado de glicólise aeróbia. Ardawi e Newsholme (1984) também verificaram que a glutamina é um substrato importante para proliferação de linfócitos. Desta forma, tanto a glicose, que é convertida em lactato, como a glutamina, que é convertida em glutamato, aspartato e lactato, são os principais substratos dessa célula, embora corpos cetônicos e ácidos graxos também sejam utilizados em menor escala, especialmente em condição homeostática (Ardawi & Newsholme, 1984; Yaqoob, Newsholme, & Calder, 1994). Vale destacar alguns hormônios, como a insulina e hormônios da tireoide, que auxiliam na função dos linfócitos, estimulando a glicólise aeróbica, o consumo de glutamina e estimulação do metabolismo aeróbio (Wasinki et al., 2014).
Nos neutrófilos, a glicose parece ser a principal fonte energética, sendo que a sua oxidação parece ser aumentada durante a fagocitose. Entretanto, a glutamina também exibe importante papel na função desta célula, especialmente quando o fornecimento de glicose é restrito. A carência desses substratos pode prejudicar drasticamente a função dos neutrófilos (Healy, Watson, & Newsholme, 2002).
Os macrófagos, por sua vez, utilizam glicose para a produção de NADPH e precursores de lipídios. Já a glutamina é utilizada para a produção de ATP e a sua escassez pode prejudicar a função dos macrófagos, uma vez que ela potencializa a expressão do MHC II. Os ácidos graxos de cadeia longa são essenciais para gerar intermediários do ciclo do ácido tricarbolxílico (Newsholme, Costa Rosa, Newsholme, & Curi, 1996).
Assim, é nítido que a função do sistema imune está intimamente relacionada com o metabolismo energético. Entretanto é preciso considerar que as funções das diferentes células imunes são associadas com configurações metabólicas distintas (Norata et al., 2015), destacam-se os diferentes fenótipos de macrófagos, sendo os macrófagos do tipo 1 (M1) realizam altas taxas de glicólise e síntese de ácidos graxos, enquanto o fenótipo tipo 2 dos macrófagos (M2) exerce alta demanda na oxidação de ácidos graxos (Mills & ONeil, 2016).
Além disso, a condição metabólica das células imunes pode sofrer reprogramação direcionando para mudanças das propriedades funcionais e viabilizando a alteração do fenótipo celular. Neste contexto, a disponibilidade de nutrientes e sinais induzidos por metabólitos regulam a sua diferenciação fenotípica, por exemplo, o M1/M2 (Norata et al., 2015) e T helper 1/T helper 2 (Caris et al., 2014). Essa mudança do status metabólico sofre influência do seu meio extracelular, que por sua vez, representa alterações sistêmicas advindas de diversas situações, desde estados de doenças à resposta ao exercício físico.
Doenças crônicas e alterações Imunometabólicas
O quadro de doença pode ser caracterizado como um estresse metabólico que culmina em um desequilíbrio das funções do organismo. Desta forma, Norata et al. (2015) consideram que as doenças podem fornecer um profundo entendimento sobre o crosstalk imunometabólico, uma vez que o milieu metabólico está completamente alterado o que impacta todo o sistema imunológico.
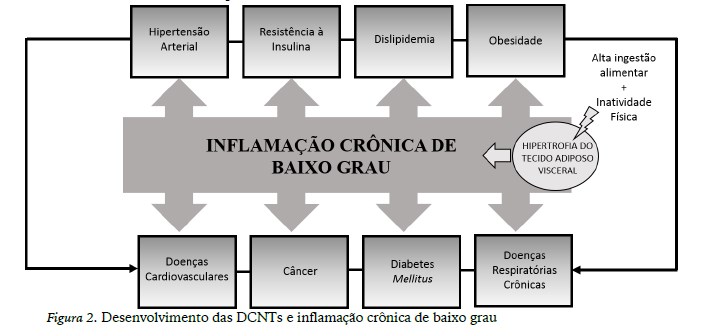
As doenças crônicas não transmissíveis (DCNTs) são doenças multifatoriais, as quais destacam-se as doenças cardiovasculares (hipertensão arterial, arterosclerose, infarto do miocárdio), diabetes mellitus do tipo 2, os diversos tipos de câncer e doenças respiratórias crônicas que possuem como fatores de riscos intermediários outras DCNT, como a hipertensão arterial sistêmica, dislipidemia, obesidade e resistência à insulina (Ouchi, Parker, Lugus, & Walsh, 2011; WHO, 2015).
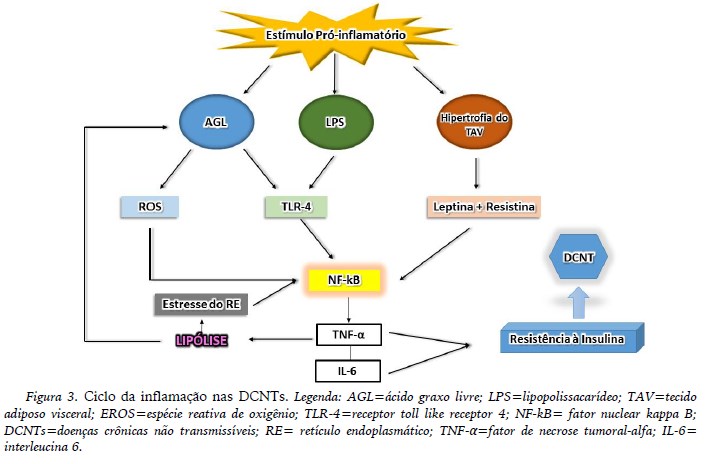
No quadro da obesidade, especialmente o acúmulo de gordura no tecido adiposo visceral (TAV), é considerada o denominador patogênico comum para as DCNTs, pois direciona ao aumento da síntese de adipocinas produzidas principalmente pelos M1 e os diversos fenótipos de linfócitos (T auxiliadores, T citotóxicos, B) residentes no tecido (Ouchi et al., 2011). Dentre as diversas adipocinas destacam-se a IL-6, TNF-α, leptina e resistina. Estas citocinas, produzidas em excesso acarretarão ao aumento da resistência à insulina e lipólise, que em longo prazo poderá contribuir para instalação e/ou progressão de doenças como diabetes mellitus tipo 2, câncer, doenças respiratórias e aterosclerose.
