










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias v.34 n.1 Lisboa jan./jun. 2011
Organic C distribution and N mineralization in soil of oak woodlands with improved pastures
Maria Xesús Gómez-Rey1, Ana Garcês1 and Manuel Madeira1
1Centro de Estudos Florestais, Instituto Superior de Agronomia (Universidade Técnica de Lisboa), Tapada da Ajuda, 1349-017 Lisboa, Portugal, Tel: +351213653264, Fax:+351213635031, E-mail: mxgomez@isa.utl.pt
ABSTRACT
Cork oak woodlands in Portugal are a multipurpose agroforestry system occurring in areas mostly degraded by former cereal crops and overgrazing. The objective of this study was to evaluate the effect of scattered cork oak trees (Quercus suber L.) on soil quality, considering two land use systems: unmanaged pastures and improved pastures. Soil samples were collected in a square grid around scattered cork oak trees to evaluate the spatial variability of soil bulk density and chemical properties. In soil samples taken beneath tree canopy and in the open, net N mineralization was evaluated by aerobic incubations. Also, laboratory incubations were carried out to evaluate the effect of roots (tree or/and herbaceous) and water solutions (bulk precipitation, throughfall or stemflow) on net N mineralization.Contents of organic C, total N and extractable P were increased beneath the tree canopy, and gradually decreased with the increase of distance to the tree trunk. Improved pastures established 26 years ago increased organic C (0.76 kg m-2), total N (0.06 kg m-2) and extractable P (2.70 g m-2) amounts in the 0-10 cm soil layer beneath the tree canopy relatively to those of unmanaged pastures. Net N mineralization was significantly greater (about 2 times) in soils beneath the tree canopy than in those of open areas. N mineralization was unaffected by addition of throughfall or stemflow, while a significant reduction resulted from tree roots addition, this effect being stronger for herbaceous plant roots than for tree roots. Maintenance of tree cover combined with permanent improved pastures should be considered to improve soil quality in oak woodlands of Southern Portugal.
Keywords: Agroforestry systems, legumes, Mediterranean climate, soil quality.
Distribuição espacial do C orgânico e da mineralização de N em solos de montado com pastagem melhorada
RESUMO
O montado em Portugal corresponde a um sistema multifuncional que ocorre em áreas frequentemente degradadas pelo cultivo de cereais e de sobre pastoreio. O objectivo deste estudo foi avaliar a influência das árvores espaçadas na variação do C orgânico, N total e P extraível num montado (Quercus suber L.) com pastagem melhorada (instalada há 26 anos) e sujeito a gestão intensiva e noutro com a vegetação herbácea natural. Amostras de solos foram colhidas numa quadrícula em torno de sobreiros espaçados, sob a respectiva copa e fora da influência da mesma, para avaliar a variabilidade espacial da massa volúmica e propriedades químicas do solo. Estudou-se a mineralização líquida de N através de incubações aeróbias. Também se efectuaram incubações para avaliar o efeito da precipitação interna (gotejo e escorrimento pelo tronco) e das raízes das herbáceas e das árvores na mineralização líquida de N. Os teores de C, N e P foram superiores no solo da área sob a projecção vertical da copa da árvore, observando-se o seu decréscimo com o aumento da distância ao tronco da mesma. A instalação de pastagem melhorada determinou um aumento das quantidades de C orgânico (0.76 kg m-2), N total (0.06 kg m-2) e P extraível (2.70 g m-2) sob a árvore (0-10 cm) comparativamente à área com vegetação herbácea natural. A mineralização líquida de N foi significativamente superior (cerca de 2 vezes) na área sob a copa da árvore do que naquela fora da influência desta. A adição de gotejo não influenciou a mineralização líquida do N, enquanto as raízes, tanto das árvores como sobretudo da vegetação herbácea, contribuiram para o seu decréscimo. Com vista a melhorar a qualidade do solo e o sequestro de carbono, deverá ser equacionada a manutenção das árvores associada a sistemas de gestão adequados à sustentabilidade do ecossistema, que poderão passar pela instalação de pastagens melhoradas.
Palavras-chave: Clima mediterrâneo, leguminosas, mineralização de N, sistemas agroflorestais.
INTRODUCTION
In Portugal, cork oak and holm oak woodlands mostly occur in dry sub-humid to semi-arid areas and they occupy about 11x105 ha (IFN, 2006). Most oak woodlands occur in an agroforestry system which is characterised by the combination of an open tree cover with an understorey of grassland, cereal crops or Mediterranean shrubs. In most of the oak woodlands occur natural pastures, which have been extensively used for livestock rearing, with minimum management and productivity. Improved sown permanent pastures (that is, mixture of legumes and grasses) have been installed at low extent, aiming mostly to increase the stocking rate.
Tree cover is scattered, with a tree density generally lower than 80 trees per hectare (DGF, 2001), and these areas are characterised by a savannah-like physiognomy (Joffre et al., 1999). The long-term sustainability of these ecosystems may be further threatened by the regional effects of global warming, which are predicted to result in increases in the length, severity and frequency of summer droughts in Mediterranean regions (Miranda et al., 2002). In similar systems, scattered trees lead to a great spatial variation of soil conditions (creating islands of greater fertility), enhancing soil organic matter and nutrient pools beneath their canopy (Dahlgren et al., 1997; Young, 1997; Gallardo, 2000) and improving soil structure (Joffre and Rambal, 1988). Such favourable conditions may also increase the biological activity in the soil and improve N mineralization and availability (Belsky et al., 1989).
