










Serviços Personalizados
Journal
Artigo
 Francês (pdf)
Francês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkSilva Lusitana
versão impressa ISSN 0870-6352
Silva Lus. vol.21 no.2 Lisboa dez. 2013
Effet du Climat sur l'Évolution des Populations de Varroa destructor chez l'Abeille Apis mellifera intermissa L. dans les Différents Écosystèmes de l'Algérie
Efeito do clima na evolução de populações de Varroa destructor em diferentes ecossistemas argelinos
Variations in the dynamics of development of Varroa destructor populations from the effects of climate in northern Algeria
*Berkani-Ghalem Zohra, **Hámi Hálima et ***Berkani Mohamed Laïd
*Maître de Conférences, Enseignante-chercheuse, École Normale Supérieure. Département de biologie. B.P. 92 Kouba-Alger 16050 -Kouba-ALGÉRIE ghalemzy@yahoo.fr
**Doctorante, École Nationale Supérieure Vétérinaire. BP 161.HacèneBadi .16200.El Harrach. Alger. ALGÉRIE hamihalima@gail.com
***Maître de Conférences, enseignant-chercheur, École Nationale Supérieure Agronomique - Département de Zootechnie- Laboratoire d'Apiculture. 16200 HacèneBadi. El Harrach. Alger. ALGÉRIE berkani_ml@hotmail.com
RÉSUMÉ
Cette étude porte sur la dynamique des populations de Varroa destructor dans trois écosystèmes du nord d'Algérie à climat méditerranéen humide ou Mitidja, continentale semi-aride des hauts plateaux et continentale aride de la steppe. Des observations mensuelles ont été effectuées, de janvier 2010 à aout 2012, sur 18 colonies réparties entre 3 ruchers tout au long de la bande nord de l'Algérie. Les populations d'abeilles n'ont pas subi de traitement acaricide. Le développement des populations d'acariens est étudié en tenant compte de la mortalité naturelle, du taux d'infestations des abeilles adultes et du taux d'infestation du couvain. Pour la mortalité naturelle, il y a des différences significatives entre les années où on enregistre une évolution importante entre les mois de mars et août avec un maximum de 577, 494 et 373 respectivement pour la Mitidja, les hauts plateaux et la steppe. Nous avons constaté une régression qui s'annule de septembre à février. Les taux d'infestation des abeilles adultes pour les trois types de climat sont respectivement de: 6,13; 6,09 et 4,81% pour 100 abeilles ouvrières. Il en est de même pour le maximum des taux d'infestation du couvain qui varie en fonction de la zone climatique. Les valeurs globales pour les trois écosystèmes étudiés s'établissent à: 7,16 pour la Mitidja, 6,55 pour les hauts plateaux et 5,39% pour la steppe. Ces valeurs baissent en passant du climat humide au sec. Les taux d'infestation diminuent sensiblement dans le temps.
Mots clés: abeille, Varroa destructor, évolution de la population, Apis mellifera intermissa, climat
SUMÁRIO
Este estudo debruça-se sobre a dinâmica das populações de Varroa destructor em três ecossistemas do norte da Argélia, com clima mediterrânico húmido como em Mitidja, continental semi-árido nos altos planaltos e continental árido na estepe. Efectuaram-se observações mensais, de janeiro de 2010 a agosto de 2012, em 18 colónias repartidas por três apiários ao longo da costa norte da Argélia. As populações de abelhas não foram tratadas com acaricidas. O desenvolvimento das populações de ácaros é estudado tendo em conta a mortalidade natural, a taxa de infestação das abelhas adultas e a taxa de infestação das larvas. Quanto à mortalidade natural, existem diferenças significativas entre os anos, e regista-se uma evolução importante entre os meses de março e agosto, com um máximo de 577, 494 e 373 respetivamente para Mitidja, os altos planaltos e a estepe. Constatámos uma regressão, que se anula de setembro a fevereiro. Nos três tipos de clima, a taxa de infestação das abelhas adultas é de, respetivamente, 6,13 ; 6,09 e 4,81% por 100 abelhas obreiras. O mesmo se passa com a taxa de infestação das larvas, que varia em função da zona climática. Os valores globais para os três ecossistemas estudados são os seguintes: 7,16 para Mitidja, 6,55 para os altos planaltos e 5,39% para a estepe. Estes valores baixam ao passar do clima húmido ao seco. As taxas de infestação diminuem sensivelmente com o passar do tempo.
