










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias vol.36 no.2 Lisboa abr. 2013
Influencia de presas criadas sobre maíces Bt sobre parámetros biológicos de Eriopis connexa (Coleoptera: Coccinellidae)
Influence of prey reared on Bt maize on biological parameters of Eriopis connexa (Coleoptera: Coccinellidae)
María Cecilia Curis e María Amalia Trode Isabel Bertolaccini*
* Zoología Agrícola, Departamento de Producción Vegetal, Facultad de Ciencias Agrarias, Universidad Nacional del Litoral, R. P. Kreder 2805, 3080-Esperanza, Santa Fe, Argentina. E-mail: mcuris@fca.unl.edu.ar, author for correspondence
RESUMEN
La toxina Bt en los cultivos genéticamente mejorados es selectiva, pero su expresión continua en los vegetales puede afectar a la fauna benéfica. Con el objetivo de evaluar el efecto de maíces Bt en la duración del estado larval y el peso de los adultos de Eriopis connexa se establecieron cuatro tratamientos, con 50 repeticiones, consistentes en alimentar larvas ad libitum con Sipha maydis y Spodoptera frugiperda, criadas en maíz convencional y Bt, hasta adultos. Para determinar la incidencia sobre la fecundidad y la fertilidad se separaron adultos de cada tratamiento en parejas, manteniendo la alimentación. Los estudios se realizaron en condiciones controladas. Mediante ANOVA y contrastes ortogonales se determinaron diferencias entre tratamientos, éstas fueron significativas entre presas. Con S. frugiperda sobre maíz Bt la duración del estado larval fue mayor y el peso de los adultos y la fecundidad fueron menores. No hubo diferencias con S. maydis, en cualquiera de los maíces.
Palabras clave: Eriopis connexa, maíz Bt, plagas no target, Sipha maidis, Spodoptera frugiperda
ABSTRACT
The Bt toxin is selective in genetically improved crops, but its constant expression on the vegetables can affect the beneficial fauna. With the objective of assessing the effects of Bt corn on the duration of the larval cycle and weight of the Eriopis connexa adult, four treatments were established, with fifty repetitions, consisting on feeding larvae ad libitum with Sipha maydis and Spodoptera frugiperda, raised on both conventional and Bt corn until adulthood. In order to evaluate the incidence on fecundity and fertility, the adults of each group were separated in couples, with the same feeding. The studies were conducted under controlled conditions. Differences among treatments were determined using ANOVA and orthogonal contrasts, and these were significant among preys. With S. frugiperda on Bt corn the larval cycle was longer while the adult weight and fecundity were lesser. There were no differences with S. maydis on any of the kinds of corn.
Keywords: Bt maize, Eriopis connexa, non target pests, Sipha maydis, Spodoptera frugiperda
Introduccion
El maíz modificado genéticamente que contiene d-endotoxinas (Cry1Ab) de Bacillus thuringiensis Berliner (maíz Bt) es uno de los métodos más avanzados para el control de plagas, principalmente del orden Lepidoptera (plagas target). Sin embargo existe preocupación por los posibles efectos sobre el medio ambiente y sobre diversos grupos de organismos non-target en los agroecosistemas.
La toxina Cry1Ab se considera muy selectiva en su acción, por lo que se espera que los efectos sobre artrópodos non-target asociados con el maíz sean mínimos, sin embargo, su expresión continua en los tejidos vegetales, ocasiona una constante exposición de los insectos, que no ocurre con el uso tradicional de B. thuringiensis en las formulaciones de bioinsecticida (Dutton et al., 2003).
La productividad del maíz se ve afectada por factores de estrés, tanto bióticos como abióticos. Entre los factores bióticos, las plagas son la causa de importantes pérdidas. Entre ellas tanto Spodoptera frugiperda Smith (Lepidoptera: Noctuidae), la oruga cogollera, como los pulgones o áfidos (Hemiptera: Aphididae), son considerados como unos de los principales organismos que afectan la producción de este cultivo, principalmente en regiones tropicales y subtropicales del mundo (Viana et al., 2004).
La oruga cogollera siempre fue considerada una plaga importante para la agricultura. A pesar de ser polífaga, presenta una marcada preferencia por las gramíneas, en especial el maíz y el sorgo (Peairs y Saunders, 1981). Debido a que forma grandes poblaciones y a que tiene una alta tasa de dispersión de adultos, se convierte en una plaga extremadamente peligrosa siendo una de la más destructiva del maíz en el continente americano. Los ataques se producen con gran rapidez y solo son detectados cuando ya hay daño irreparable en el cultivo (Artigas, 1994).