Until the seventies of the last century, the ground cover rotation of evergreen oak woodlands was composed mostly of cereal crops, fallow and natural pastures for livestock grazing, which produced soils of low fertility, compact and prone to erosion. Nowadays, most of the woodlands areas are covered by natural pastures extensively used for livestock grazing. However, in certain cases overgrazing is causing loss of pasture cover, increase of soil compaction (Coelho et al., 2004) and decrease of tree natural regeneration, thus threatening their fragile sustainability. Therefore, establishment of management systems that will improve soil quality and contribute to long-term sustainability of this agroforestry system is a crucial issue. Installation of improved pastures, with high proportion of legumes, may improve soil properties and increase stocking rate in few oak woodlands. These pastures may improve soil fertility and productivity, since they enhance organic matter and N contents in the soil (Haynes and Williams 1993; Rao et al.,1994). However, this enhancement of soil N status may cause soil acidification through excessive N mineralization and nitrate leaching during the rainy season (Bolan et al., 1991; Haynes 1983). Nevertheless, these effects may be reduced by the input of organic residues with high C/N ratio (such as tree root litter) which may affect soil N dynamics(Corbeels et al., 2003). In addition, N dynamics may be also affected by throughfall and stemflow, richer in soluble organic compounds (Edmonds et al., 1991)
In this context, a study was carried out hypothesising that scattering of oak trees could determine differences on the soil C accumulation and N dynamics and availability. For this, a study was developed in a cork oak woodland subjected to two different land use (unmanaged pasture and long-term managed improved pastures) to (i) assess the effect of scattered trees on spatial variability of soil bulk density and soil organic C, total N and extractable P contents, and (ii) evaluate the potential effects of fine roots (from trees and/or herbaceous) and tree water solutions (throughfall and stemflow) on soil net N mineralization.
MATERIAL AND METHODS
Site description
The present study was carried out in the "Herdade dos Esquerdos" located close to Monforte (Alentejo region), Southern Portugal (lat. 39º 07'- 39º 08' N; long. 7º 29'-7º 30' W; 314 m a.s.l.). The area has a Mediterranean mesothermic dry sub-humid climate, characterized by hot and dry summers. The mean annual temperature is 15 ºC and the mean annual rainfall is 600-650 mm (INMG, 1991). The area is characterized by gently undulated topography and predominant slope gradient is 5-8%. Soils are mostly Haplic Leptosols (Dystric) and Epileptic Cambisols (Dystric) (IUSS Working Group, 2006) developed over gneiss, with sandy loam texture, low organic matter content and P status and acidic.
The vegetation of the study area consists in mature Quercus suber woodland (30-40 trees ha-1, average cover canopy of 35 %) which was subjected to a rotation of cereal crops, grazing (natural pastures) and fallow until the 1970s. After abandonment of cereals crops, the study area partially became unmanaged woodland, with a natural cover of a mixed understory of herbaceous plants and spontaneous shrub patches, dominated by Cistus spp. This systemhas been extensively used for sheep grazing, with a stocking rate of about 0.9 sheep ha-1 yr-1.
Improved sown permanent pastures (Trifolium spp., Ornithopus spp., Lolium spp. as dominant species)have been progressively installed in the study area since 1973. They have been subjected to intensive grazing by sheeps (stocking rate between 5-8 sheep ha-1 yr-1 was achieved)and fertilized every year with 200 kg ha-1 of superphosphate 18 % P2O5 until 2003. Since then this application has been replaced by 300 kg ha-1 of natural rock phosphate ("fertigafsa": 26.5 % of P2O5, 35% of CaO, 3.2% of SO3 and 0.8% of MgO). Due to the high stocking rate shrub regeneration has been impeded.
Sampling
Sampling took place in two adjacent management system areas with similar geology, soil type and texture (sandy loam), and terrain morphology and slope: unmanaged cork oak woodland with natural pastures (UN), and managed cork woodland where improved pastures were installed 26 (M26) years ago. We assumed that the two areas were initially comparable in terms of soil fertility, although it was not possible to confirm this assumption. In each management system area, four circular plots (1256 m2), the common plot design used in Portugal for oak woodlands inventory studies (DGF, 2001), were delimited and located at least 200 m apart in order to ensure randomness and independency (Lima et al., 2006). In each plot (which included at least 5-6 trees) a representative cork oak tree (similar crown diameter and circumference at breast height) was selected for sampling purposes.
The effect of tree canopy on spatial variability regarding amounts of total organic C, particulate organic matter fraction, total N and extractable P content were evaluated in both management system areas. A square grid (2m x 2m) was placed in the surrounding of each selected tree in a circular area covering 13 m of radius, corresponding to about 200% of the crown radius, following the methodology described by Gallardoet al. (2000).Soil samples were collected in this area, at intervals of 2 m. In each circular plot, a total of 72 samples were taken for both 0-10 and 10-20 cm depth: 36 beneath the tree canopy (BC) and 36 in the open (OP). In each management system area, the four samples localized in the same position of each plot were combined in one composite sample. To assess the variation of organic C and N amounts with the distance to tree trunk, 40 composite samples collected at ten distances and four orientations (2, 4, 6, 8, 10 and 12 m along the N, E, S and W axis; and 2.8, 5.7, 8.5, 11.3 m along the NE, SE, SW and NW axis) were selected per depth and management system area. Samples were air-dried and sieved (< 2 mm) for the determination of chemical properties (organic C, particulate organic matter C, total N, and extractable P).