Palavras-chave: abelha, Varroa destructor, evolução da população, Apis melífera intermissa, clima
ABSTRACT
This experiment is performed on the dynamics of the population of the ectoparasitic Varroa destructor in northern Algeria where there are climatic differences from north to south. Monthly observations were conducted from January 2010 to August 2012, in 18 colonies in three apiaries distributed throughout the northern strip of Algeria. Bee populations have not undergone any acaricide treatment. Our experiment is undertaken in the areas corresponding to the following three climatic regions: Mediterranean climate or wet Mitidja (CM), Continental semiarid highlands (CSA) and continental arid steppe (CA). The development of mite populations is studied taking into account the following parameters: natural mortality (M) rates infestations of adult bees (TIA) and infestation rate of brood (ICT). For natural mortality, there are significant differences between the years, there has been a significant change during the period from March to August with a maximum of infestation of 577, 494 and 373 respectively for Mitidja highlands and steppe, which decreases to cancel from September to February. The infection rate of bees (TIA) for the three types of climate is respectively: 6.13, 6.09 and 4.81%. It is the same for the maximum infestation rate of brood, which varies depending on the climate zone. Global values for the three ecosystems studied are as follows: 7.16% for Mitidja, 6.55% for the highlands and 5.39% for the steppe. These values decrease from humid to dry climate. Infection rates decreased significantly over time.
Key words: honeybee, Varroa destructor, population evolution, Apis mellifera intermissa, climate
Introduction
Varroa destructor, appelé communément varroa, est un acarien parasite de l'abeille adulte, des larves et des nymphes. Il provoque des pertes économiques importantes en apiculture et il est l'une des causes de la perte du cheptel apicole dans le monde. Aujourd'hui, dans les cinq continents pas un rucher ne lui échappe. La transhumance, les transactions commerciales des colonies et des essaims d'un pays à un autre, font que son extension est devenue mondiale (DE JONG et al., 1982). En Algérie, toutes les tentatives de traitements chimiques et naturels ont été utilisées depuis qu'il est apparu il y a trente ans (DEFAVEAUX, 1981; BERKANI et GHALEM, 2009). Malgré l'emploi régulier de différentes molécules acaricides, la varroase reste toujours présente. Dans cette étude, nous avons tenté d'étudier l'impact du climat sur la dynamique des populations de cet ectoparasite dans quelques écosystèmes du nord d'Algérie.
Matériel et methods
Dix huit colonies d'abeilles sont choisies et mises en place dans trois régions caractérisées par des climats différents. Les stations d'observation sont réparties ainsi et comportent chacune six colonies (Figure 1):
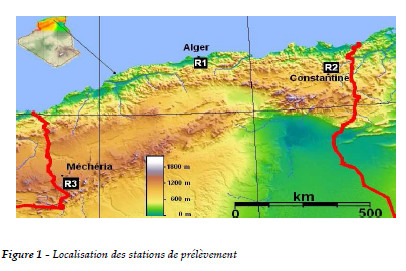
-R1 - La plaine orientale de la Mitidja (littoral) se caractérise par un climat de type méditerranéen aux hivers froids et pluvieux et des étés secs et chauds. Elle se caractérise par une richesse mellifère qui s'étale sur toute l'année.
-R2 - Les hauts plateaux constantinois à climat semi-aride. Les hivers sont très froids et peu humides et les étés secs et très chauds (SELTZER, 1946).