Los áfidos son un grupo de fitófagos comunes en los cultivos de maíz en todo el mundo (Dicke y Guthrie, 1988). Sipha maydis Rungsia es una especie que, hasta el año 2002 su distribución se limitaba a Europa, Rusia y Norte de África, colonizando hojas e inflorescencias de maíz, avena y trigo (Bonnemaison, 1980). A partir de ese año fue citada por primera vez para nuestro país en las provincias de Entre Ríos, Córdoba y Mendoza (Ríos de Saluso, 1989; Delfino, 2002; Ortego y Difabio, 2002) y su expansión preocupa.
Dentro de los insectos útiles a la agricultura, los coccinélidos (Coleoptera: Coccinellidae) forman un complejo de especies cuyo rol es importante en el establecimiento del control natural de insectos plagas en muchos agroecosistemas. Se caracterizan por su amplio rango de aceptación de presas, fundamentalmente son conocidos como consumidores de áfidos, aunque se los cita también como reguladores poblacionales de cochinillas, de estados juveniles de lepidópteros, coleópteros, himenópteros, dípteros, tisanópteros, etc. (Hodek, 1973). Una de las especies más abundantes y bien adaptadas en los sistemas agrícolas de la zona centro de la provincia de Santa Fe es Eriopis connexa Germar. En la región central de la República Argentina, su presencia fue registrada desde los meses de octubre hasta mayo en cultivos de alfalfa y cereales (Salto et al., 1990).
La toxina Bt tiene efectos indirectos sobre la fauna benéfica debido a que al aumentar la mortalidad de herbívoros presa aumenta consecuentemente la de los predadores, dependiendo tanto de la magnitud de la mortalidad que ocasiona, como de la importancia del herbívoro en la dieta del enemigo natural. Además, según Jervis y Copland (1996), la toxina puede tener efectos subletales sobre los herbívoros al reducir su calidad como alimento.
Mucha de la bibliografía disponible referida al efecto de las plantas trangénicas sobre los enemigos naturales se han realizado sobre Chrysoperla carnea (Stephens) (Neuroptera: Chrysopidae) (Hilbeck et al., 1998 a;b; Dutton et al., 2003; Pilcher et al., 2005) y sobre coccinélidos (Bai et al., 2005; Lundgren et al., 2005; Zhu et al., 2006 Zhang et al., 2006a;b), con resultados dispares. En cambio no se dispone de información sobre el impacto que los cultivos trangénicos ejercen en las especies nativas de Suramérica. El objetivo de este estudio fue evaluar los efectos de presas criadas con maíz Bt sobre la duración de los estadios larvales, la duración total de estado larval y pupal, el peso, la fecundidad y fertilidad de E. connexa.
Materiales y Metodos
Los trabajos fueron realizados en la cámara de cría perteneciente al Departamento de Producción Vegetal de la Facultad de Ciencias Agrarias (Universidad Nacional del Litoral), ubicada en la ciudad de Esperanza, Provincia de Santa Fe, Argentina.
Material vegetal (Maíces Bt y convencionales): Para la obtención de las plántulas de maíz que sirvan de soporte a las crías de S. frugiperda y de S. maydis, se realizaron siembras continuas, dos veces por semana, de maíces Bt (evento DK90MGRR2) y convencional (híbrido 38A57) sobre sustrato de tierra y arena en bandejas plásticas de 25 cm x 40 cm.
Condiciones de la cría de insectos: La cría se inició con la recolección periódica a campo, durante los meses de verano, de adultos y larvas de E. connexa, de colonias de S. maydis y de larvas de S. frugiperda, sobre cultivos de maíces convencionales, en la zona de influencia de la Facultad de Ciencias Agrarias.
• E. connexa: Los adultos recolectados se mantuvieron en recipientes plásticos cilíndricos de 15 cm de alto por 15 cm de diámetro, cubiertos en su parte superior con muselina. Diariamente fueron alimentados ad libitum con los pulgones criados sobre el híbrido convencional. Con igual procedimiento se criaron las larvas hasta el nacimiento de los adultos, los que una vez emergidos se colocaron en grupos de 20 individuos a fin de formar parejas, las que se colocaron separadamente. Diariamente se separaron los desoves. Se continuó con este procedimiento hasta lograr una población continua del depredador.