Soil bulk density was determined on undisturbed samples using the core method (Blake and Hartge, 1986). Soil column cylinders were taken (0-10 and 10-20 cm depth) beneath the tree crown (2, 4, 6 m) and in the open (8, 10 and 12 m) along the four main orientations (N, S, W, E), cut and weighed to determine the soil dry weight. Therefore, around each selected tree in each sampling plot, 12 samples by depth were taken both beneath tree crown and in the open.
The mineral soil, fine roots (cork oak tree and herbaceous species) and water solutions (bulk precipitation, throughfall and stemflow) to be used in aerobic incubations were collected in the unmanaged woodland area in May 2002, when plants showed higher quantity of fine roots. In each plot, soil cores (0-5 cm and 5-15, 65 mm diameter) were collected beneath the tree canopy (50% of radius canopy) and in the open area (150% of radius canopy) along the four main orientations (N, E, S and W), totalling four samples for each depth, which were combined to give one composite sample. The field-moist soil was sieved (< 4.75 mm) and stored at 4 ºC for less than three days until to prior incubation procedures. Sub-samples were sieved (< 2 mm) and taken for analysis. The soil showed low C/N ratio (11.7) and total and mineral N (NH4+ + NO3-) contents were 1.0 g kg-1and 8.2 mg kg-1, respectively, being the N-NH4+ the predominant form of mineral N (62%).
For tree roots collection, a pit was open (20 cm depth) close to the trunk of one selected cork oak and fine roots were collected by hand. Herbaceous plants [Geranium molle L., Ornithopus compressus L., Ornithopus pinnatus (Miller) Druce and Rumex bucephalophorus L., Poa spp.] were pulled out from the soil, and the first 5 cm of roots were collected, washed with deionized water, dried (40 ºC) and cut into sections of 1 cm. A sub-sample was taken for analysis. Samples of bulk precipitation, throughfall and stemflow were collected in winter, during a rainy period, and samples were stored at –15 ºC. A sub-sample was used for chemical analysis (Table 1). Nutrients amounts in stemflow, and to a lesser extent in throughfall, were greater than in the bulk precipitation.
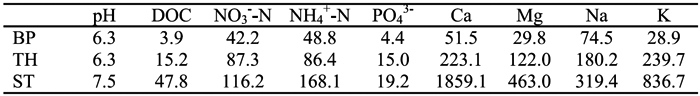
Table 1 - Values of pH and concentrations of dissolved organic C (DOC, mg L-1), cations and anions (µmolcL-1) in the bulk precipitation (BP), throughfall (TH) and stemflow (ST) solutions used in the aerobic incubations.
Laboratory incubation treatments
In order to evaluate tree influence on soil net N mineralization, sub-samples (100 g fresh weight) of the 0-5 and 5-15 cm soil layers from the open area were placed in plastic containers (7.5 cm diameter, 5 cm height) and subjected to seven different treatments, each treatment replicated six times. The treatments were applied in two experiments.
In the first, the effect of throughfall solution (TH) and stemflow solution (ST) on net N mineralization, regarding the bulk precipitation solution (BP), was assessed. In each treatment 20 g mL of solutions were added to each soil subsample, 60% of water holding capacity was achieved.
In the other experiment the effect of tree and herbaceous plant roots on net N mineralization was assessed. The treatments were: (C) mineral soil without roots addition, (H) addition of 1 g of herbaceous roots, (T) addition of 1 g of oak tree roots, (HT) addition of both 0.5 g of herbaceous root and 0.5 g of tree roots. To minimize the effect of small quantity of soil substrate used in the incubation and to stress differences between treatments, the amount of root residues added was higher than that founded in the field. The quantity of water applied (20 mL) corresponded to 60% of soil water holding capacity.
Laboratory procedures
The particulate organic matter fraction was separated by a 50 mm sieve after dispersing air-dried soil by shaking with K2SO4 (10 g / 30 ml) for one hour. The organic C in both the bulk samples (Corg) and the particulate organic matter fraction (CPOM) was determined by wet oxidation (De Leenheer and Van Hove, 1958). Non-particulate organic fraction was calculated as the difference between CPOM and organic C. Total N was determined using Kjeldhal digestion (Digestion System 40, Kjeltec Auto 1030 Analyzer). Extractable P was extracted using the Egnér-Riehm method (Egnér et al., 1960) and determined by the molybdate-blue method.
Aerobic laboratory incubation, in the dark, was carried out during 8 weeks and net N mineralization was evaluated periodically. Soil samples (100 g fresh weight) were incubated in plastic containers (7.5 cm diameter, 5 cm height), at 30 ºC and 60% of water holding capacity. Plastic containers were covered with parafilm to allow gas exchange and to minimize evaporation initial soil moisture content being weekly adjusted by adding distilled water. Initially, and every two weeks, NO3--N and NH4+-N were extracted by shaking 5 g of soil (soil: solution ratio 1:10) for one hour in 2 M KCl and extracts were filtered. Contents of NO3--N and NH4+-N in extracts were determined by a segmented flow autoanalyzer (Skalar, SANplus System, Breda), using the hydrazinium reduction and the modified Berthelot method, respectively (Houba et al., 1994). Soil moisture was measured by drying a subsample at 105 ºC to constant weight. Net N mineralization was calculated by difference between NO3--N and NH4+-N at each incubation time and their respective initial levels.