A aterosclerose representa uma das doenças cardiovasculares mais comuns apresentando um processo inflamatório característico à doença no qual ocorrem lesões nos vasos sanguíneos podendo evoluir para a formação de placas fibrosas. Neste processo as células endoteliais secretam proteínas quimioatraentes, como IL-8 e MCP-1, que atraem monócitos/macrófagos para o tecido lesionado estimulando a secreção de outras citocinas pró-inflamatórias, como a IL-1β, IL-3, IL-18 e TNF-α, a fim de restabelecer os danos locais de tais lesões (Golia et al., 2014). Além disso, a relação do aumento de neutrófilos e diminuição de linfócitos tem sido considerada como marcador de eventos cardiovasculares e de mortalidade, já que os neutrófilos estão relacionados com proteínas de fase aguda da inflamação e linfócitos com a formação de aterosclerose (Prats-Puig et al., 2015).
Por outro lado, o processo aterosclerótico apresenta íntima relação com alterações nas concentrações lipídicas circulante, também reconhecida como dislipidemia, e nesta perspectiva nossas investigações evidenciaram recentemente que em adolescentes obesos a ingestão de lipídeos e carboidratos está intimamente associada com a dislipidemia, via alterações nas concentrações de colesterol total, triacilglicerol e HDL-c, e a inflamação crônica de baixo grau, via alterações nas concentrações de IL-1ra, IL-6, adiponectina e insulina (Antunes et al., 2015).
Neste contexto, visto que a maioria das DCNTs citadas parece estar relacionada à inflamação crônica de baixo grau, advinda da inflamação do TAV que ocorre na obesidade, provavelmente o estilo de vida é visto como um dos principais gatilhos para as alterações imunometabólicas, estabelecendo um ciclo vicioso para a inflamação e permanência/manutenção das doenças crônicas (figura 3). Sendo assim, o exercício físico é uma ferramenta estratégica para a prevenção e tratamento de DCNTs, bem como, dos processos inflamatórios que circunda cada tipo de doença.
Exercício Físico: implicações práticas na regulação do Imunometabolismo
A prática insuficiente de exercício físico está relacionada com o desenvolvimento de diversas doenças e alterações metabólicas, tais como diminuição da sensibilidade à insulina, alterações no metabolismo de lipídios, aumento da adiposidade visceral, diminuição da massa corporal magra e perda de força muscular, resultando em um quadro de inflamação crônica de baixo grau (Perdersen, 2009).
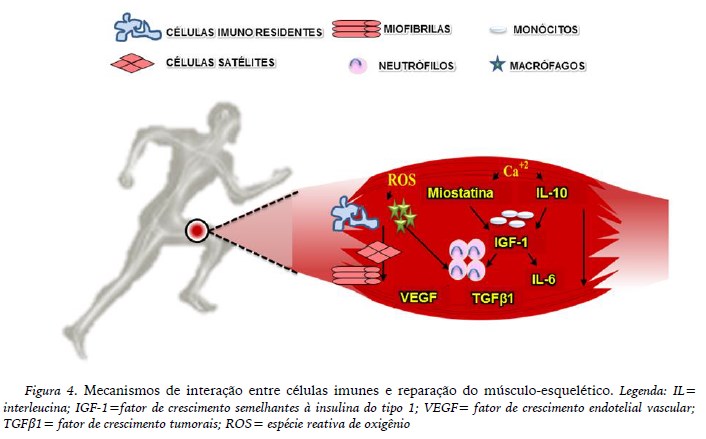
Por outro lado, a contração muscular, advinda do exercício físco, pode elevar a produção de miocinas (tais como miostatina, IL-6, IL-7, IL-8, IL-10, IL-15, LIF) e ativar células do sistema imunológico (monócitos/macrófagos, neutrófilos) a fim de direcionar células satélites para o local do dano muscular com o objetivo de iniciar o processo de remodelamento e reparação tecidual (Tidball & Villalta, 2010) e, desta forma, reverter e equilibrar o possível quadro inflamatório devido sua potente ação anti-inflamatória (Pedersen & Febbraio, 2012). Além disso, o gene da miostatina não ativado pode resultar no aumento da hipertrofia muscular e talvez contribuir para a redução da gordura corporal (Rodgers & Garikipati, 2008).
Em relação à resposta emitida pelas células imunológicas frente ao exercício físico, os neutrófilos são as primeiras células a se instalarem no músculo-esquelético após lesão aguda, pois iniciam o processo de reparação tecidual por meio do aumento na liberação de ROS ou, ainda, por favorecer o recrutamento de monócitos para ação fagocitária e/ou aumento na secreção de citocinas na tentativa de recrutar um número maior de células de defesa (Lockhart & Brooks, 2008).