• S. frugiperda: las larvas recolectadas a campo fueron criadas individualmente, para evitar el canibalismo. Diariamente se alimentaron con plantas frescas del híbrido convencional, momento en que se procedió a retirar el material vegetal seco, procedimiento se continuó hasta la obtención de las pupas, las que se colocaron en conjunto en jaulas de 50 cm de largo, 40 cm de ancho y 40 cm de alto, con tres paredes de muselina y la superior con vidrio para la emergencia de los adultos. La finalidad fue la obtención de los desoves, los que diariamente fueron separados y a partir de los cuales se criaron las larvas en forma individual. Este procedimiento se continuó de igual modo hasta lograr una producción continua de larvas y una cantidad que permita la realización del trabajo. Para la realización de los estudios solo se utilizaron larvas de primer y segundo estadio, a fin de facilitar la captura por las larvas del predador. Previamente se las alimentó durante 24 horas sobre el material genético correspondiente.
• S. maydis: las colonias recolectadas procedentes de maíces a campo fueron llevadas a laboratorio. Inmediatamente fueron inspeccionadas con el fin liberarlas de predadores. Para asegurarse de la eliminación de parasitoides, las colonias se mantuvieron en jaulas separadas del resto de población de áfidos. Los individuos parasitados fueron eliminados y el resto de la población se crió sobre el híbrido convencional. Periódicamente, dependiendo del aumento poblacional, las colonias fueron divididas y repicadas en plántulas nuevas, hasta lograr un nivel poblacional adecuado y constante.
Tanto las crías de los insectos presas y del predador como los ensayos se realizaron en cámaras de cría bajo condiciones controladas de temperatura (25 ± 2ºC), fotoperíodo (16:8 hs luz/oscuridad) y 60 ± 10% de humedad relativa.
• Recolección de polen: se recolectaron inflorescencias masculinas de los mismos cultivares utilizados en los ensayos en laboratorio, las que se colocaron en bolsas de papel y se mantuvieron en condiciones de 5-6ºC, hasta la realización de los estudios de fecundidad y fertilidad.
• Diseño experimental: El estudio consistió en la realizaron cuatro tratamientos que consistieron en alimentar a larvas recién nacidas de E. connexa con: S. maydis criados con maíz Bt; 2) S. maydis criado con maíz convencional; 3) S. frugiperda criadas con maíz Bt y, 4) S. frugiperda criadas con maíz convencional.
• Influencia de las presas en el ciclo de E. connexa: Para cada tratamiento se realizaron 50 repeticiones, en caso de muerte de las larvas éstas no se reemplazaron. Para ello, mediante un pincel fino, se colocaron individualmente larvas recién nacidas del predador, provenientes de varios desoves, en cajas de Petri de 5 cm de diámetro. Diariamente se las alimentó ad-libitum con las presas señaladas y se registraron las mudas de los distintos estadios, hasta que empuparon, determinando la duración de este estado. Una vez nacido el adulto se los pesó, mediante balanza (Scaltec SBA 041) de precisión de tres decimales.
• Influencia de las presas en los parámetros reproductivos de E. connexa: Para determinar la influencia de las presas en la fecundidad y en la fertilidad del predador se utilizaron los adultos provenientes de los ensayos anteriores, los que, recién emergidos se colocaron juntos pero agrupados por tratamiento, en jaulas de 30 cm de alto, 25 cm de largo y 25 cm de altura, con la finalidad de separar las parejas que se encontraban copulando, obteniendo 20 parejas para S. maydis y 10 para S. frugiperda. Detectadas las parejas éstas fueron retiradas y se colocaron en recipientes plásticos de 5 cm de diámetro y 7 cm de alto, en cuyo interior de los mismos se colocó papel absorbente para que sirviera de soporte a los desoves y para facilitar su retiro. La alimentación ad libitum de las parejas se continuó del mismo modo según el tratamiento del que provenían los adultos, adicionando además, según corresponda, polen proveniente de maíces Bt y no Bt. Para ello diariamente se colocaron 3-4 flores individuales masculinas, las que se renovaron diariamente. Los desoves se retiraron y se registró el número de huevos (fecundidad) y se colocaron individualmente a fin de determinar el número de larvas nacidas (fertilidad). Las observaciones se continuaron durante 30 días.
• Análisis estadísticos: Los datos fueron analizados estadísticamente mediante ANOVA. Para determinar la existencia de diferencias entre los tratamientos planteados, se realizó una prueba de contrastes ortogonales previa transformación de los datos en log x con un nivel de significancia P< 0,01. Los datos fueron procesados mediante el software InfoStat (Universidad Nacional de Córdoba, Córdoba, Argentina, 2011).
Resultados
Duración de los estadios larvales y duración total de los estados larvales y pupal.
