










Serviços Personalizados
Journal

Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
 Similares em
SciELO
Similares em
SciELO Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias v.34 n.1 Lisboa jan./jun. 2011
Seleção de genótipos de Brachiaria Ruziziensis quanto à tolerância ao alumínio em solução nutritiva. I: Resposta a diferentes concentrações de alumínio e valores de ph em solução nutritiva
Carlos Eugênio Matins1, Paulo Sérgio Balbino Miguel2, Wadson Sebastião Duarte da Rocha1, Fausto Souza Sobrinho1, Fernando Teixeira Gomes3 e André Vicente de Oliveira1
1Pesquisadores – D. Sc. – Embrapa Gado de Leite Rua Eugênio do Nascimento, 610 – Dom Bosco CEP 36038-300 – Juiz de Fora/MG – Brasil. caeuma@cnpgl.embrapa.
2Estudante de Mestrado da Universidade Federal de Viçosa – Departamento de Microbiologia Agrícola – bolsista da Fapemig; Avenida Peter Henry Rolfs, Campus Universitário, s/n, Centro – CEP 36570-000 – Viçosa/MG – Brasil. paulo.b@ufv.br– (31) 98974905.
3Professor – D. Sc. – Centro de Ensino Superior de Juiz de Fora (CES-JF) Rua Luz Interior, 345, Estrela Sul, 36030776, Juiz de Fora/MG – Brasil.
4Aluno de graduação – Centro de Ensino Superior de Juiz de Fora - Bolsistas de Iniciação Científica do CNPq/CNPGL.
RESUMO
A seleção de gramíneas forrageiras tolerantes ao alumínio tóxico, abundante em solos ácidos, é etapa indispensável em programas de melhoramento que procuram identificar materiais mais adaptados a essas condições de estresse abiótico. Visando aumentar a eficiência das avaliações realizadas em solução nutritiva, foi realizado um experimento em casa de vegetação para avaliar os efeitos de doses crescentes de alumínio e o valor de pH da solução nutritiva no desenvolvimento de genótipos de Brachiaria ruziziensis. Dois tratamentos, com avaliações de pH, foram estudados. Em um deles, o pH da solução nutritiva era corrigido diariamente e mantido em 4,5 ± 0,1 (na primeira semana) e em 4,5 ± 0,1 (no início da segunda semana até o final do experimento) e um outro tratamento sem controle de pH. As concentrações de alumínio utilizadas foram: 0, 15, 30, 45 e 60 mg/L. Foram avaliados: a produção de matéria seca da parte aérea e das raízes (MSPA e MSR), o incremento no crescimento da parte aérea e das raízes (IPA e IR), expresso pela diferença entre o crescimento final e o inicial, razões entre MS e MV da parte aérea (MS/MV – PA) e das raízes (MS/MV - R) e o número de perfilhos (NP). Oaumento na concentração de alumínio em solução nutritiva não afetou o crescimento das raízes (IR), nem a sua matéria seca (MSR), a razão entre a matéria seca e a massa verde da raiz (MS/MV – R) e o número de perfilhos. De modo geral, o controle do pH da solução nutritiva não influenciou a seleção de genótipos de B. ruziziensis quanto à tolerância ao alumínio, proporcionando os menores valores para as características incremento no crescimento das raízes (IR) e razão entre a matéria seca e a massa verde da parte aérea (MS/MV – PA), enquanto as demais características avaliadas apresentaram os maiores valores nos tratamentos sem a correção e manutenção do valor de pH da solução nutritiva.
Palavras-chave: Acidez, braquiária, produção de forragem, tolerância ao alumínio.
Brachiaria ruziziensis genotypes selection. I: Responses under different aluminum concentrations and ph values in nutritive solution
ABSTRACT
The selection of aluminum tolerance tropical forage grass abundant in acid soil is an important step in a plant breeding program that aims to identify more productive Brachiaria ruziziensis genotypes with higher adaptability under stress conditions. It were developed studies aiming to evaluate the more adequated pH value by five genotypes of Brachiaria ruziziensis, congo signal grass, and the effect as crescent aluminum concentrations upon plant growth. pH values were: 4.5 ± 0,1 (in the first week), 4.5 ± 0,1 (in the beginning of the second week to the end of the experiment) and a control treatment without pH control and the aluminum concentrations were: 0, 15, 30, 45 and 60 mg/L of Al. The following variables were evaluated. Dry matter yield of the aerial part and roots (MSPA and MSR), the increasing of aerial part (IPA) and roots (IR), expressed by the difference between the final and initial weight dry matter, rations between dry (MS) and green (MV) matter weight of the aerial part (MS/MV – PA) and root (MS/MV - R) as well as the numbers of tillers. Theincreasing aluminum concentration in nutrient solution had no effect in the increasing of the roots (IR) and dry root matter weight (MSR), the rations between dry (MS) and green (MV) root matter weight (MS/MV - R) and the numbers of tillers, affecting the other characteristics. The absence of pH control in the nutrient solution provided the smallest values for the characteristics, increasing of the roots (IR) and ration between dry (MS) and green (MV) matter weight of the aerial part (MS/MV – PA), while the other evaluated characteristics presented the largest values, when had no pH control and adjustment in the nutrient solution.