Os monócitos, após se infiltrarem no tecido alvo, convergem para macrófagos, os quais aumentam a produção de citocinas nos espaços intersticiais, atraindo novas moléculas quimioatraentes de monócitos (Ly-6Cbaixa/ CD11cintermediaria /CX3CR1alta), que induzirão a fagocitose de células musculares apoptóticas e necróticas (Pillon et al., 2013). Os primeiros macrófagos a se acumularem no início da inflamação são caracterizados como M1; quando passam a atuar no processo de recuperação tecidual, mediado por citocinas como IL-10 e fatores de crescimento tumorais (TGFβ1), aumentando a proliferação de mioblastos e crescimento miofibrilar, são polarizados em macrófagos de caráter anti-inflamatório (M2) (Deng, Wehling-Henricks, Villalta, Wang & Tidball, 2012). Os macrófagos M2 têm também a capacidade de produzir fatores de crescimento semelhantes à insulina do tipo 1 (IGF-1) e de crescimento endotelial vascular (VEGF), aumentando o número de vasos sanguíneos, o fluxo de sangue (Wan et al., 2010) e, novamente, induzindo a proliferação de mioblastos e diferenciação de células satélites, os quais resultam na formação de novas miofibrilas (Figura 4) (Rocheteau, Gayraud-Morel, Siegl-Cachedenier, Blasco, & Tajbakhsh, 2012).
Os fatores hormonais também estão intimamente relacionados com a resposta imunometabólica. A adrenalina e o cortisol são dois exemplos de hormônios que respondem ao exercício físico ao mesmo tempo em que modulam o metabolismo de linfócitos, além de ser dependentes da intensidade e duração do estímulo. A adrenalina pode estimular a proliferação de linfócitos além de aumentar o seu consumo de glicose e glutamina, sendo que em exercícios de alta intensidade podem gerar efeito de imunossupressão reduzindo a atividade de enzimas chave que prejudicam o metabolismo de glicose e glutamina. O cortisol, por sua vez, é considerado um hormônio de imunossupressão, especialmente também, em exercícios de alta intensidade, prejudicando a capacidade proliferativa dos linfócitos entre 30 minutos à duas horas após o esforço físico (Wasinki et al., 2014).
Dessa maneira, a manipulação das variáveis do exercício físico, como tipo de estímulo (aeróbio ou anaeróbio), frequência do estímulo (agudo ou crônico), intensidade e volume, contração excêntrica ou concêntrica e intervalos de recuperação, são variáveis que podem influenciar (positiva ou negativamente) o processo de recrutamento de células do sistema imunológico (Gjevestad, Holven & Ulven, 2015; Paulsen, Mikkelsen, Raastad & Peake, 2012).
Em relação ao tipo de estímulo, tem-se observado que os linfócitos circulantes aumentam durante exercício aeróbio de moderada à alta intensidade (van de Vyver, Engelbrecht, Smith & Myburgh, 2015), no entanto, o exercício de alta intensidade e longa duração (ex: maratona), pode suprimir o sistema imune e a quantidade celular pode ficar abaixo dos valores de repouso, como discutido em revisão conduzia por Hoffman-Goetz e Pedersen (1994). Em estudo realizado por van de Vyver et al. (2015), o qual verificou a resposta imune pré, imediatamente pós estímulo e após 4 horas de exercício aeróbio em alta intensidade (12 estímulos de 5 minutos/cada, 10% declive, 15 km/h), observou-se que monócitos e neutrófilos aumentaram após 4 horas do término do exercício, além disso, estes autores sugerem que a presença de monócitos foi responsável pela elevação na concentração de IL-6, via sinalização de STAT-3.
Em relação ao estímulo anaeróbio, que tem como principal tipo o exercício resistido, Stupka, Tarnopolsky, Yardley e Phillips (2001) compararam a resposta da infiltração de células do sistema imune (macrófagos e neutrófilos) e marcador de proteólise [Creatine Kinase (CK)] de acordo com sexo, por meio de biópsia muscular, após três séries de 12 repetições no leg press, com três minutos de intervalo entre as séries, seguida por 10 séries de 10 repetições em cadeira extensora unilateral, com a mesma perna exercitada no leg press (120% de 1RM) e observaram que o número de neutrófilos foi significativamente aumentado em mulheres após 24 horas, mas não no sexo masculino, sendo que os macrófagos aumentaram em ambos os sexos, demostrando que o a resposta inflamatória pode ser diferente de acordo com o tempo de estímulo excêntrico e o gênero, no entanto, ainda não está bem esclarecida a influência do gênero neste desfecho (St. Pierre, Correia & Cannon, 1999).
Nosso grupo investigou, recentemente, os efeitos do treinamento físico aeróbio de intensidade moderada combinado com o treinamento resistido e concluímos que, independentemente do tipo de periodização (linear ou ondulatória), adolescentes obesos apresentaram aumento das concentrações de adiponectina quando comparados com aqueles que realizaram apenas o treinamento aeróbio, havendo, portanto, melhora no quadro de resistência à insulina e, consequentemente, do milieu inflamatório da obesidade (Inoue et al., 2015).