Statistical analysis
Differences in soil bulk density and C, N and P contents, as dependent variables, between position (beneath canopy and open area), management (unmanaged and improved pasture) and depth (0-10 and 10-20 cm), as independent variables, were evaluated by three-way ANOVAs. The effect of treatments on cumulative net N mineralization was tested by one-way ANOVA. Differences between treatments were tested using the Tukey multiple range test. Statistical procedures were performed using Statgraphics plus 5.1 (Manugistics Inc.).
RESULTS
Bulk density and C, N and P contents
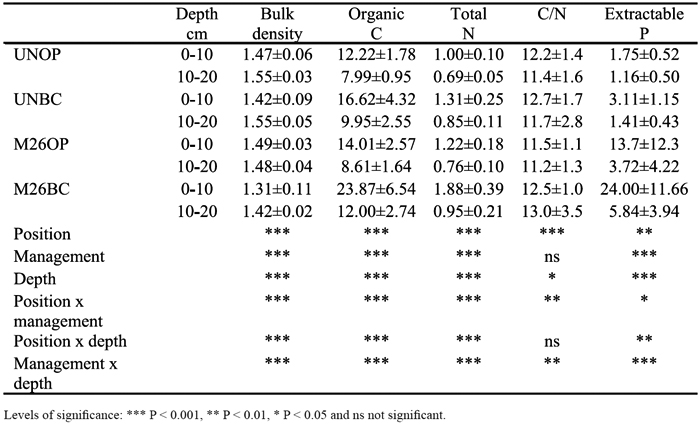
Soil bulk density was significantly lower beneath the tree canopy than in the open (Table 2). The position x management interaction showed that differences between beneath trees and the open were stronger in improved pastures. Bulk density in the 0-10 cm soil layer was of the same magnitude in unmanaged and improved pastures, while in the 10-20 cm soil layer values were higher in unmanaged than in managed pastures (interaction management x depth).
Table 2 - Soil bulk density (g cm-3) and contents of organic C, total N (g kg-1) and extractable P (mg kg-1) in the 0-10 and 10-20 cm soil layers beneath the tree canopy (BC) and in open (OP), in unmanaged (UN) and improved pastured plot established 26 years ago (M26). Values are mean ± standard deviation (n=12 for bulk density, and 36 for other parameters).
In the 0-10 cm soil layer, amounts of organic C and total N decreased with the increase of the distance to the tree trunk, in both unmanaged and 26 years old improved pastures (Figure 1), but the gradient was greater in the improved than in the unmanaged pasture. The gradient in the 10-20 cm soil layer was not affected by pasture management, and was smaller than in the 0-10 cm soil layer.
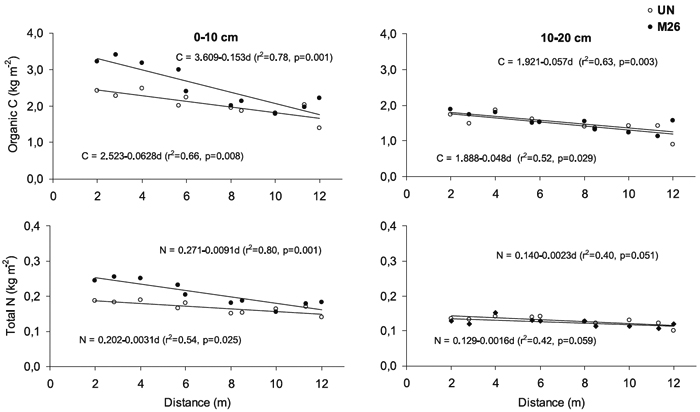
Figure 1 - Variation of organic C and total N amount (kg m-2) with increasing distance from the tree trunk (m) in the 0-10 and 10-20 cm soil layers in the unmanaged (UN) and managed plot with improved pasture established 26 years ago (M26). (C: amount of organic C, N: amount of N, d: distance to the tree trunk). Values are mean (n=4).
As observed for the gradient regarding the distance to tree trunk, mean contents of organic C and total N were significantly greater beneath canopy than in open areas (Table 2). This increase was more pronounced in improved (1.6 and 1.4 times for organic C and N, respectively) than in unmanaged pastures (1.3 and 1.2 times for C and N, respectively; interaction position x management). Irrespectively to the position, a significant increase was also observed in improved pastures relatively to unmanaged pastures. Contents significantly decreased with soil depth (Table 2). Differences between positions and managements were attenuated with depth (interaction position x depth and management x depth).
In improved pastures, values of C/N ratio were higher beneath tree canopy than in open, while in unmanaged pastures (interaction position x management) the values were of the same magnitude. The position x management interaction also showed that C/N ratio in open areas of unmanaged and improved pastures was similar.
Organic C mostly corresponded to the non-particulate fraction (58-79%, Figure 2). The increase of particulate fraction in soils sampled beneath the tree canopy relative to those in the open (2.2-3.0 times) was higher than that observed for non-particulate fraction (1.4-2.1 times). The percentage of organic C in the particulate fraction decreased with soil depth.