Keywords: Acidity, aluminum tolerance, congo signal grass, forage production.
INTRODUÇÃO
No Brasil, a maioria dos solos destinados à produção vegetal apresenta baixa fertilidade e problemas de elevada acidez e toxicidade por alumínio, fatores responsáveis pelo baixo rendimento da maioria das culturas não adaptadas (Souza e Lobato, 2004). No caso específico das pastagens, que normalmente ocupam as áreas marginais, esses problemas são ainda mais sérios. A alta concentração de alumínio nos solos ácidos assume, portanto, papel importante na agricultura e pecuária nacionais, afetando diretamente os processos fisiológicos e metabólicos da grande maioria das espécies cultivadas.
Quando cultivadas em altas concentrações de Al3+, as plantas apresentam sintomas característicos causados pela toxicidade. Dentre esses, destacam-se: o retardamento do crescimento radicular e da parte aérea, a redução do número de raízes laterais, além do maior diâmetro dos eixos radiculares (Mattiello et al., 2008).
O calcário é freqüentemente utilizado com o objetivo de reduzir a acidez do solo, devido ao aumento no valor do pH da solução do solo, aumentar a concentração de cálcio e de magnésio trocáveis, além de diminuir o teor de alumínio tóxico, ferro e manganês, causando maior capacidade de troca catiônica efetiva (CTC). A adição do calcário, também, aumenta a disponibilidade de nitrogênio, enxofre, boro e outros nutrientes que resultam da mineralização da matéria orgânica, melhora o aproveitamento de adubos e estimula a atividade microbiana. No entanto, a sua capacidade corretiva não ultrapassa as camadas superficiais, o que impossibilita o crescimento normal do sistema radicular dos vegetais, que precisam de um maior volume de solo para explorar, principalmente em profundidade, a fim de garantir a absorção de nutrientes e água. Além disso, a redução na saturação de Al ao longo do perfil do solo, com a calagem, necessita, de modo geral, de muito tempo, devido à baixa solubilidade do calcário (Malavolta et al., 1980; Bataglia et al., 1985; Fageria, 1989; Mascarenhas e Tanaka, 1995; Amaral et al., 2004).
A seleção de plantas que toleram diferentes níveis de alumínio vem sendo considerada a melhor alternativa para o aumento da produção em solos ácidos com concentrações altas deste catião. Sendo assim, práticas envolvendo diferentes métodos estão sendo empregadas tanto em cultura de campo como em soluções nutritivas (Echart e Cavalli-Molina, 2001; Sanchez-Chacon et al., 2002). Esta estratégia é considerada indispensável em programas de melhoramento genético, que visam à identificação de plantas mais produtivas e que apresentem maior adaptabilidade em condições de estresse. Além disso, constituições genotípicas superiores que proporcionam melhores ajustes a condições nutricionais adversas, poderão amenizar os problemas de produtividade em solos ácidos ocasionado pelo estresse causado pelo Al (Freitas et al., 2006).
A maioria dos trabalhos realizados sobre toxicidade do Al é conduzida com metodologias que utilizam soluções nutritivas, que não apresentam as desvantagens do trabalho em campo, sujeito às intempéries do meio-ambiente, bem como doenças e pragas, não sendo este elemento o único fator limitante a interferir, de forma negativa, na precisão experimental (Echart e Cavalli-Molina, 2001). Além disso, essa metodologia permite a observação imediata dos efeitos causados pelo alumínio (Camargo et al., 2000).
No Brasil, as espécies de Brachiaria mais cultivadas são B. decumbens, B. brizantha, B. humidicola e B. ruziziensis. Estas espécies são responsáveis pela maior parte das áreas de pastagens cultivadas no Brasil. Embora sejam consideradas plantas tolerantes aos fatores de acidez de solo, há grandes diferenças entre elas.