La duración de los estadios larvales y la duración total del estado larval y pupal se vieron afectados por la dieta. En el primer estadio se hallaron diferencias significativas entre las presas, con un 25,8 y un 40,4% mayor cuando el alimento fue S. frugiperda, según provinieran de maíces no Bt y Bt, respectivamente. Dentro de las variables correspondientes a la misma presa no se registraron diferencias, ya que cuando el alimento fue S. maydis criado en maíz convencional la duración en días fue solo un 3,4% menor que cuando las presas se criaron sobre maíces Bt, mientras que cuando la presa fue S. frugiperda alimentadas con maíces Bt, la duración del estadio se prolongó en un 15,4% (Cuadro 1).
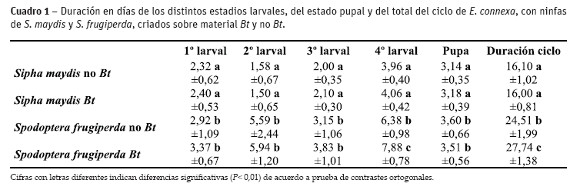
Similar comportamiento se registró para el segundo estadio ya que con respecto a las distintas presas suministradas la duración fue de 253,8% y 296,0% mayor cuando el alimento fueron larvas del lepidóptero, criados con maíz no Bt y Bt, respectivamente, aunque en ninguno de los dos casos existieron diferencias según si provinieran de una alimentación con maíz Bt o convencional. También se observaron diferencias según la presa en el tercer estadio larval aunque las mismas no fueron tan marcadas alargándose el estadio en un 57,5% y 90,5% cuando la alimentación fue S. frugiperda en maíz convencional y Bt respectivamente (Cuadro 1).
La duración del cuarto estadio fue 2,5% mayor cuando la alimentación fue con S. maydis cridado sobre maíces Bt, sin diferencias significativas con respecto al pulgón criado sobre maíz convencional. Entre las presas las diferencias fueron significativas, siendo la duración del estadio 61,1 y 98,9 % mayor cuando se utilizaron larvas del lepidóptero, en maíces no Bt y Bt, respectivamente. Cuando el alimento fueron larvas de S. frugiperda la duración del estadio fue un 23,5% mayor cuando la cría provino de material transgénico, con respecto al convencional siendo la diferencia significativa.
El estado pupal también se vio afectado por la dieta, con diferencias significativas entre las distintas presas, siendo un 14,6 y un 10,4% mayor cuando la alimentación fue con larvas de S. frugiperda, criadas con maíz convencional y Bt, respectivamente. En el caso del áfido no hubo diferencias según fueron criados con material transgénico o convencional, mientras que en el caso del lepidóptero, la duración fue un 2,5% menor cuando las larvas provenían de maíces Bt, aunque sin diferencias significativas con respecto al no Bt.
En la duración total del ciclo no se hallaron diferencias significativas en la alimentación con pulgones provenientes de la cría sobre diferentes germoplasma, ya que en ambos casos fue cercano a los 16 días. En cambio cuando la presa fueron larvas de lepidópteros se halló un incremento en la duración total del ciclo de un 52,2% cuando las mismas fueron criadas sobre maíces genéticamente modificados (Cuadro 1).
Cuando en la alimentación se emplearon áfidos no se observaron muertes de larvas en ningún tratamiento ni estadio del desarrollo, concluyéndose el trabajo con las 50 larvas iniciales. En cambio, cuando se emplearon larvas del lepidóptero hubo una mortalidad que fue del 32% y 30%, cuando fue alimentado con maíz convencional y Bt, lográndose finalizar el ensayo con 34 y 35 individuos, que llegaron a completar el ciclo, respectivamente. En ambos casos el 80% de las muertes se produjo durante el primer estadio de desarrollo del predador.
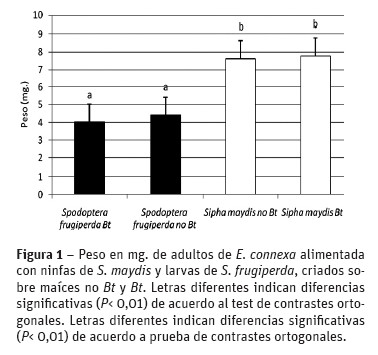
Peso del adulto
En el peso de los adultos no hubo diferencia cuando las larvas de E. connexa fueron alimentadas con S. maydis criada en maíz no Bt y Bt, pero sí se ob servaron diferencias altamente significativas y una disminución del 49% del mismo cuando su dieta fue S. frugiperda (Figura 1). En este último caso no se registraron diferencias en el peso según S. frugiperda fuera criada en maíces no Bt y Bt.