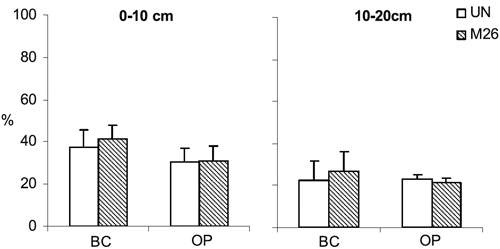
Figure 2 - Percentage of particulate organic matter C (CPOM) in relation to total organicC in the 0-10 and 10-20 cm soil layers beneath the tree canopy (BC) and in open (OP), in unmanaged (UN) and improved pasture established 26 years ago (M26). The bars represent the standard deviation of the mean (n=36).
As observed for organic C and total N, contents of extractable P were significantly greater beneath tree canopy than in the open, decreasing with soil depth (Table 2). Improved pasture establishment led to higher extractable P contents, differences being more pronounced (eight times higher) in the 0-10 cm soil layer (interaction management x depth).
Soil N mineralization
Net ammonification was not observed in soil samples during the incubation period (data not shown). At the end of the incubation, soils collected beneath the tree canopy showed higher cumulative net N mineralization (2.5 and 1.8 times, for the 0-5 and 5-15 cm soil layers, respectively) than those taken in open areas (Figure 3). Also, rates of mineralized N per unit of initial total N were significantly higher in soils beneath the tree canopy (62.98 and 17.98 mg N g-1 total N, in the 0-5 and 5-15 cm soil layers, respectively) than in the open (44.65 and 9.73 mg N g-1 total N, respectively).
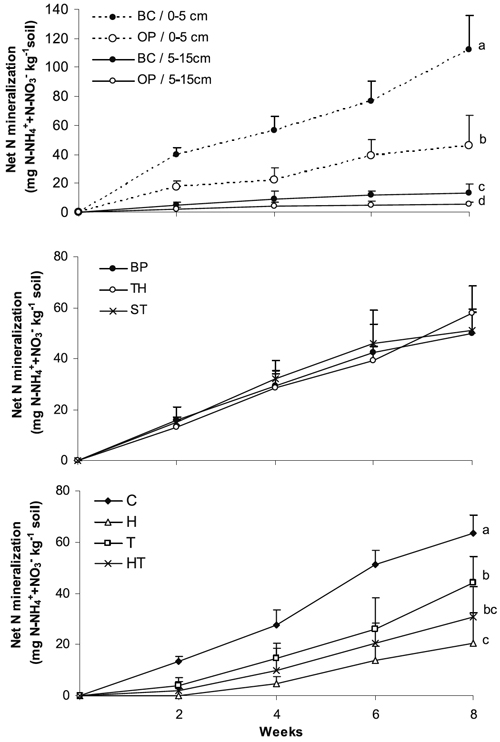
Figure 3 - Cumulative net N mineralization (aerobic incubation) in the 0-5 and 5-15 cm soil layers collected beneath the tree canopy (BC) and in the open (OP) from unmanaged oak woodland; and in the 0-5 cm soil layer (of the open area) incubated with water solutions (BP: bulk precipitation, TH: throughfall, ST: stemflow) or with roots (C: absence of roots; H: herbaceous, T: tree, HT: herbaceous and tree). Different letters for data at the end of the incubation indicate significant differences among means (n=6) at the P = 0.05 level.
Effect of solutions and roots on N mineralization
Net N mineralization was not significantly affected by the addition of throughfall or stemflow solutions, values being similar to those observed for bulk precipitation addition (Figure 3). In this incubation net ammonification was not also observed.
After 8 weeks of incubation, soil cumulative net N mineralization was significantly lower in treatments with roots (20.5-44.2 mg kg-1) than in the control (63.6 mg kg-1). The treatment with only herbaceous roots (H) showed significantly lower cumulative net mineralization (20.5 mg kg-1) than that observed for tree roots addition (44.2 mg kg-1); intermediate values were observed for the HT treatment (mixture of tree and herbaceous species roots).
DISCUSSION
Spatial organic C distribution
Scattered trees induced a strong variation on soil organic C and total N amounts, which gradually decreased with the increase of distance to the tree trunk, a tendency which was also observed in other scattered oak woodlands in Portugal (Nunes, 2005), Spain (Moreno et al., 2007) and California (Dahlgren et al., 1997) under similar climate conditions. As the organic C content variation beneath the tree canopy was mostly observed in the surface soil layer (0-10 cm), it may be mostly associated with the spatial variation of litterfall amounts which has been observed for similar scattered oak trees (Escudero et al., 1985). The spatial variation of organic C content may also be affected by the gradual increase of tree canopy size, as reported by Belsky et al. (1989) for semi-arid savannah; also, the influence of root pattern distribution should be also taken into account. In addition, the stronger variation gradient of soil organic C amount beneath the tree canopy with improved pastures areas (see Figure 1) suggests that this gradient may be also influenced by the interaction between the oak trees and the improved pastures. The similar distribution pattern observed for C and N amounts may be associated with the fact that the latter is cycled via organic matter, as reported by Gallardo (2003) for similar systems.