Quando analisada a frequência do estímulo (agudo ou crônico), observa-se que as células imunes instalam-se no tecido muscular imediatamente após uma única sessão de exercício físico. Exercícios de curta duração e de intensidade entre moderada e elevada (3 séries de 10 repetições e 70% de 1RM ou 30 minutos de corrida a 75% do VO2max) podem resultar em picos na expressão de IL-6, IL-8 e TNF-α RNAm de 2 a 12 horas (Louis, Raue, Yang, Jemiolo, & Trappe, 2007). Simonson e Jackson (2004), analisaram os efeitos de uma série de exercício resistido sobre a resposta imune em homens ativos (3 séries de 8-10 repetições a 75% de 1 RM para oito grandes agrupamentos musculares) e concluíram que os leucócitos, principalmente os monócitos e neutrófilos, aumentaram imediatamente pós-exercício, exceto basófilos e eosinófilos, e retornaram as concentrações pré-exercício após 15 e 30 minutos do término da sessão, no entanto, os neutrófilos permaneceram elevados mesmo após 30 minutos pós-exercício.
Apesar de serem escassos na literatura estudos que verificaram a resposta imune após longos períodos de treinamento, Yakeu et al. (2010) investigaram os efeitos de oito semanas de treinamento leve (caminhada=10.000 passos/dia, três vezes/semana) em indivíduos sedentários e observaram que até mesmo o exercício de baixa intensidade tem associação positiva com macrófagos M2, Th2, PGC-1α e PGC-1β e negativa com macrófagos M1, demonstrando que o exercício em intensidade leve, porém realizado consistentemente, pode polarizar os macrófagos.
Sobre a intensidade e volume, em estudo clássico realizado por Tvede et al. (1989), o qual verificou a influência de diferentes intensidades de exercício em ciclo ergómetro (25%, 50% e 75% do VO2max) durante 1 hora, foi observado que algumas células de defesa [Natural Killer (NK) e células Killer Linfócitos ativadas (LAK)] foram aumentadas em todas as intensidades de esforço, no entanto, foram suprimidas na carga de 75% do VO2max. Além disso, somente essa intensidade apresentou monócitos infiltrados após o término do exercício, sugerindo que essa intensidade de exercício pode potencializar o processo de remodelamento da musculatura esquelética.
Fragala et al. (2011) analisaram a expressão do recetor adrenérgico de β2 (β2-ADR) em monócitos, granulócitos e linfócitos e observou-se que em linfócitos houve aumento da expressão após exercício resistido severo (6 séries de 5 RM no agachamento) e concluíram que em monócitos e granulócitos há diminuição durante o exercício, porém com elevação durante o processo de recuperação. Em adição, estudos do nosso grupo têm demostrado que exercício exaustivo pode promover aumento de citocinas (IL-6, IL-10, TNF-α) e assim, via TLR-4, contribuir para mobilização de substrato energético (Rosa Neto et al., 2011). Como citado anteriormente no estudo de Yakeu et al. (2010), o exercício físico leve, realizado sistematicamente, também foi suficiente para induzir benefícios anti-inflamatórios. Em contrapartida, em uma situação de overtraining, pode haver aumento exacerbado de citocinas pró-inflamatórias sintetizadas e liberadas pelo tecido adiposo (adipocinas), induzindo um estado de inflamação tecidual (Lira et al., 2010a).
Em relação ao intervalo de recuperação, nosso grupo verificou a influência de 30 e 90 segundos de intervalo em jovens saudáveis submetidos a 4 séries de agachamento seguido por 4 séries de supino horizontal, com 70% de 1RM até a falha do movimento; concluímos que apesar do desempenho diminuir em ambos os intervalos, somente o tempo de 90 segundos induziu aumento nas concentrações de IL-6 e redução de IL-10, porém sem significância estatística nessa última, assim, intervalos moderados de recuperação poderiam favorecer uma maior resposta metabólica e benefícios anti-inflamatórios durante o processo de reparação tecidual (Rossi, Gerosa-Neto, Zanchi, Cholewa, & Lira, 2016).
Apesar de o sistema imune e o metabolismo serem de extrema importância tanto na resolução de um quadro inflamatório crônico de baixo grau (obesidade, DM2) como no remodelamento da musculatura esquelética, a relação imunometabolismo e exercício deve ser interpretada com cautela, uma vez que os níveis de aptidão física dos sujeitos, faixa etária, gênero, tipo de análise ou técnica empregada diferem significativamente entre os estudos, entretanto, tanto indivíduos altamente treinados (Lira et al., 2010b), moderadamente ativos (Diniz et al., 2015) ou inclusive que apresentam um quadro de doença, como a caquexia associada ao câncer (Lira et al., 2014), podem apresentar efeitos anti-inflamatórios induzidos pelo exercício físico regular e menor risco de dislipidemia em relação a indivíduos sedentários (Lira et al., 2014).
Perspetivas futuras
A inter-relação entre as respostas inflamatória e metabólica é determinante para a instalação de diferentes doenças, por outro lado, a mesma também é determinante para as respostas anti-inflamatória e anti-aterogênica mediadas pelo exercício físico (Lira et al. 2014a; Lira et al 2014b). Estudos que explorem os mecanismos envolvidos nesta inter-relação são fundamentais para o entendimento desta nova área do conhecimento científico.
CONSIDERAÇÕES FINAIS
Mediante ao exposto acima, podemos concluir que há uma interação entre sistema imunológico, especialmente a resposta inflamatória, e o metabolismo celular em diferentes condições, seja frente às doenças crônicas ou exercício físico. Torna-se de suma importância a compreensão dos mecanismos de interação entre os sistemas, sendo esta fundamental para a utilização destas informações em prol da promoção da qualidade de vida, na elaboração efetiva de programas de prevenção e tratamento de doenças, bem como na melhora do desempenho físico.