Fecundidad
La fecundidad no se vio influenciada cuando la dieta fue S. maydis criada en maíz no Bt y Bt, mientras que cuando se alimentó con S. frugiperda las diferencias fueron significativas siendo la cantidad de huevos puestos un 46% menor. También se hallaron diferencias significativas en la cantidad de huevos entre E. connexa criadas con S. frugiperda sobre maíz convencional o Bt siendo un 63% menor la cantidad de huevos en este último. En los 30 días de observación se obtuvieron 9 desoves del depredador con una cantidad de huevos por desove entre 14 y 39 para S. maydis criada en maíz convencional y entre 17 y 39 para S. maydis criada en maíz Bt. Para S. frugiperda criada sobre maíz convencional los desoves fueron 8, con una variación en el número de huevos por desove que variaron de 3 a 28; en el caso de S. frugiperda criada en maíz Bt las oviposiciones fueron 5 con una cantidad de huevos que varío entre 3 a 17. El número de huevos fue disminuyendo al final del período de estudio.
Fertilidad
La fertilidad, representada por la cantidad de individuos nacidos por postura, no se vio afectada por ninguna de las dietas ofrecidas. Los valores alcanzados fueron: 50,52% para el tratamiento S. maydis no Bt, 50,48% para S. maydis Bt; 50,42% para frugiparda no Bt y 50,41% para S. frugiperda Bt.
Discusión
Los maíces trangénicos que expresan el gen de la dendotoxina de la bacteria B. thuringiensis (Bt), son una tecnología promisoria para el manejo de los lepidópteros plagas, ofreciendo la posibilidad de reducir el uso total de insecticidas químicos de amplio espectro con menos riesgo para la salud humana y el medio ambiente. Sin embargo el uso generalizado de los cultivos mejorados genéticamente pueden ocasionar resistencia por parte de los insectos plagas (Zhu et al., 2006).
Los efectos de las toxinas sobre los enemigos naturales non-target de las plagas han recibido poca atención, si bien están expuestos a los riesgos ya sea por la alimentación de los vegetales (por ejemplo, el polen) o de las presas que puedan contener el producto transgénico (Schuler et al., 1999). Las interacciones entre las plantas genéticamente mejoradas y los artrópodos enemigos naturales puede ser ventajosa, desventajosa o neutral (Van Emden, 1991).
Los resultados hallados en la duración del ciclo larval no se vieron influenciados cuando la alimentación larval fue con S. maydis, tanto criados sobre maíces Bt como convencional, siendo una relación neutral y en coincidencia con los obtenidos por Dutton et al. (2002) para el depredador Chysoperla carnea (Stephens), que no vio afectada la duración del ciclo larval al ser alimentado con áfidos, ya sea criados sobre maíces Bt o convencional. Esto pudo deberse a que los pulgones, considerados una de las principales presas para E. connexa, no ingieren la toxina debido a que la misma no es transportada por el floema (Lozzia et al., 1998; Raps et al., 2001). Así Dutton et al. (2002) concluyeron que solo se hallaron pequeñas cantidades de la toxina Cry1Ab, en análisis realizados a Rhopalosiphum padi L, mientras que en plagas que se alimentan de los tejidos completos, como en Tetranychus urticae Koch y S. littoralis (Boisduval), la actividad biológica de la toxina Cry1Ab se mantiene después de haber sido ingerida, reduciendo la calidad de la presa y afectando el desarrollo de las larvas del predador, como lo mencionaron Obrist et al. (2006) cuando estudiaron el efecto de presas criadas con material genéticamente mejorado sobre C. carnea.
Las plantas genéticamente mejoradas para lograr resistencia a insectos no causan, necesariamente efectos negativos, especialmente cuando herbívoros non-target son expuestos a ellos (Zhang et al., 2006a). Dogan et al. (1996) demostraron que Hippodamia convergens (Guering-Meneville) no vio afectado su supervivencia, consumo de áfidos, desarrollo o reproducción, cuando se alimentó de Myzus persicae (Sulzer) criados sobre plantas de patatas Bt, indicando que los pulgones no incorporan la toxina. A igual conclusión llegaron Dutton et al. (2002) con C. carnea después de alimentarse con R. padi y T. urticae criados con maíces que producen la proteína Cry1Ab. En el caso de los coccinélidos son capaces de acumular la toxina y pasarla a su progenie aunque las consecuencias de dicha transmisión no son aún conocidas (Zhang et al., 2006a).