Em condições de campo a seleção para tolerância ao alumínio é afetada pela variabilidade natural dos solos, bem como pela dificuldade de avaliar danos na raiz, o que pode determinar erros na identificação de genótipos tolerantes. Além disso, pode apresentar o inconveniente de reunir grande número de variáveis não controláveis, tais como pH, concentração de Al, e sua correlação com os teores de Ca, Mg e K presentes na solução do solo (Souza,2001). Por estes motivos é importante que a primeira fase na seleção de clones ou genótipos tolerantes seja desenvolvido em condições controláveis, como laboratório ou casa de vegetação, onde há maior possibilidade de controlar fatores, que via de regra tornam-se difíceis de serem controlados, em condições de campo. Após conhecer os genótipos mais tolerantes, a segunda etapa do trabalho deve ser desenvolvida em condições de campo, para tornar mais realístico os resultados (Mazzocato et al., 2002).
Existe um número considerável de trabalhos que objetivam avaliar o comportamento de gramíneas forrageiras quanto à tolerância ao Al, especialmente, as espécies de Brachiaria (Wenzl et al., 2003) e Panicum maximum (Oliveira et al., 2000). Para a espécie Brachiaria ruziziensis Germ. & C.M.Evrardnão se conhece o efeito do pH da solução nutritiva utilizada nas avaliações de tolerância ao alumínio, nem a dose mais aconselhada para a realização da seleção. Pelo exposto, o objetivo do trabalho foi avaliar os efeitos de doses crescentes de alumínio e o pH da solução nutritiva no desenvolvimento de genótipos de Brachiaria ruziziensis.
MATERIAL E MÉTODOS
Este experimento foi conduzido em casa de vegetação na Embrapa Gado de Leite, em Juiz de Fora/MG, Brasil.
Utilizaram-se cinco genótipos de Brachiaria ruziziensis (P1, P2, P3, P4, P5), componentes do programa de melhoramento genético desta espécie na Embrapa Gado de Leite. Os valores de pH avaliados foram 4,5 e um tratamento sem controle de pH, e as concentrações de alumínio foram: 0, 15, 30, 45 e 60 mg/L.
As sementes das diferentes progênies foram colocadas para germinar em caixas plásticas contendo substrato comercial à base de casca de Pinus. Cerca de 60 dias depois, as mudas, após lavagem do sistema radicular, foram transplantadas para vasos contendo dois litros de solução nutritiva de Clark (1975) modificada em relação à concentração de fósforo (0,069 mol/L) com o objetivo de evitar a sua precipitação pelo alumínio. Nesse momento foi realizada a mensuração dos comprimentos iniciais da parte aérea (CIPA) e das raízes (CIR) de cada planta. Os vasos, mantidos em aeração constante, foram revestidos internamente com filme plástico de polietileno de cor preta e externamente por tinta preta betuminosa e alumínica, a fim de evitar a passagem de luz para o não crescimento de algas (Camargo e Camargo, 1985). Durante 37 dias, em um dos tratamentos o pH da solução foi monitorado diariamente e ajustado com NaOH 0,5 mol/L ou HCl 0,5 mol/L, permitindo-se uma variação de ± 0,1, em função do tratamento. Durante os primeiros sete dias, para adaptação dos genótipos ao ambiente aquático e uniformização dos materiais, as plantas cresceram em solução nutritiva com metade da concentração de nutrientes recomendada, sem aplicação de alumínio, e com pH mantido em 4,5 ± 0,1. Daí em diante, promoveu-se, semanalmente, a troca da solução nutritiva, colocando-se nos vasos solução completa, com concentrações crescentes de alumínio e valores de pH de acordo com os tratamentos. No tratamento sem ajuste, o pH não foi controlado diariamente.
Após o período mencionado, as plantas foram retiradas da solução nutritiva, o sistema radicular foi seccionado da parte aérea, lavado com água destilada (por três vezes) e seu comprimento novamente determinado (CFSR, cm). Também foi medido o comprimento final da parte aérea (CFPA, cm). O excesso de umidade das raízes foi retirado com papel toalha, procedendo-se, em seguida, à pesagem e acondicionamento em sacos de papel para secagem em estufa de circulação forçada de ar (96 horas a 55°C). Determinou-se em seguida a produção de matéria seca da parte aérea e das raízes (MSPA e MR).