-R3 - La zone steppique a un climat aride aux hivers froids et peu arrosés et des étés très chauds et secs.
Les prélèvements d'échantillons sont effectués entre janvier 2010 et août 2012. Toutes les colonies étudiées sont issues d'essaims précoces de l'année précédente avec des reines jeunes et indemnes de varroase. Durant nos travaux, les ruches n'ont subi aucune thérapie à base d'acaricides et cela dans le but de voir la progression naturelle de Varroa sous les différents climats. Les colonies sont échantillonnées mensuellement et à la deuxième année, elles sont toutes rémérées par de nouvelles reines.
Trois paramètres sont ciblés dans ce travail. La mortalité naturelle des varroas (M) qui est le nombre d'acariens morts naturellement par colonie et par mois. Le second est le taux d'infestation des ouvrières par le varroa (TIA). Chaque mois 100 à 150 abeilles sont échantillonnées au hasard grâce à la méthode décrite par Ritter (RITTER, 1981; RITTER et al., 1984; DE JONG et al., 1982; PAPAS et al., 1986; GARCIA-FERNANDEZ et al., 1995).
Le troisième paramètre ciblé est le taux d'infestation du couvain (TIC); c'est le pourcentage des varroas occupant 100 alvéoles ouvertes et operculées. L'obtention du couvain se fait par prélèvement mensuel et au hasard avec des dimensions de 10 cm sur 10 cm.
Pour le volet statistique, les trois paramètres ciblés sont les variables considérées dans l'étude de l'évolution de la parasitose et mis en évidence pour vérifier s'il y a des différences ou non entre les trois écosystèmes. L'analyse de la variance est faite avec le total de chacun des taux d'infestation, tout en procédant à l'analyse des différentes paires possibles qui peuvent être formées avec ces trois groupes de climats. Pour l'interprétation des résultats obtenus, les données ont fait l'objet d'analyses statistiques. Ces dernières ont été effectuées à l'aide des logiciels Excel, Excel-Stat et SPSS.
Résultats
Les valeurs moyennes de la mortalité de varroa (M), du taux d'infestation des abeilles adultes (TIA) et du taux d'infestation du couvain (TIC) au cours de l'échantillonnage pour l'ensemble des colonies sont regroupées dans le Tableau 1 selon le paramètre climatique auquel elles appartiennent.
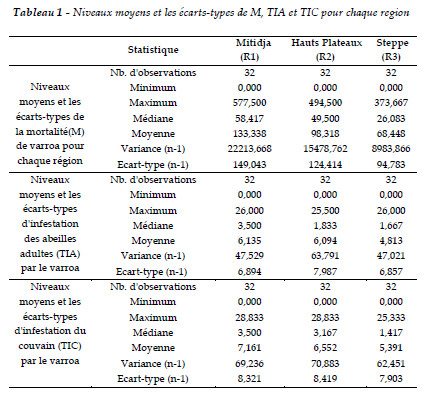
Évolution de la mortalité (M) de Varroa pour les régions d'étude
La comparaison des données de l'évolution des taux de mortalité en 2010, 2011 et 2012, montre que l'impact de M pour les trois régions a atteint une augmentation sensible de février à août avec des seuils au mois de juin de 577 pour la Mitidja (R1), 494 pour les hauts plateaux (R2) et 373 pour la steppe (R3) (Tableau 1 et Figure 2). La confrontation des données de l'évolution de M, entre les régions et entre les trois années, apportent des éclairages additionnels sur les résultats. Ainsi nous notons qu'il y a une diminution de M en allant du nord vers le sud. La mortalité enregistrée en Mitidja est supérieure à celle des hauts plateaux et de la steppe. Nous avons aussi enregistré une régression progressive de cette mortalité pour les trois années.