Lo expresado explicaría los resultados hallados en el presente estudio en la duración de los distintos estadios larvales de E. connexa cuando el alimento fueron larvas de S. frugiperda, ya que presentó diferencias con respecto a la alimentación con áfidos, coincidiendo con los trabajos de Obrycki et al. (1989) y Klingen et al. (1996), para el depredador C. carnea el cual se desarrollo más lentamente y tuvo una mayor tasa de mortalidad cuando el alimento fueron larvas de lepidópteros comparado con otras presas. Además es de esperar que los predadores que poseen aparato bucal masticador, como los coccinélidos, incorporen la toxina al predar artrópodos alimentados sobre plantas Bt, porque consumen el intestino que es donde se localizan las toxinas (Romeis et al., 2006).
Respecto de los altos porcentajes de mortandad que se registraron, entre 30-32%, especialmente en el primer estadio larval cuando la dieta fueron larvas de lepidópteros, versus los áfidos (presas non-target de la toxina) coinciden con los hallados por Hilbeck et al. (1998b) quienes encontraron que en larvas de C. carnea aumentó al 62% cuando se utilizó como presa Ostrinia nubilalis (Hübner) criadas en maíz Cry1Ab versus S. littoralis, en que la mortalidad fue del 37%. Estudios de preferencia, demostraron que las larvas de primer estadio de Coleomegilla quadrifasciata (Mulsant) descartan las larvas de II estadio de Colias lesbia (F.) para su alimentación (Salto et al., 1986). La razón para ello pudo deberse a: 1) que no sean presas de alta calidad para cubrir las necesidades alimenticias del predador; 2) a que tienen la capacidad de escapar y defenderse cuando son atacadas por un predador pequeño (Dutton et al., 2002). Las plantas con el gen del Bt, normalmente poseen altos niveles de resistencia a plagas de los órdenes lepidópteros y coleópteros. Sin embargo un gran número de insectos que son controlados pueden sobrevivir, aunque son de menor tamaño y sus tasas de crecimiento y fecundidad pueden verse reducidas (Obrist et al., 2006). Esa disminución en la calidad del huésped puede afectar negativamente la sobrevivencia larval, el tamaño del adulto y la fecundidad de los parasitoides o predadores que los controlan en condiciones naturales (van Emden, 1990).
Es sabido que los coccinélidos adultos utilizan polen como recurso alimenticio en su dieta en condiciones naturales (Bertolaccini et al., 2008). Por lo tanto la fecundidad puede verse influenciada en forma significativa por la dieta larval, por lo que en éste estado de desarrollo la alimentación sería un factor limitante en la formación de huevos, no así en la viabilidad de los mismos. Los índices de reproducción para los tratamientos con polen difirieron significativamente entre sí, por lo que la toxina de Bt tendría efectos negativos. Estos resultados contrastan con los de Bai et al. (2005) cuando estudiaron la expresión de la toxina en polen del arroz sobre Propylea japonica (Thunberg). Para este mismo coccinélido Zhang et al. (2006b) concluyeron que no vio afectado la supervivencia y el desarrollo larval, la mortalidad pupal, la fecundidad y la longevidad del adulto.
Podemos concluir que E. connexa se vio más afectada por el tipo de presa que por la alimentación de éstas. Los pulgones son el alimento preferido por este coccinélido, sin embargo las larvas de lepidópteros, presas comunes en los cultivo de maíz y que ingieren la toxina pueden resultar letales o sub-letales para el predador, afectando el ciclo y la fecundidad.
Nuestro trabajo sugiere que las plagas target al efecto de las plantas genéticamente modificadas pueden tener efectos negativos sobre los predadores, mientras que los áfidos, al no ser directamente influenciados cuando se crían sobre maíces trangénicos, no tienen efectos directos sobre E. connexa, aportando, por lo tanto, conclusiones importantes para ser tenidas en cuenta en la evaluación de los análisis de riesgos potenciales para el ambiente del uso de los maíces genéticamente mejorados. Sin embargo, los estudios a campo y en laboratorio deben continuar a fin de esclarecer la importancia de las plantas genéticamente mejoradas a los tres niveles tróficos.
Agradecimientos
A la Universidad Nacional del Litoral (Argentina) por la subvención de las investigaciones, a través de los Proyectos CAI+D.