The amount of organic C in the 0-10 cm soil layer significantly increased beneath the tree canopy in both unmanaged (2.36 kg C m-2) and improved (3.13 kg C m-2) pastures (see Table 1) as compared with the open (1.80 and 2.09 kg C m-2,respectively). The increment of C observed beneath the tree canopy may be mostly ascribed to the additional input of organic matter by litterfall, which can reach 0.36-0.61 kg m-2 yr-1 in Spanish oak woodlands with unmanaged pastures (Escudero et al., 1985; Caritat et al., 1996). This trend is in agreement with the fact that holm oak root length density up to 10 cm depth can be similar beneath and beyond the tree crown (Moreno et al., 2005). The increase in amount of particulate organic matter fraction under oak tree crown was stronger than that observed for total organic C, suggesting that the scattered oak tree effect on soil organic matter is sensitively expressed by the labile fraction of C, such as reported by Lehmann et al. (2001) for single trees in the Brazilian Amazon. Tree litter fall residues, which are in general more recalcitrant to decomposition than those of herbaceous vegetation (Sá, 2001; Sá et al., 2004), may have contributed to the relative increment of soil particulate organic matter fraction. Also, the higher amounts of N and P observed beneath the tree canopy (0.04 and 0.07 kg N m-2, 0.18 and 1.11 g P m-2,in the 0-10 cm soil layer of UN and M26 plots, respectively) may be also explained by litterfall inputs.
In unmanaged pastures, similar C/N ratios (11.4-12.7) were observed beneath the tree canopy and in the open, which is in agreement with results reported for oak woodlands systems with unmanaged pastures of southwest Spain (Gallardo et al., 2000) and California (Dahlgren et al., 1997). Moreover, C/N ratios were also similar in open areas of both unmanaged and improved pastures (11.2-12.2). This is in conformity with the similar increment of organic C and N beneath the tree canopy in both unmanaged and managed pastures (Table 1). As greater C/N ratios were observed in senesced Q. suber leaf fall (59) than in unmanaged (46) and improved (42) pasture residues (Sá, 2001), a proportional higher N accumulation may have taken place beneath tree canopy in the study site, such as suggested by Lehmann et al. (1998) in a decomposition study where tree biomass with the slowest decomposition was the most efficient in increasing the N content in a savannah soil. Also, animal resting under the shade of trees may transfer nutrients from adjacent pastures and contribute to higher proportional N beneath the tree canopy.
The present study shows that the positive influence of oak trees on soil quality was enhanced by improved pastures. Indeed, after 26 years, improved pastures establishment increased organic C (0.76 kg m-2), total N (0.06 kg m-2) and extractable P (2.70 g m-2) amounts in the 0-10 cm soil layer beneath canopy relatively to those of unmanaged pasture. Such as reported by other authors (Haynes and Williams, 1993; Guggenberger et al., 1995; Milne and Haynes 2004), this increase may be partially attributed to greater above and belowground dry matter production in improved pastures. In fact, in the study area, beneath the tree canopy the aboveground biomass of improved pastures (0.45 kg m-2 yr-1)was 1.6 times greater than that of unmanaged pastures (0.28 kg m-2 yr-1; Nuno Cortez, personal communication); however, data on belowground pasture biomass are neededtoconfirm such trend.Moreover, other factors might be involved, such as dung deposition due to greater stocking rate, or a possible enhancement of oak litterfall associated with higher N availability.
Tree influence on N mineralization
The greater net N mineralization observed in soils beneath the tree canopy is in agreement with results reported for Mediterranean oak ecosystems of Spain (Gallardo et al., 2000) and California (Jackson et al., 1990; Herman et al., 2000; Perakis et al., 2007). Although less pronounced, an increment for N mineralization per unit of N (that is, for N turnover rate) was also observed for soils beneath tree canopy. This suggests that differences observed for net N mineralization were not only associated with N accumulation beneath trees. In fact, other factors, such as more favourable soil environmental conditions beneath tree canopies (Joffre and Rambal, 1993; Herman et al., 2000), differences in substrate quality (Matson and Vitousek, 1981) or quantity of ammonifying and nitrifying microrganisms (Charley and West, 1977), may also account for the higher N availability in soils beneath tree canopy.
Results of laboratory incubations suggest that surface soils beneath canopy may have higher potential for NO3--N leaching. However, under field conditions, the deep root system of oak trees, according to Moreno et al. (2005), may contribute to decrease these losses by nitrate uptake from deep soil layers. In addition, input of oak residues with high C/N ratio might promote the immobilization of N during their decomposition as reported by Sá (2001). Our results also suggest that decomposing tree and herbaceous plant roots may have a strong influence on soil N dynamics. In fact, soil N net mineralization was significantly greater in the absence than that in the presence of oak roots, suggesting that decomposing roots in oak woodlands may promote the retention or immobilization of N through their decomposition.
The stronger reduction of soil net N mineralization induced by herbaceous plant roots may be explained by their lower C/N ratio (41), when compared with tree roots (about 60). Herbaceous roots may have decomposed quickly and immobilized N at a higher rate at a short-term. This is in a good agreement with results reported by Sá et al. (2004) for decomposition of similar herbaceous plants in the field, which lost about 25% of their initial mass during the first 40 days of incubation. This pattern suggests that roots of herbaceous species in the open area may strongly contribute to reduce N leaching following the rain events in autumn, when environmental soil conditions are favorable for N mineralization. Such effect will be more pronounced, and over a longer period, beneath the tree canopy, where the combined effect of herbaceous and tree roots occurs. This factor, associated with precipitation reduction observed beneath the tree canopy (David, 2000), may contribute to reduce N losses due to leaching, resulting in an improvement of soil N status in the ecosystem.