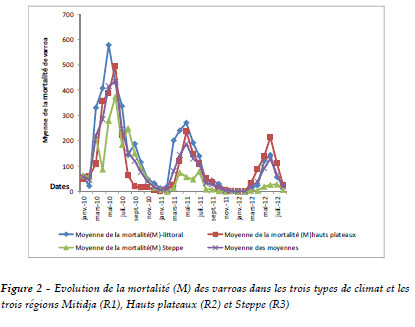
Pour les analyses descriptives de M sous les trois climats, il ressort que les niveaux moyens et les écarts-types de la mortalité de varroa pour chaque région sont consignés dans le Tableau 2. Ce dernier révèle l'évolution des moyennes de M et les écarts-types moyens pour les trois régions. Nous constatons que non seulement ces régions se sont distinguées par leur niveau très élevé de mortalité mais aussi par leurs écarts types. En effet, celui de Mitidja dépasse les autres régions, tandis que celui de la steppe demeure très bas.
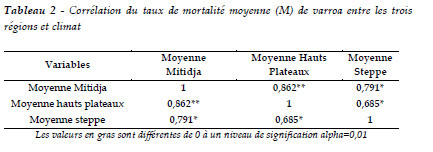
L'analyse de la variance des résultats de la mortalité, révèle des différences hautement significatives pour les trois régions au seuil de 5%. Ces données confirment l'effet de la région et de l'étage bioclimatique sur la mortalité de varroa. C'est en climat méditerranéen qu'on enregistre une mortalité importante de varroa. Il s'est avéré que la mortalité en Mitidja est mieux corrélée à celle du climat semi aride (r = 0.86) qu'à celle de l'aride (r = 0.79). Cette zone désertique est en relation linéaire avec la mortalité des Hauts Plateaux (r =0,68) (Tableau 2). Par conséquent, la mortalité a été retenue comme indicateur pour l'évaluation de la performance des colonies en fin de saison. Ceci indique l'importance de la région et du climat lors de l'évaluation de la mortalité. Dans une autre perspective, la corrélation de Pearson nous apprend aussi que le climat devrait être considéré lors des traitements contre cet ectoparasite.
Évolution des taux d'infestations (TIA) des abeilles
L'évolution de l'infestation des abeilles en 2010, 2011 et 2012 met en évidence l'importance du facteur d'évolution des abeilles durant ces années. Il existe des différences entre les années où nous avons enregistré une importante évolution de mars à août avec un maximum d'infestation de l'ordre de 25,5% pour les hauts plateaux et 16,16% pour la Mitidja et la steppe et une régression pour s'annuler durant la période de septembre à février (Tableau 1 et Figure 2).
L'analyse descriptive des TIA pour les trois régions montre que la moyenne d'infestation des abeilles varie d'une région à une autre. Elle diminue en allant du nord vers le sud. Le Tableau 1 et la Figure 3 montrent une répartition homogène d'infestation des abeilles de la steppe, ainsi que celles de la Mitidja et des hauts plateaux.
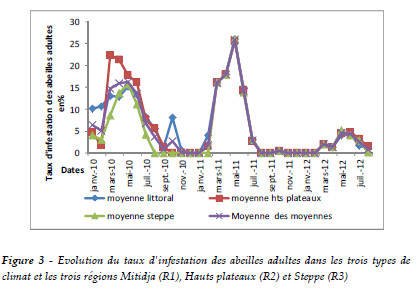
L'effet de la région sur le TIA est confirmé par l'analyse de la variance. Il montre que la différence des TIA des trois régions est hautement significative à 5% de probabilité tel que (Fc1= 9,1, Fth1= 0,005; Fc2= 5,88, Fth2= 0,02, p=0,05 ; Fc3= 2,22, Fth1= 0,14, p=0,05). Ces résultats nous permettent d'affirmer l'influence directe de l'étage bioclimatique sur l'infestation des abeilles.