Conhecidos os comprimentos inicial e final da parte aérea e raízes, determinou-se o incremento no crescimento da parte aérea (IPA) e das raízes (IR), expressos pela diferença entre o crescimento final e inicial de cada parte. O número de perfilhos (NP) foi também determinado quando da colheita do experimento, assim como as razões entre matéria seca e massa verde da parte aérea (MS/MV – PA) e das raízes (MS/MV - R).
O delineamento experimental foi o de blocos ao acaso, com três repetições, em um esquema fatorial. Cada unidade experimental consistiu de um vaso contendo uma planta. Foi efetuada a análise de variância dos dados e das médias, quando necessário, foram comparadas pelo teste de Scott-Knott (1974), a 5 % de probabilidade.
RESULTADOS E DISCUSSÃO
A concentração de alumínio na solução, o controle e ajuste do pH e os genótipos utilizados foram responsáveis por alterações na maioria das características avaliadas (Quadro 1). Porém, algumas exceções foram verificadas. O controle ou não do pH da solução não influenciou a relação entre a matéria seca e a massa verde das raízes e nem a produção de perfilhos. O aumento na concentração do alúmínio em solução não influenciou nas características relacionadas com o sistema radicular e no número de perfilhos produzidos. Em relação aos genótipos testados não houve diferença nas razões matéria seca/massa verde da parte aérea e das raízes. Tal fato indica a existência de variabilidade genética entre os genótipos de B. ruziziensis para a tolerância ao alumínio (Quadro 1). Desta forma, verificaram-se modificações nas características da planta pela adição de alumínio na solução, tal como foi verificado por Crestani et al. (2009). O tempo de exposição ao alumínio foi suficiente para discrimiar genótipos tolerantes e sensíveis (Quadro 1). Entretanto, o contato por tempo inferior a esse já foram demonstrados como discriminatórios para a seleção de genótipos tolerantes (Pintro et al., 2001, Peixoto et al., 2007, Martins et al., 2010), os quais relatam 7, 10 e 21 dias respectivamente.
Quadro 1 - Incremento no crescimento de parte aérea (IPA) e de raízes (IR), matéria seca de parte aérea e raízes (MSPA e MSR), razões entre MS e MV da parte aérea (MS/MV – PA) e das raízes (MS/MV - R) e número de perfilhos (NP) em cinco genótipos de Brachiaria ruziziensis, em dois valores de pH e cinco concentrações de alumínio, em solução nutritiva.
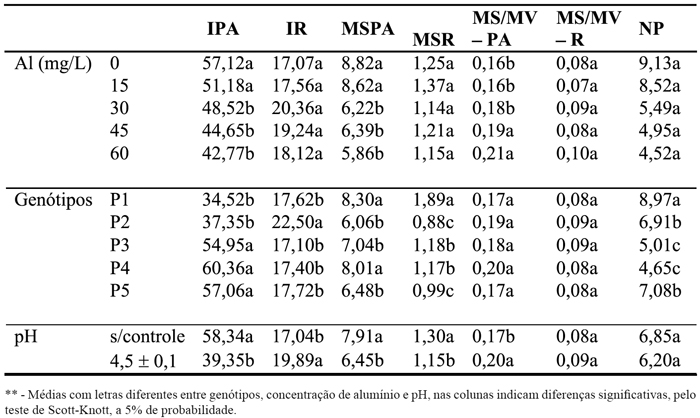
Em 15 mg/L o comportamento dos genótipos de B. ruzizensis foi semelhante ao controle em todas as caracteristicas analisadas, não sendo esta concentração suficiente para separar plantas tolerantes de sensíveis. Com o aumento na concentração de alumínio em solução nutritiva (30, 45 e 60 ppm) as plantas foram afetadas (Quadro 1), especialmente nas características IPA, MSPA, bem como MS/MV – PA não afetando as demais. Fortunato e Nicoloso (2004), em trabalhos com grápia utilizando quatro doses de alumínio (0; 2,5; 5,0 e 10 mg/L), não encontraram alterações significativas para a altura da planta, massa seca da parte aérea e a relação entre a massa de raízes e da parte aérea, comprimento da raiz principal, comprimento total das raízes e massa seca de raízes. Esses resultados corroboram parcialmente com os do presente experimento (Quadro 1), uma vez que a variação na concentração de alumínio não influenciou nas características relacionadas com o sistema radicular e com a produção de perfilhos.