REFERÊNCIAS
Abbas, A. K., Lichtman, A. H., & Pober, J. S. (1998). Citocinas. In: Imunologia celular e molecular (2nd ed.) Rio de Janeiro: Revinter. [ Links ]
Ahlborg, B., & Ahlborg, G. (1970). Exercise leukocytosis with and without beta-adrenergic blockade. Acta medica scandinavica, 187(4), 241-246. [ Links ]
Antunes, B. M. M., Monteiro, P. A., Silveira, L. S., Brunholi, C. C., Lira, F. S., & Freitas Júnior, I. F. (2015). Macronutrient intake is correlated with dyslipidemia and low-grade inflammation in childhood obesity but mostly in male obese. Nutrición Hospitalaria, 32(3), 997-1003. [ Links ]
Ardawi, M. S. M., & Newsholme, E. A. (1984). Intracellular localization and properties of phosphate-dependent glutaminase in rat mesenteric lympy nodes. Biochemical Journal, 217(1), 289-296. [ Links ]
Bhatt, S. P., & Dransfield, M. T. (2013) Chronic obstructive pulmonary disease and cardiovascular disease. Translational Research, 162(4), 237-251. [ Links ]
Calder, P. C., Dimitriadis, G., & Newsholme, P. (2007). Glucose metabolism in lymphoid and inflammatory cells and tissues. Current Opinion in Clinical Nutrition and Metabolism Care, 10(4), 531-40. [ Links ]
Candore, G., Caruso, C., & Colonna-Romano, G. (2010). Inflammation, genetic background and longevity. Biogerontology, 11(5), 565-573. [ Links ]
Caris, A. V., Lira, F. S., de Mello, M. T., Oyama, L. M., & Santos, R. V. T. (2014). Carbohydrate and Glutamine supplementation modulates the Th1/Th2 balance after performed at stimulated altitude of 4500m. Nutrition, 30(11-12),1 331-1336. [ Links ]
Catalán, V., Goméz-Ambrosi, J., Rodriguez, A., & Frühbeck, G. (2013). Adipose tissue immunity and cancer. Frontiers Physiology, 4, 275. [ Links ]
Chawla, A., Nguyen, K. D., & Sharon Goh, Y. P. (2012). Machrophage-mediated inflammation in metabolic disease. Nature Reviews Immunology, 11(11), 738-749. [ Links ]
Chen, L., Chen, R., Wang, H., & Liang, F. (2015). Mechanisms linking inflammation to insulin resistance. International Journal of Endocrinology, 2015, 508409. [ Links ]
Cordova Martinez, A., & Alvarez-Mon, M. (1999). O sistema imunológico (I): conceitos gerais, adaptação ao exercício físico e implicações clínicas [Versão eletrônica]. Revista Brasileira de Medicina do Esporte, 5(3), 120-125. [ Links ]
Costa Rosa, L. F. P. B., & Vaisberg, M. W. (2002). Influências do exercício na resposta imune. Revista Brasileira de Medicina do Esporte, 8(4), 167-172. [ Links ]
Deng, B., Wehling-Henricks, M., Villalta, S. A., Wang, Y., & Tidball, J. G. (2012). IL-10 triggers changes in macrophage phenotype that promote muscle growth and regeneration. Journal of Immunology, 189(7), 3669–3680. [ Links ]
DiGimolamo, M., Fine, J. B., Tagra, K., & Rossmanith, R. (1998). Qualitative regional differences in adipose tissue growth and cellularity in male Wistar rats fed ad libitum. American Journal of Physiology, 274(5 Pt 2), R1460-467. [ Links ]
Diniz, T. A., Fortaleza, A. C., Buonani, C., Rossi, F. E., Lira, F. S., & Freitas-Júnior, I. F. (2015). Relationship between moderate-to-vigorous physical activity, abdominal fat and immunometabolic markers in postmenopausal women. European Journal of Obstetrics Gynecology and Reproductive Biology, 194, 178-182. [ Links ]
Fragala, M. S., Kraemer, W. J., Mastro, A. M., Denegar, C. R., Volek, J. S., Häkkinen, K., Anderson, J. M., Lee, E. C, & Maresh, C. M. (2011). Leukocyte β2-adrenergic receptor expression in response to resistance exercise. Medicine and Science in Sports and Exercise, 43(8), 1422-1432. [ Links ]
Ghigliotti, G., Barisione, C., Garibaldi, S., Fabbi, P., Brunelli, C., Spallarossa, P., Altieri, P., Rosa, G., Spinella, G., Palombo, D., Arsenescu, R., & Arsenescu, V. (2014). Adipose tissue immune response: novel triggers and consequences for chronic inflammatory conditions. Inflammation, 37(4), 1337-1353. [ Links ]
Gjevestad, G.O., Holven, K.B., & Ulven, S.M. (2015). Effects of exercise on gene expression of inflammatory markers in human peripheral blood cells: a systematic review. Current Cardiovascular Risk Reports, 9(7), 34. [ Links ]
Golia, E., Limongelli, G., Natale, F., Fimiani, F., Maddaloni, V., Pariggiano, I., Bianchi, R., Crisci, M., D'Acierno, L., Giordano, R., Di Palma, G., Conte, M., Golino, P., Russo, M.G., Calabrò, R., & Calabrò, P. (2014). Inflammation and cardiovascular disease: from pathogenesis to therapeutic target. Current Atherosclerosis Reports, 16(9), 435. [ Links ]