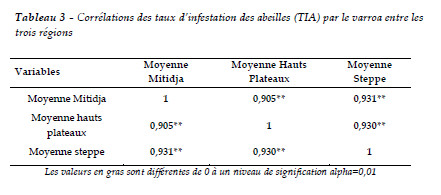
Les corrélations entre les trois régions concernant le TIA indiquent que les colonies les plus fortes, surtout en couvain, présentent des taux d'infestation les plus élevés en fin de saison. Dans la corrélation de Pearson, nous avons remarqué de fortes corrélations positives entre les TIA de la région de Mitidja et des hauts plateaux où r = 0.90 et le TIA de Mitidja avec le TIA de la steppe où r =0,93 et elle est positive et linéaire entre le TIA de la région des hauts plateaux avec le TIA de la région steppe avec r = 0.93 (Tableau 3).Cependant, il semble indiquer que le climat de la région joue un rôle dans la résistance des abeilles aux varroas. Le fait que cette corrélation soit positive laisse supposer que la région et l'étage bioclimatique ont un impact sur l'évolution de l'infestation des abeilles.
Évolution du taux d'infestation de couvain (TIC) pour les trois régions
Le seuil du TIC moyen varie d'une année à une autre. L'infestation de couvain est plus élevée en 2011 qu'en 2010 et elle est moindre en 2012. La lecture du Tableau 1 et de la Figure 4 montre qu'il y a une évolution importante de février à août, pour atteindre un maximum de 28% pour la Mitidja et les hauts plateaux et de 25% pour la steppe. Aucune infestation n'est relevée entre septembre-février. Cela est en relation étroite avec la présence de couvain.
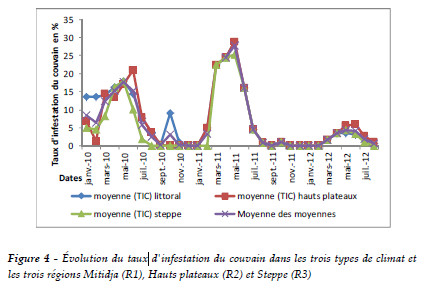
Pour l'effet de la région sur le TIC par le varroa, l'analyse de la variance des résultats obtenus pour les trois régions montre des différences statistiquement significatives au seuil de 5%. Cela confirme l'influence de la région sur les TIC.
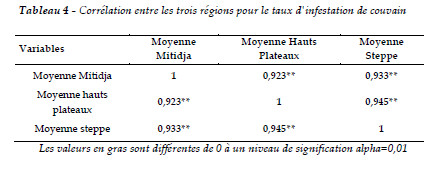
Selon la méthode de Pearson, les taux d'infestation du couvain entre les trois régions sont corrélés positivement. Le TIC de la Mitidja est en relation linéaire avec le TIC des hauts plateaux (r =0,92) et avec le TIC de la steppe avec r = 0,93. La corrélation est également positive entre le TIC des hauts plateaux et le TIC de la steppe où r =0,94. La mise en évidence de cette corrélation confirme l'importance de l'effet du climat sur la résistance et l'infestation de couvain. Certaines colonies montrent en effet un TIC supérieur (cas de la Mitidja). Cet aspect est très important et doit être pris en compte dans toute recherche de traitement et de lutte afin de mieux développer la résistance du couvain à cette varroase (Tableau 4).
Corrélation entre les trois paramètres étudiés (M, TIA et TIC) pour chaque région d'étude
Pour les colonies en Mitidja, l'analyse de la corrélation entre M, TIA et TIC, montre qu'il y a une relation linéaire. Elle est positive et statistiquement significative entre TIA et TIC, où les deux paramètres évoluent de la même façon (r= 0,98, p= 0,01). Positive mais non significative (r= 0,71, p= 0,01); cette relation s'établit entre M et TIA. Elle est aussi positive mais non significative (r= 0,67, p= 0,01); cette relation s'établit entre M et le TIC. Il est possible que des facteurs saisonniers aient eu une certaine influence sur les résultats. Toutefois, le varroa vit et se multiplie dans un environnement protégé et maintenu à température constante par l'activité de thermorégulation des colonies. Pour cela dans un même lieu géographique, les saisons entrainent la fluctuation de façon importante des taux de l'infestation (Tableau 2).