Em 30, 45 e 60 mg/L tanto o IPA quanto a MSPA apresentaram decréscimos quando comparadas com o controle, não sendo as demais características afetadas pelas doses avaliadas. Estes resultados contrariam a tendência geral no que diz respeito a estudos envolvendo tolerância ao alumínio. Isso porque a redução da parte aérea é considerada um sintoma secundário, sendo uma consequência dos danos ocorridos no sistema radicular (Foy et al., 1978). No entanto, tais resultados concordam com os obtidos por Simões et al. (1973), que trabalhando com pinheiro do Paraná demonstraram crescimento reduzido das plantas na presença das maiores doses do metal (2, 3 e 4 ppm), comparadas às menores (0, 0.25, 0.50 e 1 ppm). De acordo com Diniz e Calbo (1990), o excesso de Al pode conduzir à formação de complexos na superfície radicular pela precipitação com fósforo, impedindo sua absorção de fósforo e o crescimento da planta. Para Foy et al. (1978) os sintomas característicos para a parte aérea em estudos relacionados com a toxicidade por alumínio incluem a rigidez das folhas e colmos, além de atrofia e coloração púrpura dessas estruturas, os quais se confundem com os de deficiência de cálcio e fósforo. No entanto, Ruschell et al. (1968), em trabalhos com feijoeiro em solução nutritiva, observaram que os efeitos deletérios na parte aérea eram provocados pelo excesso de alumínio e não pela deficiência de fósforo nesses órgãos.
O efeito negativo do alumínio sobre o desenvolvimento dos vegetais é expresso pela redução na matéria seca da parte aérea e raízes (Rengel, 1996), o que ocorre devido à inibição da expansão e divisão celular no sistema radicular e bloqueio da absorção de nutrientes para a parte aérea (Lazof e Holland, 1999). No presente experimento, entretanto, o aumento gradativo das concentrações de alumínio na solução afetou a produção de matéria seca da parte aérea, mas não das raízes (Quadro 1). Esses resultados diferem dos apresentados por Nascimento et al. (1997) que ao estudarem o comportamento de Coffea arabica frente ao alumínio encontraram resultado inverso, para as mesmas variáveis. Nolla et al. (2007) e Mattiello et al. (2008), por sua vez, em soja e café conilon, respectivamente, verificaram decréscimos tanto para matéria seca da parte aérea quanto para as raízes quando aumentadas às doses de alumínio. Ao comparar a tolerância entre milho e sorgo, Akhter et al. (2009) demonstraram menor efeito inibitório do alumínio na matéria seca do milho em relação ao sorgo, indicando maior tolerância do primeiro quando comparado ao segundo. Os mesmos autores ainda relataram que o alumínio em solução diminuiu não só a produção de matéria seca, mas também, a concentração de nutrientes tais como P, Ca, K, Mg, Fe e Mn, sendo esta redução maior no sorgo do que no milho. Por outro lado, em espécies consideradas tolerantes ao alumínio, a produção da parte aérea é comumente mais afetada que o sistema radicular (Jan e Petterson, 1995). Assim, neste experimento a produção de massa seca da parte aérea mostrou-se mais sensível para discriminar o efeito do alumínio no crescimento de B. ruziziensis (Quadro 1). Neste sentido, os genótipos 1 e 4 são os que mais se destacam quando considerada a MSPA, com o primeiro apresentando maior número de características favoráveis (MSPA, MSR e NP) para considerá-lo mais tolerante. Tal fato demonstra serem estes genótipos os mais promissores para a utilização em estudos posteriores em condições de campo.
O crescimento do sistema radicular é a característica mais afetada tanto na susceptibilidade quanto na tolerância a fatores de acidez do solo (Howeler e Cavadid, 1976), contrastando com os resultados deste experimento. No entanto, Fortunato e Nicoloso (2004), em grápia (Apuleia leiocarpa) não observaram diferenças significativas no crescimento radicular para as concentrações 0; 2,5; 5,0 e 10 mg/L. Eles relataram, ainda, que as plantas submetidas a 10 mg/L não apresentavam sintomas visuais nas raízes de toxicidade ao Al, apresentando-se claras, normais e sem alterações morfológicas, sendo o mesmo verificado neste experimento em 15 mg/L (Quadro 1). Porém, com o aumento das concentrações as plantas apresentavam-se grossas, amareladas, quebradiças e pouco volumosas nas concentrações mais altas de alumínio (30, 45 e 60 mg/L). Esse engrossamento, provavelmente, é causado pela inibição do alongamento celular e aumento de algumas camadas de células do córtex (Sasaki et al., 1996). Haussler et al. (2006), por outro lado, demonstraram que B. ruziziensis e B. dictyoneura cultivadas em solos com alta saturação de alumínio (76 a 87%) são capazes de desenvolverem o sistema radicular sem apresentarem sintomas de toxicidade a esse elemento.