Gruchała-Niedoszytko, M., Małgorzewicz, S., Niedoszytko, M., Gnacińska, M., & Jassem, E. (2013). The influence of obesity on inflammation and clinical symptoms in asthma. Advances in Medical Science, 58(1), 15-21. [ Links ]
Healy, D. A., Watson, R. W. G., & Newsholme, P. (2002). Glucose, but not glutamine, protects against spontaneous and anti-Fas antibody-induced apoptosis in human neutrophilis. Clinical Science, 103(2), 179-189. [ Links ]
Hoffman-Goetz, L., & Pedersen, B. K. (1984). Exercise and the immune system: a model of the stress response? Immunology today, 5(8), 382-387. [ Links ]
Husain, K., Hernandez, W., Ansari, R.A., & Ferder, L. (2015). Inflammation, oxidative stress and renin angiotensin system in atherosclerosis. World Journal of Biololgy Chemistry, 6(3), 209-217. [ Links ]
Iizuka, K., Machida, T., & Hirafuji, M. (2014) Skeletal muscle is an endocrine organ. Journal of Pharmacological Science, 125(2), 125-131. [ Links ]
Inoue, D. S., de Mello, M. T., Foschini, D., Lira, F. S., de Piano Ganen, A., da Silveira Campos, R. M., de Lima Sanches, P., Silva, P. L., Corgosinho, F. C., Rossi, F. E., Tufik, S, & Dâmaso, A. R. (2015). Linear and undulating periodized strength plus aerobic training promote similar benefits and lead to improvement of insulin resistance on obese adolescents. Journal of Diabetes and its Complications, 29(2), 258-264. [ Links ]
Lee, B. C., & Lee, J. (2014) Cellular and molecular players in adipose tissue inflammation in the development of obesity-induced insulin resistance. Biochimica et Biophyssica Acta, 1842(3), 446-462. [ Links ]
Lira, F. S., Neto, J. C., & Seelaender, M. (2014a). Exercise training as treatment in cancer cachexia. Applied Physiology, Nutrition, and Metabolism, 39(6), 679-686. [ Links ]
Lira, F. S., Rosa Neto, J. C., Antunes, B. M., & Fernandes, R. A. (2014). The relationship between inflammation, dyslipidemia and physical exercise: from the epidemiological to molecular approach. Current Diabetes Review, 10(6), 391-396. [ Links ]
Lira, F. S., Rosa, J. C., Lima-Silva, A. E., Souza, H. A., Caperuto, E. C., Seelaender, M. C., Santos, R. V. T. (2010). Sedentary subjects have higher PAI-1 and lipoproteins levels than highly trained athletes. Diabetology & Metabolic Syndrome, 2, 7. https://doi.org/10.1186/1758-5996-2-7 [ Links ]
Lira, F. S., Rosa, J. C., Pimentel, G. D., Tarini V. A., Arida R. M., Faloppa F., Alves, E. S., do Nascimento, C. O., Oyama, L. M., Seelaender, M., de Mello, M. T., & Santos, R.V. (2010a). Inflammation and adipose tissue: effects of progressive load training in rats. Lipids in Health and Disease, 9, 109. [ Links ]
Lockhart, N. C., & Brooks, S. V. (2008). Neutrophil accumulation following passive stretches contributes to adaptations that reduce contraction-induced skeletal muscle injury in mice. Journal of Applied Physiology, 104(4), 1109–1115. [ Links ]
Louis, E., Raue, U., Yang, Y., Jemiolo, B., & Trappe, S. (2007). Time course of proteolytic, cytokine, and myostatin gene expression after acute exercise in human skeletal muscle. Journal of Applied Physiology, 103(5),1744–1751. [ Links ]
Lumeng, C. N., Bodzin, J. L., & Saltiel, A. R. (2007). Obesity induces a phenotypic switch in adipose tissue macrophage polarization. The Journal of Clinical Investigation, 117(1), 175-184. [ Links ]
Mathis, D., & Shoelson, S. E. (2011). Immunometabolism: an emerging frontier. Nature Reviews. Immunology, 11(2), 81. [ Links ]
McNelis, J. C., & Olefsky, J. M. (2014). Macrophages, immunity, and metabolic disease. Immunity, 41(1), 36-48. [ Links ]
Mills, E. L., & O´Neil, L. A. (2016). Reprogramming mitochondrial metabolism in machrophage as an anti-inflammatory signal. European Journal of Immunology, 46(1), 13-21. [ Links ]
Mraz, M., & Haluzik, M. (2014). The role of adipose tissue immune cells in obesity and low-grade inflammation. The Journal of Endocrinology, 222(3), R113-127. [ Links ]
Newsholme, P. (2001). Why is l-glutamine metabolism important to cells of immune system in health, post injury, surgery or infection? The Journal of Nutrition, 131(Suppl.9), 2515S-2522S. [ Links ]
Newsholme, P. Costa Rosa, L. F. B. P., Newsholme, E. P., & Curi, R. (1996). The importance of fuel metabolism to machrophage function. Cell Biochemistry and Function, 14(1), 1-10. [ Links ]
Nieman, D. C. (1994). The effect of exercise on immune function. Bulletin on the Rheumatic Diseases, 43(8), 5-8. [ Links ]