Pour le groupe des hauts plateaux, l'analyse de la corrélation entre M, TIA et TIC, montre qu'il y a une relation linéaire. Elle est positive et statistiquement significative entre le TIC et le TIA, où les deux paramètres évoluent de la même façon (r= 0,93, p= 0,01); une relation positive mais non significative (r= 0,69, p= 0,01) entre le M et le TIA et une corrélation positive mais non significative (r= 0,65, p= 0,01) entre le M et le TIC (Tableau 2).
La relation entre les deux paramètres TIA et TIC est plus forte pour chaque région, ce qui signifie que l'évolution biologique et l'état des abeilles ont un effet sur la relation entre ces deux paramètres.
Pour ceux de la steppe, le Tableau 4 montre également les corrélations entre les trois paramètres étudiés. La corrélation du TIA et du TIC a permis de révéler une relation positive et statistiquement significative (r= 0,97, p= 0,01). La corrélation entre M et TIA est positive et non significative (r = 0,32, p = 0,01) et la corrélation entre M et TIC est positive et non significative (r = 0,25, p = 0,01).
Discussion
Le développement de Varroa est très lié à l'évolution biologique de son hôte. En Algérie, il existe une diversité d'écosystèmes et de climats, qui fait que les cycles biologiques des abeilles diffèrent dune région à une autre. Cela a un effet direct sur le développement du parasite. Varroa a suivi un modèle de croissance bimodal qui se manifeste dans les zones étudiées. Dans le climat aride, la population de Varroa a atteint son seuil de croissance maximale 1 à 2 mois après par rapport à celle de la Mitidja où le développement des colonies est précoce. Ce retard de croissance des acariens pourrait être lié au climat et aux périodes de floraison qui ont un effet direct sur le développement des colonies. Au nord de l'Algérie, surtout en Mitidja, le cycle biologique de Varroa est plus précoce qu'au sud. CALATAYUD et al. (1992) ont étudié la dynamique des populations de Varroa dans un rucher en Espagne et ont observé des maxima d'infestation (TIA) en avril-mai. Quant à GARCIA-FERNANDEZ et al. (1995), ils ont montré que la période dinfestation maximale varie selon la zone climatique. Ces résultats sont identiques aux notres. Dans notre étude, nous avons constaté que les maxima des populations de Varroa se situent en mai et juin, tandis qu'en Allemagne, RITTER et al. (1984) et MORITZ et al. (1990) ont observé les seuils aux mois de juillet et août. Les différences observées dans la dynamique des populations de Varroa destructor pourraient être dues à l'apparition précoce des miellées en Mitidja. Ces floraisons entrainent la précocité du développement des abeilles et par conséquent celle de Varroa. Cela a été mis en évidence par WOYKE (1987) et ORTEGA (1986) où les températures inférieures à 10°C peuvent aussi modifier la période de développement du couvain operculé provoquant ainsi une croissance accrue de la population de l'acarien. La température et la précocité des floraisons peuvent provoquer, dans les zones froides des hauts plateaux et de la steppe, l'apparition des maxima en mars-avril et surtout quand les périodes de gelées ne sont plus fréquentes, ce qui aboutira à l'affaiblissement des colonies d'abeilles. En revanche, en Mitidja, où les températures sont douces en hiver, le développement de l'ectoparasite est constant jusqu'en août, moment qui coïncide avec l'effondrement des colonies. Par ailleurs, les niveaux moyens élevés d'infestation sont observés pour le climat méditerranéen que pour les semi-aride et aride. MORETTO et al., 1991, ont réalisé une étude au Brésil sous 3 types de climats où ils ont constaté que les TIA et TIC sont plus élevés en climat froid, et ils ont conclu que l'évolution de Varroa dépend plus du climat que de la génétique de l'abeille. En climat méditerranéen, nous avons constaté des TIC avec des maxima pendant les mois de mars, avril, mai et juin. Malgré cela, les colonies restent vigoureuses et actives. Le développement des populations de Varroa est accéléré en climat méditerranéen avec une disponibilité