Os genótipos de B. ruziziensis foram influenciados pelas doses de alumínio e condições de pH utilizados, sendo que nas características MS/MV – PA e MS/MV – R, não houve diferença entre os genótipos (Quadro 1). O genótipo 1 apresentou melhor desempenho, pois teve maior produção de matéria seca de raízes, perfilhou mais (NP) e junto com o genótipo 4 produziu mais matéria seca da parte aérea. Porém, em relação ao crescimento da parte aérea (IPA) e das raízes (IR), o genótipo 1 não foi superior aos demais (Quadro 1), mas, o seu menor crescimento não influenciou, de forma negativa, a produção de massa, que é um dos atributos mais importantes para selecionar o material a ser utilizado para a alimentação animal.
A razão entre MS e MV das raízes (MS/MV - R) e o número de perfilhos não foram afetados pelo pH (Quadro 1). Todas as demais características apresentaram efeito significativo.
Há que destacar que as características incremento no crescimento das raízes (IR) e razão entre a matéria seca e a massa verde da parte aérea (MS/MV – PA) apresentaram valores menores quando não se corrigiu o pH da solução nutritiva (s/ controle), enquanto as demais características avaliadas apresentaram os maiores valores quando não se corrigiu o pH da solução nutritiva. Foi verificado que mais de 70% das características avaliadas não foram prejudicadas pelo não controle e ajuste do valor de pH da solução nutritiva. Portanto, nas condições em que se decorreram o experimento, a realização da correção do pH da solução nutritiva durante o desenvolvimento da braquiária não é necessária. Este resultado é de grande praticidade, pois os ajustes diários nos valores de pH, oneram o custo de experimentos como este, principalmente se forem utilizados muitos vasos.
Almeida et al. (2000) trabalhando com 30 genótipos de Panicum maximum em solução nutritiva, verificaram, para a maioria deles, comportamento diferenciado, com inibição do comprimento das raízes na presença do Al independentemente do pH.
CONCLUSÕES
Os genótipos 1 e 4 são os que mais se destacaram quanto a tolerância ao alumínio nesse estudo, o que demonstra serem eles os mais promissores para a utilização em estudos posteriores em condições de campo.
Oaumento na concentração de alumínio em solução nutritiva não afetou o incremento no crescimento das raízes (IR) e a produção de matéria seca das raízes (MSR), a razão entre a matéria seca e a massa verde da raiz (MS/MV – R) e o número de perfilhos, afetando todas as demais características (IPA, MSPA e MS/MV – PA).
A ausência de controle e ajuste do valor de pH da solução nutritiva proporcionou os menores valores em somente duas características avaliadas, incremento no crescimento de raízes (IR) e razão entre a matéria seca e a massa verde da parte aérea (MS/PV – PA), enquanto as demais características avaliadas (70%) apresentaram os maiores valores, quando não se corrigiu o pH da solução nutritiva.
Não há necessidade de controlar e ajustar o pH da solução nutritiva, diariamente, nos estudos com Brachiaria ruziziensis.
Em função dos resultados obtidos, uma segunda etapa, em condições de campo será desenvolvida para comprovação dos resultados.