In contrast to root addition, soil net N mineralization was not significantly influenced by the addition of throughfall and stemflow solutions, where soluble C and nutrient concentrations were within the ranges previously reported for such solutions in cork oak woodlands in Portugal (Nunes, 2004). The lack of net N mineralization response to addition of these solutions, may be associated with the fact that their organic C and nutrient contents were insufficient to stimulate mineralization dynamics. Nevertheless, this fact should be also related to the relatively high soil biological activity, which could hinder the effect of increased soluble organic C and nutrient contents in the aforementioned solutions. Thus, further studies are needed to get sound information on the subject.
CONCLUSIONS
Our results show that trees in oak woodlands have a positive impact on soil C accumulation and fertility, resulting in a useful process to enhance soil C sequestration and to face desertification in Mediterranean areas. Such effect is clearly intensified by improved pastures installation. Oak trees also enhance net N mineralization and turnover, but simultaneously may decrease N leaching potential by root uptake and root decomposition dynamics. Although the decomposition of herbaceous plants may also have a stronger influence on the decrease of N leaching, further studies are necessary to confirm this trend. Thus, to reverse the degradation which has affected soils in southern Portugal the maintenance of tree cover should be a strategy, which should be associated with adequate land use, such as installation of improved pastures. The improvement of soil quality reached by this system may also positively affect pastures productivity and quality, and therefore increase stocking rate and soil C accumulation.
REFERENCES
Belsky, A.J.; Amundson, R.G.; Duxbury, J.M.; Riha, S.J.; Ali, A.R. and Mwonga, S.M. (1989) – The effects of trees on their physical, chemical and biological environments in a semi-arid savanna in Kenya. J. App.Ecol., 26: 1005-1024. [ Links ]
Blake, G.R. and Hartge, K.H. (1986) – Bulk Density. In: Klute, A. (Ed.) - Methods of Soil Analysis, Part I. Physical and Mineralogical Methods. Agronomy Monograph no. 9 (2nd ed.), p. 363-375. [ Links ]
Caritat, A.; Bertoni, G.; Molinas, M.; Oliva, M. and Dominguez-Planella, A. (1996) - Litterfall and mineral return in two cork-oak forests in northeast Spain. Ann. For. Sci., 53: 1049-1058. [ Links ]
Charley, J.L. and West, N.E. (1977) – Micropatterns of N-mineralization activity in soils of some shrubs dominated semidesert ecosystems of Utah. Soil Biol. Biochem., 9: 357-365. [ Links ]
Corbeels, M., O'Connell, A.M., Grove, T.S., Mendham, D.S. and Rance, S.J. (2003) – Nitrogen release from eucalypt leaves and legume residues as influenced by their biochemical quality and degree of contact with soil. Plant and Soil, 250: 15–28, 2003. [ Links ]
Dahlgren, R.A.; Singer, M.J. and Huang, X. (1997) – Oak tree and grazing impacts on soil properties and nutrients in a California oak woodland. Biogeochemistry, 39: 45-64. [ Links ]
David, T.S. (2000) – Intercepção da precipitação e transpiração em árvores isoladas de Quercus rotundifolia Lam. Dissertação de doutoramento, Lisboa, Universidade Técnica de Lisboa, Instituto Superior de Agronomia, 155 p. [ Links ]
De Leenheer, L. and Van Hove, J. (1958) – Determination de la teneur en carbone organique des sols. Etude critique des méthodes titrimétriques. Pédologie, 8: 39-77. [ Links ]
DGF (2001) – Inventário Florestal Nacional. Portugal Continental. 3ª Revisão 1995-1998. Lisboa, Direcção-Geral das Florestas. [ Links ]
Edmonds, R.L.; Thomas, T.B. and Rhodes, J.J. (1991) – Canopy and soil modification of precipitation chemistry in a temperate rain forest. Soil Sci. Soc. Am. J., 55: 1685-1693. [ Links ]
Egnér, H.; Riehm, H. and Domingo, W.R. (1960) – Untersuchungen über die chemische Bodenanalyse als Grundlage für die Beurteilung des Nährstoff-zustandes der Böden. II Chemische Extraktionmethod zur Phosphor-und Kaliumbestimmung. Kungl. Lantbr. Högsk. Ann., 26: 199. [ Links ]
Escudero, A.; Garcia, B.; Gomez, J.M. and Luis, E. (1985) – The nutrient cycling in Quercus rotundifolia in Quercus pyrenaica ecosystems ("dehesas") of Spain. Acta Oecol-Oec Plant., 6, 20: 73-86. [ Links ]
Gallardo, A. (2003) – Effect of tree canopy on the spatial distribution of soil nutrients in a Mediterranean Dehesa. Pedobiologia, 47: 117-125. [ Links ]
Gallardo, A.; Rodríguez-Saucedo, J.J.; Covelo, F. and Fernández-Alés, R. (2000) – Soil nitrogen heterogeneity in a Dehesa ecosystem. Plant Soil, 222: 71-82. [ Links ]
Guggenberger, G.; Zech, W. and Thomas, R.T. (1995) – Lignin and carbohydrate alteration in particle-size separates of an oxisol under tropical pastures following native savanna. Soil Biol. Biochem., 27: 1629-1638. [ Links ]