Referencias Bibliográficas
Artigas, J.N. (1994) - Entomología económica. Insectos de interés agrícola, forestal, médico y veterinario (nativos, introducidos y susceptibles de ser introducidos). Vol 2. Concepción, Chile, Eds. Universidad de Concepción , 943 p. [ Links ]
Bai, Y.Y.; Jiang, M.X. y Cheng, J.A. (2005) - Effects of transgenic cry1Ab rice pollen on fitness of Propylea japonica (Thunberg). Journal of Pest Science, 78, 3: 123-128. [ Links ]
Bertolaccini, I.; Núñez-Pérez E. y Tizado, E.J. (2008) - Effect of wild flowers on oviposition of Hippodamia variegata (Coleoptera: Coccinellidae) in laboratory. Journal of Economic Entomology, 101, 6: 1792-1796. [ Links ]
Bonnemaison, L. (1980) - Wheat principal animal pest. Wheat Document Ciba-Geigy Technical Monograf. Ciba-Geigy Corporation, 95 p. [ Links ]
Delfino, M.A. (2002) - Dos especies de pulgones de interés fitosanitario nuevas para la afidofauna argentina (Hemiptera: Aphididae). In: Resúmenes XI Jornadas Fitosanitarias Argentinas. Córdoba, Argentina, Universidad Nacional de Río Cuarto, Facultad de Agronomía y Veterinaria, p.123. [ Links ]
Dicke, F.F. y Guthrie, W.D. (1988) - The most important corn insects. In: Sprague, G.F. y Dudley, J.W. (Eds.) - Corn and corn improvement. Madison, Wisconsin, USA, ASA, CSSA, and SSSA Publishers, p. 767-867. [ Links ]
Dogan, E.B.; Berry, R.E; Reed, G.L. y Rossignol, P.A. (1996) - Biological parameters of convergent lady beetle (Coleoptera: Coccinellidae) feeding on aphids (Homoptera: Aphididae) on transgenic potato. Journal of Economic Entomology, 89, 5: 1105-1108. [ Links ]
Dutton, A.; Klein, H.; Romeis, J. y Bigler, F. (2002) - Uptake of Bt-toxin by herbivores feeding on transgenic maize and consequences for the predator Chrysoperla carnea. Ecological Entomology, 27, 4: 441-447. [ Links ]
Dutton, A.; Romeis, J. y Bigler, F. (2003) - Assessing the risks of insect resistant transgenic plants on entomophagous arthropods: Bt-maize expressing Cry1Ab as a case study. Biocontrol, 48, 6: 611-636. [ Links ]
Hilbeck, A.; Baumgartner, M.; Fried, P.M. y Bigler, F. (1998a) - Effects of transgenic Bacillus thuringiensis corn-fed prey on mortality and development time of inmature Chrysoperla carnea (Neuroptera: Crysopidae). Environmental Entomology, 27, 2: 480-487. [ Links ]
Hilbeck, A.; Moar, W.J.; Pusztai Carey, M.; Filippini, A. y Bigler, F. (1998b) - Toxicity of Bacillus thuringiensis Cry1Ab toxin to the predator Chrysoperla carnea (Neuroptera: Crysopidae). Environmental Entomology, 27, 5: 1255-1263. [ Links ]
Hodek, I. (1973) - Biology of coccinellidae. The Hague, Neederlands, Dr. W. Junk N.V. Publishers, 254 p. [ Links ]
Jervis, M. y Copland, M. (1996) - The live cycle. In: Jervis, M. y Kidd, N. (Eds.) - Insect natural enemies: practical approaches to their study and evaluation. London, UK, Chapman and Hall, p. 62-161. [ Links ]
Klingen, I.; Johansen, N. S. y T. Hofsvang. 1996. The predation of Chrysoperla carnea (Steph.) (Neuroptera: Chrysopidae) on eggs and larvae of Mamestra brassicae (Lepidoptera: Noctuidae). Journal of Applied Entomology Norway, 120: 363-367. [ Links ]
Lozzia, G.; Furlanis, M.; Manachini, B. y Rigamonti, I. (1998) - Effects of Bt corn on Rhopalosiphum padi L. (Rhynchota, Aphididae) and on its predators Chrysoperla carnea (Neuroptera, Chrysopidae). Bollettino di Zoologia Agraria e Bachicoltura, 30, 2:153-164. [ Links ]
Lundgren, J.G. y Wiedenmann, R.N. (2005) - Tritrophic interactions among Bt (Cry3Bb1) corn, aphid prey, and the predator Coleomegilla maculata (Coleoptera: Coccinellidae). Environmental Entomology, 34, 6: 1621-1625. [ Links ]
Obrist, L.B.; Dutton, A.; Romeis, J. y Bigler, F. (2006) - Biological activity of Cry1Ab toxin expressed by Bt maize following ingestion by herbivorous arthropods and exposure of the predator Chrysoperla carnea. BioControl, 51, 1:31-48. [ Links ]