mellifère permanente pendant toute l'année. Le climat et la phénologie sont deux facteurs qui peuvent contrôler la dynamique des populations de Varroa. Le constat à faire est de confirmer que des températures supérieures à 37°C, surtout en zones climatiques semi arides et arides allant de mai à la fin d'août, ont un effet d'éradication de l'ectoparasite. Cela nous permet de penser que les populations de Varroa ont tendance à diminuer sensiblement au cours des années. Selon HOULE (2004), des expériences ont été menées sur l'utilisation de la chaleur contre le varroa. Certaines ont abouti avec un certain succès et d'autres pas. Avec la thermothérapie, il s'agit de trouver la température et la durée de traitement qui permettent de réduire le nombre d'acariens sans tuer les abeilles, car les acariens sont très sensibles à la chaleur. Une température supérieure à 36°C met en péril la reproduction du varroa. Ce même auteur note que dans certains pays, comme l'Algérie et le Maroc, les abeilles arrivent à cohabiter avec le varroa. La dynamique de développement de l'acarien est ralentie par la température du couvain restant supérieure à 36°C. Des expériences, réalisées en Louisiane aux USA ont démontré qu'une température de 39°C pendant 48 heures décimait les populations de Varroa (ANONYME, 1993). À 44°C pendant quatre heures, les varroas présents dans le couvain meurent mais pas nécessairement ceux qui sont sur les abeilles. Ces dernières réussissent à garder la température plus basse. Afin d'élever la température dans les ruches, les chercheurs les ont simplement peintes d'une couleur foncée plutôt qu'en blanc. Dans une ruche foncée, les abeilles passent plus de temps à bourdonner pour baisser la température de l'air, ce qui fait qu'elles s'échauffent elles-mêmes. Comme les acariens sont très sensibles à cette élévation de température, ils en tombent sur le fond de ruche et meurent. Ce sont des techniques qui nécessitent un appareillage spécial et peuvent avoir un effet négatif sur le développement de la colonie si cette dernière est surchauffée. Des recherches ont aussi démontré que des ruches placées à l'ombre avaient plus de varroas par rapport à celles exposées au soleil (CHARRIÈRE et al., 2011).
Conclusion
En apiculture les stratégies de thérapie chimiques et médicamenteuses pour léradication des maladies montrent de l'essoufflement et comportent aussi plusieurs inconvénients, comme les coûts élevés et les possibilités de contamination des produits de la ruche destinés à la consommation humaine. Ils présentent aussi des risques pour la santé de l'éleveur.
Pour l'apiculture algérienne, il faut de toute urgence accélérer le développement d'une stratégie de lutte intégrée contre la varroase et cela par la recherche de méthodes alternatives de contrôle pouvant éventuellement être combinées. Il faudrait mettre en place une stratégie de lutte intégrée la plus durable possible, la plus économique et qui demanderait un minimum d'effort de la part de l'apiculteur. Idéalement elle éliminerait aussi les risques de contamination des produits de la ruche.
Dans cette situation, le cheptel apicole algérien bénéficie de plusieurs atouts qui le prémunissent contre cette parasitose à savoir la rusticité de l'insecte et un milieu hostile au développement de cet ectoparasite Varroa destructor.
Cette étude confirme l'effet de la région et de l'étage bioclimatique sur la mortalité de varroa. C'est en climat méditerranéen qu'on enregistre une mortalité importante de varroa. Cela nous permet de penser que les populations de Varroa ont tendance à diminuer sensiblement au cours des années. Dans le cas des climats semi-arides et arides aux étés chauds, nous avons constaté que les températures élevées ont un effet néfaste sur les acariens. Cela a ralenti l'effondrement des colonies comparativement à celles logées sous des climats septentrionaux; cela apporte une preuve tangible quant à la grande résistance de l'abeille Apis mellifera intermissa par rapport aux autres races vivant dans des régions septentrionales.