REFERÊNCIAS BIBLIOGRÁFICAS
Akhter, A.; Khan, S.H.; Hiroaki, E.; Tawaraya, K.; Rao, I.M. e Wenzl, P. (2009) - Greater contribution of low-nutrient tolerance to sorghum and maize growth under combined stress conditions with high aluminum and low nutrients in solution culture simulating the nutrient status of tropical acid soils. Soil Science and Plant Nutrition 55, 3/6: 394–406. [ Links ]
Almeida, A.A.S.; Monteiro, F.A. e Jank, L. (2000) - Avaliação de Panicum maximum Jacq. para tolerância ao alumínio em solução nutritiva. Revista Brasileira de Ciência do Solo, 24, 1/6: 339-344. [ Links ]
Amaral, A.S.; Anghinoni, I.; Hinrichs, R. e Bertol, I. (2004) - Movimentação de partículas de calcário no perfil de um cambissolo em plantio direto. Revista Brasileira de Ciência do Solo, 28, 1/6: 359-367. [ Links ]
Bataglia, O.C.; Camargo, C.E.O.; Oliveira, O.F.; Nagai, V. e Ramos, V.J. (1985) - Resposta da calagem de três cultivares de trigo com tolerância diferencial ao alumínio. Revista Brasileira de Ciência do Solo, 9, 1/6: 129-147. [ Links ]
Camargo, C.E.O e Camargo, O.B.A. (1985) - Estudo comparativo da tolerância à toxicidade de Ferro e Alumínio em arroz. Bragantia, 44, 1/6: 97-114. [ Links ]
Camargo, C.E. de O.; Ferreira Filho, A.W.P e Felicio, J.C. (2000) - Herança da tolerância ao alumínio em populações híbridas de trigo. Pesquisa Agropecuária Brasileira, 35, 3/12: 517-522. [ Links ]
Clark, R.B. (1975) - Caracterization of phosphatase in intact maize roots. Journal of Agricultural and Food Chemistry, 23, 3/4: 458-460. [ Links ]
Crestani, M.; Carvalho, F.I.F.; Oliveira, C.O.; Silva, J.A.G.; Souza, V.Q.; Parachu, E. A.M.; Silveira, G.; Ribeiro, G. e Luche, H.S. (2009) - Estresse por alumínio em genótipos de aveia preta em condição hidropônica. Bragantia, 68, 3/4: 639-649. [ Links ]
Diniz, V.P.M. e Calbo, M.E.R. (1990) - Efeito da aplicação foliar de fósforo sobre a toxidez de alumínio em plantas de tomate. Revista Brasileira de Fisiologia Vegetal, 2, 2/3: 57-61. [ Links ]
Echart, C.L. e Cavalli-Molina, S. (2001) - Fitotoxicidade do alumínio: efeitos, mecanismo de tolerância e seu controle genético. Revista Ciência Rural, 31, 3/4: 531-541. [ Links ]
Fageria, N.K. (1989) - Solos Tropicais e Aspectos Fisiológicos das Culturas. Brasília, EMBRAPA – DPU, 425 p. [ Links ]
Foy, C.D.; Chaney, R.L. e White, M.C. (1978) - The physiology of metal toxicity in plants. Annual Review of Plant Physiology, 29, 1/1: 511-566. [ Links ]
Fortunato, R.P. e Nicoloso, F.T. (2004) - Toxidez de alumínio em plântulas de grápia (Apuleia leiocarpa Vog. Macbride). Revista Ciência Rural, 34, 1/6: 89-95. [ Links ]
Freitas, F.A.; Kopp, M.M.; Souza, R.O.; Zimmer, P.D.; Carvalho, F.I.F. e Oliveira, A.C. (2006) - Absorção de P, Mg, Ca e K e tolerância de genótipos de arroz submetidos a estresse por alumínio em sistemas hidropônicos.Revista Ciência Rural, 36, 1/6: 72-79. [ Links ]
Haussler, K.; Rao, I.M.; Schultzekraft, R. e Marschner, L.H. (2006) - Shoot and root growth of two tropical grasses, Brachiaria ruziziensis and B. dictyoneura, as influenced by aluminium toxicity and phosphorus deficiency in a sandy loam Oxisol of the eastern plains of Colombia. Tropical Grasslands,40, 4/4: 213–221. [ Links ]
Howller, R.H. e Cavadid, L.F. (1976) - Screening of rice cultivars for tolerance to Al-toxicity in nutrients solutions as compared with a field screening method. Agronomy Journal, 68, 4/6: 551-555. [ Links ]
Jan, F. e Pettersson, S. (1995) - Aluminium sensitivy of two upland rice cultivars at various levels of nutrient suplly. Journal Plant nutrition, 18, 6/12: 1323-1335. [ Links ]
Lazof, D.B. e Holland, M.J. (1999) - Evaluation of the aluminium-induced root growth inhibition in isolation from low pH effects in Glycine max, Pisum sativum and Phaseolus vulgaris. Australian Journal of Plant Physiology, 26, 2/8:147-157. [ Links ]