Nimmo, M. A., Leggate, M., Viana, J. L., & King, J. A. (2013). The effect of physical activity on mediators of inflammation. Diabetes, Obesity and Metabolism, 15(suppl.3), 51-60. [ Links ]
Norata, G. D., Caligiuri, G., Chavakis, T., Matarese, G., Netea, M. G., Nicoletti, A., ONeill, L. A., & Merelli-Berg, F. M. (2015). The cellular and molecular basis of translational immunometabolism. Immunity, 43(15), 421-434. [ Links ]
Ohashi, K., Shibata, R., Murohara, T., & Ouchi, N. (2014). Role of anti-inflammatory adipokines in obesity-related diseases. Trends in Endocrinology and Metabolism, 25(7), 348-355. [ Links ]
Olefsky, J. M., & Glass, C. K. (2010). Macrophages, inflammation, and insulin resistance. Annual Review of Physiology, 72,219-246. [ Links ]
Ouchi, N., Parker, J. L., Lugus, J. J., & Walsh, K. (2011). Adipokines in inflammation and metabolic disease. Nature Reviews. Immunology, 11(2),85-97. [ Links ]
Paulsen, G., Mikkelsen, U. R., Raastad, T., & Peake, J. M. (2012). Leucocytes, cytokines and satellite cells: what role do they play in muscle damage and regeneration following eccentric exercise? Exercise Immunology Review, 18, 42–97. [ Links ]
Pedersen, B. K. (2009). The diseasome of physical inactivity – and the role of myokines in muscle–fat cross talk. The Journal of Physiology, 587(Pt23), 5559–5568. [ Links ]
Pedersen, B. K. (2011). Muscles and their myokines. The Journal of Experimental Biology, 214(Pt 2), 337-346. [ Links ]
Pedersen, B. K., & Febbraio, M. A. (2012). Muscles, exercise and obesity: skeletal muscle as a secretory organ. Nature Reviews. Endocrinology, 8(8), 457-465. [ Links ]
Pedersen, B. K., Akerström, T. C., Nielsen, A. R., & Fischer, C. P. (2007). Role of myokines in exercise and metabolism. Journal of Applied Physiology (1985), 103(3), 1093-1098. [ Links ]
Pedersen, B. K., Rohde, T., & Ostrowski, K. (1998). Recovery of the immune system after exercise. Acta Physiologica Scandinavica, 162(3), 325-332. [ Links ]
Periyalil, H. A., Gibson, P. G., & Wood, L. G. (2013). Immunometabolism in Obese asthmatic: Are we there yet? Nutrients, 5(9), 3506-30. [ Links ]
Petersen, A., Marie, W., & Pedersen, B. K. (2005). The anti-inflammatory effect of exercise. Journal of applied physiology (1985), 98(4), 1154-1162. [ Links ]
Pillon, N.J., Bilan, P.J., Fink, L.N., & Klip, A. (2013). Cross-talk between skeletal muscle and immune cells: muscle-derived mediators and metabolic implications. American Journal of Physiology, Endocrinology and Metabolism, 304(5), E453-E465. [ Links ]
Pizza, F. X., Baylies, H., & Mitchell, J. B. (2001). Adaptation to eccentric exercise: neutrophils and E-selectin during early recovery. Canadian Journal of Applied Physiology, 26(3), 245-253. [ Links ]
Prats-Puig, A., Gispert-Saüch, M., Díaz-Roldán, F., Carreras-Badosa, G., Osiniri, I., Planella-Colomer, M., de Zegher, F., Ibánez, L., Bassols, J., & López-Bermejo, A. (2015). Neutrophil-to-lymphocite ratio: an inflammatory marker related to cardiovascular risk in children. Thrombosis and Haemostasis, 114(4), 727-734. [ Links ]
Ringseis, R., Eder, K., Mooren, F. C., & Krüger, K. (2015). Metabolic signals and innate immune activation in obesity and exercise. Exercise Immunology Review, 21, 58-68. [ Links ]
Rocheteau, P., Gayraud-Morel, B., Siegl-Cachedenier, I., Blasco, M. A., & Tajbakhsh, S. (2012). A subpopulation of adult skeletal muscle stem cells retains all template DNA strands after cell division. Cell, 148(1-2), 112–125. [ Links ]
Rodgers, B. D., & Garikipati, D. K. (2008). Clinical, agricultural, and evolutionary biology of myostatin: a comparative review. Endocrinoly Review, 29(5), 513–534. [ Links ]
Rosa Neto, J. C., Lira, F. S., Eguchi, R., Pimentel, G. D., Venâncio, D. P., Cunha, C. A, Oyama, L. M., De Mello, M. T., Seelaender, M., & do Nascimento, C.M. (2011). Exhaustive exercise increases inflammatory response via Toll like receptor-4 and NF-κBp65 pathway in rat adipose tissue. Journal of Cellular Physiology, 226(6), 1604-1607. [ Links ]
Rosa Neto, J. C., Lira, F. S., Oyama, L. M., Zanchi, N. E., Yamashita, A. S., Batista, M. L. Jr., Oller do Nascimento, C. M., & Seelaender, M. (2009). Exhaustive exercise causes an anti-inflammatory effect in skeletal muscle and a pro-inflammatory effect in adipose tissue in rats. European Journal of Applied Physiology, 106(5), 697-704.