Références bibliographiques
BERKANI, M.L., GHALEM-BERKANI, Z., 2009.Composition of chemical and acaricide activity of 4 essential oils resulting from plants against "Varroa destructor" in the Algerian Center. Actes du 41econgrès Apimondia 15 au 20 septembre 2009, Montpellier-France. [ Links ]
CALATAYUD, F., VERDU, MJ., 1992. Evolucion anual de parametros poblacionales de colonias de Apis Mellifera L. (Hymenoptera: Apidae), parasitadas por Varroa jacobsoni Oud (Mesostigmata : Varroidae). Bol. Sani. Veget. Plag. 18: 775-789. [ Links ]
CHARRIÈRE, J.D., DIETEMANN, V., SCHÄFER, M., DAINAT B., NEUMANN, P., GALLMANN, P., 2011. LeitfadenBienengesundheit des ZentrumsfürBienenforschung. ALP forum. 84: 1-35. [ Links ]
DEFAVEAUX, A., 1981. La varroatose dans les ruchers de l'est Algérien. 1981. Rev. Rech.CURER. 7: 1-15 [ Links ]
DE JONG, D., ROGER, A., MORSE, G., EICKWORT, C., 1982b. Mite pest of honey bees. Annu. Rev. Entomol. 27: 225-252. [ Links ]
GARCIA-FERNANDEZ, P., BENITEZ-RODRIGUEZ, R., ORENTES-BERMEJO, F.J., 1995. Influence of climate on the evolution of the population dynamics of the Varroa mite on honeybees in the south of Spain. Apidologie 5:361-440. [ Links ]
HOULE, E., 2004. Les méthodes physiques en lutte intégrée. Centre de Référence en agriculture et agroalimentaire. Québec, Canada, 3-4. [ Links ]
MORETTO, G., GONÇALVES, L.S, DE JONG, D., BICHUETTE, M.T., 1991. The effects of climate and bee race on V jacobsoni Oud infections in Brasil. Apidologie 22: 197-203. [ Links ]
MORITZ, R.F.A., MAUTZ, D., 1990. Development of Varroa jacobsoni in colonies of Apis mellifera capensis and Apis mellifera carnica. Apidologie 21: 53-58. [ Links ]
ORTEGA, J.L., 1986. Flora de interés apicola y polinizacion de cultivos. Mundi-Prensa, 149 pp. [ Links ]
PAPAS, N., THRASYVOULOU, A., 1986. Searching for an accurate method to evaluate the degree of Varroa infestation in honeybees colonies. Proceedings of meeting of the EC-Experts Group, European Research on Varroosis Control, Bad Homburg, Oct 1986 (R Cavarollo, ed), 85-86, Comiss Europ Commun Joint Res Centre, I-21020 Ispra. [ Links ]
RITTER, W., 1981. Varroa disease of the honeybees. Apis mellifera. Bee World 62: 141-153. [ Links ]
RITTER, W., DE JONG, D., 1984. Reproduction of V. jacobsoni Oud in Europe, the Middle East and tropical South America. Z .Angew. Entomol. 98: 55-57. [ Links ]
SELTZER, P., 1946. Travaux de l'Institut de météorologie et de physique du globe de l'Algérie. Institut de météorologie et de physique du globe de l'Algérie, édit Carbonel., 219 pp. [ Links ]
USDA, 1993. Dark-colored hives help protect bees. Quarterly Report of Selected Research Projects, juillet-septembre 1993: 11
WOYKE, J., 1987. Comparative population's dynamics of Tropilaelaps clareae and Varroa jacobsoni mites on honeybees. J. Apic. Res. 26: 196-202. [ Links ]
Entregue para publicação em abril de 2013
Aceite para publicação em outubro de 2013