Malavolta, E (1980) - Elementos de Nutrição Mineral de Plantas. São Paulo, Editora Agronômica Ceres Ltda., 251 p. [ Links ]
Martins, C.E.; Souza-Sobrinho, F.; Gomes, F.T.; Rocha, W.S.D. e Brighenti, A.M. (2010) - Tolerância à toxidez por alumínio em capim-elefante. Pesquisa Aplicada & Agrotecnologia, 2,1/4:21-28. [ Links ]
Mascarenhas, H.A.A. e Tanaka, R.T. (1995) - Crescimento em vasos, de cultivares de soja e de trigo em função da saturação de Alumínio.Revista Scientia Agricola, 52, 2/3: 257-262. [ Links ]
Matttiello, E.M.; Pereira, M.G.; Zonta, E.; Mauri, J.; Matiello, J.D.; Merieles, P.G. e Silva, I.R. (2008.) - Produção de matéria seca, crescimento radicular e absorção de cálcio, fósforo e alumínio por Coffea canephora e Coffea arabica sob influência da atividade do alumínio em solução. Revista Brasileira de Ciência do Solo, 32,1/6: 425-434. [ Links ]
Mazzocato, A.C.; Rocha, P.S.G.; Sereno, M.J.C.M.; Bohnen, H.; Grongo, V. e Neto, J. F.B. (2002) - Tolerância ao alumínio em plântulas de milho. Revista Ciência Rural, 32, 1/6: 19-24. [ Links ]
Nascimento, C.W.A.; Souza Júnior, J.O.; Martinez, H.E.P.; Teixeira, P.R.G. e Fontes, P.C.R. (1997) - Absorção de fósforo e alumínio em cafeeiro (Coffea arabica) cultivado em solução nutritiva com sistema radicular parcialmente submetido a doses de alumínio. Revista Ceres, 44, 2/6: 253:31-338. [ Links ]
Nolla, A.; Schlindwein, J.A. e Anghinoni, I. (2007) - Crescimento, morfologia radicular e liberação de compostos orgânicos por plântulas de soja em função da atividade de alumínio na solução do solo de campo natural. Revista Ciência Rural, 37, 1/6: 97-101. [ Links ]
Oliveira, A.C.; Usberti Filho, J.A. e Siqueira, W.J. (2000) - Nova metodologia de avaliação da reação de genótipos de capim-colonião ao alumínio. Pesquisa Agropecuária Brasileira, 35, 11/12: 2261-2268. [ Links ]
Peixoto, P.H.P.; Pimenta, D.S. e Cambraia, J. (2007) - Alterações morfológicas e acúmulo de compostos fenólicos em plantas de sorgo sob estresse de alumínio. Bragantia, 66, 1/6: 17-25. [ Links ]
Pintro, J.C. (2002) - Efeitos da toxicidade de duas espécies químicas de alumínio (Al) sobre o crescimento de plantas de milho cultivadas em solução nutritiva. Acta Scientiarum, 24, 5/5, 1219-1222. [ Links ]
Rengel, Z. (1996) - Uptake of aluminium by plants cells. New Physiology, 134, 3/4: 389-406. [ Links ]
Ruschel, A.P.; Alvahydo, R. e Sampaio, I.B.M. (1968) - Influência do excesso de Al no feijão (P. vulgaris L.) cultivado em solução nutritiva. Pesquisa Agropecuária Brasileira, 3, 2/12: 229-233. [ Links ]
Sanchez-Cacon, C.D.; Federeizzi, L.C.; Milach, S.C.K. e Pacheco, M.T. (2002) - Variabilidade genética e herança da tolerância à toxicidade do alumínio em aveia. Pesquisa Agropecuária Brasileira, 35, 9/12: 1797-1808. [ Links ]
Sasaki, M.; Yamamoto, Y. e Matsumoto, H. (1996) - Lignin deposition induced by aluminium in wheat (Triticum aestivum) roots. Physiologya Plantarum, 96, 2/12: 193-198. [ Links ]
Scott, A.J. e Knott, M.A.A. (1974) - Cluster analysis method for grouping means in the analysis of variance. Biometrics, 30, 3/4: 507-512. [ Links ]
Souza, L.A.C. (2001) - Reação de genótipos de soja ao alumínio em hidroponia e no solo. Pesquisa Agropecuária Brasileira, 36, 10/12: 1255-1260. [ Links ]
Simões, J.W.; Couto, H.T.Z. e Kajyia, S. (1973) - Tolerância do pinheiro do paraná (Araucaria angustifolia (Bert.) O. Ktze) a teores crescentes de alumínio.IPEF, 6, 2/2: 93-102. [ Links ]
Sousa, D.M. e Lobato, E. (2004) - Cerrado: correção do solo e adubação. Brasília, Embrapa Informação Tecnológica, 416 p. [ Links ]
Wenzl, P.; Mancilla, L.I.; Mayer, J.E.; Albert, R. e Rao, I.M. (2003) - Simulating acid soils with nutrient solutions: The effects on Brachiaria species. Soil Science Society of American Journal, 67, 5/6: 1457-1469. [ Links ]
Recepção/Reception: 2010.04.13
Aceitação/Acception: 2011.03.01