Rossi, F. E., Gerosa-Neto J., Zanchi, N. E. Cholewa, J. M., & Lira, F.S (2016). Impact of short and moderate rest intervals on the acute immunometabolic response to exhaustive strength exercise: part I. Journal of Strength and Conditioning Research, 30(6), 1563-1569. [ Links ]
Schipper, H. S., Prakken, B., Kalkhoven, E., & Boes, M. (2012). Adipose tissue-resident immune cells: key players in immunometabolism. Trends in Endocrinology and Metabolism, 23(8), 407-415. [ Links ]
Seelaender, M., Batista, M. Jr., Lira, F., Silverio, R., & Rossi-Fanelli, F. (2012). Inflammation in cancer cachexia: to resolve or not to resolve (is that the question?). Clinical Nutritional, 31(4), 562-566. [ Links ]
Simonson, S. R., & Jackson, C. G. (2004). Leukocytosis occurs in response to resistance exercise in men. Journal of Strength and Conditioning Research, 18(2), 266-271. [ Links ]
St. Pierre Schneider, B., Correia, L. A. & Cannon, J.G. (1999). Sex differences in leukocyte invasion in injured murine skeletal muscle. Research in Nursing & Health, 22(3), 243–250. [ Links ]
Steensberg, A., Fischer, C. P., Keller, C., Møller, K., & Pedersen, B. K. (2003). IL-6 enhances plasma IL-1ra, IL-10, and cortisol in humans. American Journal of Physiology, Endocrinology and Metabolism, 285(2), E433-737. [ Links ]
Stupka, N., Tarnopolsky, M. A., Yardley, N. J., & Phillips, S. M. (2001). Cellular adaptation to repeated eccentric exercise-induced muscle damage. Journal of Applied Physiology (1985), 91(4), 1669-1678. [ Links ]
Tidball, J. G., & Villalta, S. A. (2010). Regulatory interactions between muscle and the immune system during muscle regeneration. American Journal of Physiology Regulatory, Integrative and Comparative Physiology, 298(5), R1173–R1187. [ Links ]
Trayhurn, P., & Wood, I. S. (2004). Adipokines: inflammation and the pleiotropic role of white adipose tissue. The British Journal of Nutrition, 92(3), 347–355. [ Links ]
Trayhurn, P., Drevonm C. A., & Eckel, J. (2011). Secreted proteins from adipose tissue and skeletal muscle - adipokines, myokines and adipose/muscle cross-talk. Archives of Physiology and Biochemistry, 117(2), 47-56. [ Links ]
Tvede, N., Pedersen, B. K., Hansen, E. R., Bendix, T., Christensen, L. D., Galbo, H., & Halkjaer-Kristensen, J. (1989). Effect of physical exercise on blood mononuclear cell subpopulations and in vitro proliferative responses. Scandinavian Journal of Immunology, 29(3), 383-389. [ Links ]
van de Vyver, M., Engelbrecht, L., Smith, C., & Myburgh, K. H. (2016). Neutrophil and monocyte responses to downhill running: Intracellular contents of MPO, IL-6, IL-10, pstat3, and SOCS3. Scandinavian Journal of Medicine & Science in Sports, 26(6), 638-647. [ Links ]
Wan, C., Shao, J., Gilbert, S. R., Riddle, R. C., Long, F., Johnson, R. S., Schipani, E., & Clemens, T.L. (2010). Role of HIF-1 alpha in skeletal development. Annals of the New York Academy of Science, 1192, 322–326. [ Links ]
Wasinki, F., Gregnani, M. F., Ornellas, F. H., Bacuraru, A. V. N., Câmara, N. O., Araújo, R. C., & Bacurau, R. F. (2014). Lynphocyte glucose and glutamine metabolism as targets of the anti-inflammatory effects of exercise. Mediators of Inflammation, 2014, 326803. [ Links ]
Wellen, K. E., & Hotamisligil, G. S. (2005). Inflamation, stress, and diabetes. The Journal of Clinical Investigation, 115(5), 1111-1119. [ Links ]
World Health Organization (2015). Global status report on non-communicable diseases 2014. Geneva: World Health Organization [ Links ]
Yakeu, G., Butcher, L., Isa, S., Webb, R., Roberts, A. W., Thomas, A. W., Backx, K., & Morris, K. (2010). Low-intensity exercise enhances expression of markers of alternative activation in circulating leukocytes: roles of PPARgamma and Th2 cytokines. Atherosclerosis, 212(2), 668–673. [ Links ]
Yaqoob, P., Newsholme, E. A., & Calder, P. C. (1994). Fatty oxidation by lymphocytes. Biochemical Society Transactions, 22(2), 116S. [ Links ]
Yeung, C. Y., Tso, A. W. K., Xu, A., Wang, Y., Woo, Y. C., Lam, T. H., Lo, S. V., Fong, C. H., Wat, N. M., Woo, J., Cheung, B. M., & Lam, K.S. (2013). Pro-Inflammatory adipokines as predictors of incident cancers in a Chinese cohort of low obesity prevalence in Hong Kong. Plos One, 8(10), e78594. [ Links ]
Agradecimentos:
Nada a declarar
Conflito de Interesses:
Nada a declarar.
Financiamento:
Fabio Santos Lira thanks CNPq for their support (305263/2015-3).
Artigo recebido a 15.01.2016; Aceite a 05.06.2016
* Autor correspondente: Universidade Estadual Paulista, Departamento de Educação Física, Rua Roberto Simonsen, 305, CEP. 19060-900, Presidente Prudente, SP-Brazil. E-mail: ba.antunes2@gmail.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}