Haynes, R.J. and Williams, P.H. (1993) – Nutrient cycling and soil fertility in the grazed pasture ecosystem. Adv. Agron., 49: 119-199. [ Links ]
Herman, D.J.; Halverson, L.J. and Firestone, M.K. (2003) – Nitrogen dynamics in an annual grassland: oak canopy, climate, and microbial population effects. Ecological Applications, 13, 3: 593–604 [ Links ]
Houba, V.J.S.; Novozamsky, I. and Termminghoff, E. (1994) – Soil Analysis Procedures. Wageningen, Wageningen Agricultural University. [ Links ]
IFN (2006) – Inventário Florestal Nacional (2005-2006). Divisão de Inventário e Estatísticas Florestais. Lisboa, Direcção Geral das Florestas. [ Links ]
INMG (1991) – Normas climatológicas da região de "Alentejo e Algarve" correspondentes a 1951-1980. O Clima de Portugal. Fasciculo XLIX, vol. 4 - 4a Região, Lisboa, Instituto Nacional de Metereologia e Geofísica. [ Links ]
IUSS Working Group WRB (2006) – World Reference Base for Soil Resources 2006, vol 103. World Soil Resources Report, 2nd edition. Rome, FAO. [ Links ]
Jackson L.E.; Strauss R.B.; Firestone M.K. and Bartolome J.W. (1990) – Influence of tree canopies on grassland productivity and nitrogen dynamics in deciduous oak savanna. Agr. Ecosyst. Environ., 32: 89-105. [ Links ]
Joffre, R. and Rambal, S. (1988) – Soil water improvement by trees in ragelands of southern Spain. Acta OEcologica, 9: 405-422. [ Links ]
Joffre, R. and Rambal, S. (1993) – How tree cover influences the water balances of Mediterranean rangelands. Ecology, 74: 570-582. [ Links ]
Joffre, R.; Rambal, S. and Ratte, J.P. (1999) – The dehesa system of southern Spain and Portugal as a natural ecosystem mimic. Agroforestry Systems, 45: 57-79. [ Links ]
Lehmann, J.; Poidy, N.; Schroth, G. and Zech, W. (1998) – Short-term effects of soil amendment with tree legume biomass on carbon and nitrogen in particle size separates in Central Togo. Soil Biol. Biochem., 30, 1545-1552. [ Links ]
Lehmann, J.; da Silva, M. and Zech W. (2001) – Organic matter stabilization in a Xanthic Ferralsol of the central Amazon as affected by single trees: chemical characterization of density, aggregate, and particle size fractions. Geoderma, 99: 147-168. [ Links ]
Lima, A.M.N.; Silva, I.R.; Neves, J.C.L.; Novais, R.F.; Barros, N.F.; Mendonça, E.S.; Smyth, T.J.; Moreira, M.S. and Leite, F.P. (2006) – Soil organic carbon dynamics following afforestation of degraded pastures with eucalyptus in southeastern Brazil. Forest Ecol. Manag., 235: 219–231. [ Links ]
Matson, P.A. and Vitousek, P.M. (1981) – Nitrification potentials following clearcutting in the Hoosier National Forest, Indiana. For. Sci. 27: 781-791. [ Links ]
Milne, R.M. and Haynes, R.J. (2004) – Soil organic matter, microbial properties, and aggregate stability under annual and perennial pastures. Biol. Fertil. Soils 39: 172-178. [ Links ]
Moreno, G.; Obrador, J.J.; Cubera, E. and Dupraz, C. (2005) – Fine root distribution in dehesas of central-western Spain. Plant Soil, 277: 153-162. [ Links ]
Moreno, G.; Obrador, J.J.; Garcia, E.; Cubera, E.; Montero, M.J.; Pulido, F. and Dupraz, C. (2007) – Driving competitive and facilitative interactions in oak dehesas through management practices. Agroforest. Syst., 70: 25-40. [ Links ]
Nunes, J. (2005) – Interacção Solo – Árvore Isolada em Montados de Azinho (Quercus rotundifolia Lam.): Processos Fundamentais. Dissertação de doutoramento, Évora, Universidade de Évora, 252 p. [ Links ]
Perakis, S.S. and Kellogg, Ch.H. (2007) – Imprint of oaks on nitrogen availability and d15N in California grassland-savanna: a case of enhanced N inputs? Plant Ecol., 191: 209-220. [ Links ]
Sá, C.S. (2001) – Influência do Coberto Arbóreo (Quercus suber L.) em Processos Ecofisiológicos da Vegetação Herbácea em Áreas de Montado. Dissertação de doutoramento, Évora, Universidade de Évora. [ Links ]
Sá, C.S., Madeira, M. and Gazarini, L. (2004) – Decomposição e libertação de nutrientes da necromassa de comunidades herbáceas. Rev. de Ciências Agrárias 27, 1: 457-469. [ Links ]
Young, A. (1997) – Agroforestry for Soil Management. 2nd Edition. New York, CAB International. [ Links ]
ACKNOWLEDGEMENTS
The study was developed within the activities of the project POCTI/AGG/48914/2002 (Impacto da árvore e da pastagem melhorada na qualidade do solo e sustentabilidade dos montados: Quantidades e dinâmicas do C e do N). We thank to farm owner, David Crespo and family, for facilitation of field sampling. Isabel Pina (Departamento de Química Agrícola e Ambiental) and Laboratory staff of the Departamento de Ciências do Ambiente (Instituto Superior de Agronomia) are acknowledgedfor their technical assistance for analytical work.
Recepção/Reception: 2011.03.21
Aceitação/Acception: 2011.03.29