Obrycki, J. J.; Hamid, M. N. y S. A. Sajap. 1989. Suitability of corn insect pests for development and survival of Chrysoperla carnea and Chrysopa oculata (Neuroptera: Chrysopidae). Environmental Entomology, 18: 1126-1130. [ Links ]
Ortego, J. y Difabio, M.E. (2002) - Primer registro de Sipha (Rungsia) maydis Passerini 1860 (Hemiptera: Aphididae) potencial plaga de cereales en Argentina. En: XI Jornadas Fitosanitarias Argentinas. Córdoba, Argentina, Universidad Nacional de Río Cuarto, Fac. de Agronomía y Veterinaria, p.126. [ Links ]
Peairs, F. y Saunders, J. (1981) - Plant damage and yield response to Diatraea saccharalis and Spodoptera frugiperda in selection cycles of two tropical maize populations in Mexico. Turrialba, 31, 1: 55-62. [ Links ]
Pilcher, C.D.; Rice, M.E. y Obrycki, J.J. (2005) - Impact of transgenic Bacillus thuringiensis corn and crop phenology on five nontarget arthropods. Environmental Entomology, 34, 5: 1302-1316. [ Links ]
Ríos de Saluso, M.L.A. (1989) - Nueva plaga para los sorgos graníferos de la zona. Paraná, Instituto Nacional de Tecnología Agropecuaria-Estación Experimental Agropecuaria Paraná, 2 p. (Información Técnica 1). [ Links ]
Raps A.; J. Kehr y P. Gugerli. 2001. Immunological analysis of phloem sap of Bacillus thuringiensis corn and of the non-target herbivore Rhopalosiphumm padi (Homoptera: Aphididae) for the presence of Cry1Ab. Mol Ecol. 10: 525-533. [ Links ]
Romeis, J.; Meissle, M. y Bigler, F. (2006) - Transgenic crops expressing Bacillus thuringiensis toxins and biological control. Nature biotechnology, 24, 1: 63-71. [ Links ]
Salto, C.E., Berra, I.A.; Imwinkelried, J.M. y Frana, J.E. (1986) - Preferencia en la alimentación de: II Coleomegilla quadrifasciata var. octodecimpustulata Mulsant (Coleoptera, Coccinellidae). Rafaela, Instituto Nacional de Tecnología Agropecuaria-Estación Experimental Agropecuaria Paraná, 12 p. (Informe Técnico 21). [ Links ]
Salto, C.; Bertolaccini, I.; Imwinkelried, J. M. y Frana, J. (1990) - Parámetros biológicos e influencia de distintos niveles alimenticios en el desarrollo de Eriopis connexa (Coleoptera, Coccinellidae). Rafaela, Instituto Nacional de Tecnología Agropecuaria-Estación Experimental Agropecuaria Paraná, 12 p. (Publicación Técnica 50). [ Links ]
Schuler, T.H.; Poppy, G.M.; Kerry, B.R. y Denholm, I. (1999) - Potential side effects of insect-resistant transgenic plants on arthropod natural enemies. Trends in Biotechnology, 17, 5: 210-216. [ Links ]
van Emden, H.F. (1990) - The interaction of host plant resistance to insects with other control measures. In: Proceedings of the Brighton Crop Protection Conference on Pests and Diseases. Brighton, UK, British Crop Protection Council, p. 939-948. [ Links ]
van Emden, H.F. (1991) - The role of host plant resistance in insect pest mismanagement. Bulletin of Entomological Research 81, 2: 123-126. [ Links ]
Viana, P.A.; Waquil, J.M.; Valicente, F. y Cruz, I. (2004) - Ocorrência e controle de pragas na safrinha de milho nas regiões Norte e Oeste do Paraná. Embrapa. 8 p. (Circular Técnica 45). [ Links ]
Zhang, G-F.; Wan, F-H.; Lövei, G.L.; Liu, W-X. y Guo, J-Y. (2006a) - Transmission of Bt toxin to the predator Propylaea japonica (Coleoptera: Coccinellidae) through its aphid prey feeding on transgenic Bt cotton. Environmental Entomology, 35, 1: 143-150. [ Links ]
Zhang, S.Y.; Li, D-M.; Cui, J. y Xie, B-Y. (2006b) - Effects of Bt-toxin Cry1Ac on Propylaea japonica Thunberg (Col., Coccinellidae) by feeding on Bt-treated Bt-resistant Helicoverpa armigera (Hübner) (Lep., Noctuidae) larvae. Journal of Applied Entomology, 130, 4: 206-212. [ Links ]
Zhu, S.R.; Su, J.W.; Liu, X.H.; Du, L.; Yardim E.N. y Ge, F. (2006) - Development and reproduction of Propylaea japonica (Coleoptera: Coccinellidae) raised on Aphis gossypii (Homoptera: Aphididae) fed transgenic cotton. Zoological Studies, 45, 1: 98-103. [ Links ]
Recebido/Received: 2012.06.19
Aceitação/Accepted: 2013.01.25

