










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias vol.41 no.4 Lisboa dez. 2018
https://doi.org/10.19084/RCA18126
ARTIGO
Morfologia de sementes e plântulas de Handroanthus impetiginosus (Mart. ex DC.) Mattos
Morphology of seeds and seedlings of Handroanthus impetiginosus (Mart. ex DC.) Mattos
Francival C. Felix, Josenilda A. D. de Medeiros e Mauro V. Pacheco*
Unidade Acadêmica Especializada em Ciências Agrárias, Universidade Federal do Rio Grande do Norte, RN 160 - Km 03, Distrito de Jundiaí, CEP 59280-000, Macaíba, Rio Grande do Norte, Brasil
(*E-mail: pachecomv@hotmail.com)
RESUMO
Estudos morfológicos com sementes e plântulas de espécies florestais são fundamentais para a compreensão da dinâmica florestal e interpretação de resultados dos testes de germinação. Assim, o objetivo deste trabalho consistiu em descrever a morfologia de sementes e plântulas de Handroanthus impetiginosus (Bignoniaceae). Para isso, fez-se a biometria e descrição morfológica de sementes com e sem asas, bem como a morfometria e descrição morfológica de plântulas durante o processo germinativo. As sementes de H. impetiginosus pouco se diferenciam de outras espécies do género Handroanthus Mattos, mas apresentam diferenças quanto aos atributos biométricos. Os aspectos morfológicos da plântula de H. impetiginosus permitem distingui-la de outras espécies do género. A germinação de semente de H. impetiginosus é epígea foliáceo fanerocotiledonar, com o início da formação de plântulas normais ao décimo terceiro dia após a semeadura.
Palavras-chave: Biometria de sementes, germinação, morfometria de plântulas, ipê-roxo, plântula.
ABSTRACT
Morphological studies with seeds and seedlings of forest species are fundamental for the understanding of forest dynamics and interpretation of results of germination tests. Thus, the objective of this work was to describe the morphology of seeds and seedlings of Handroanthus impetiginosus (Bignoniaceae). For this, the biometry and morphological description of seeds with and without wings were made, as well as the morphometry and morphological description of seedlings during the germination process. Seeds of H. impetiginosus differ little from some species of the genus Handroanthus Mattos, but they do differ in the biometric aspects. The morphological characteristics of the seedling of H. impetiginosus allow its distinction of other species of the genus. Seed germination of H. impetiginosus is phytogean foliar epidermis, with the beginning of normal seedling formation on the thirteenth day after sowing.
Keywords: Seed biometry, germination, seedling morphometry, ipê-roxo, seedling.
INTRODUÇÃO
Handroanthus impetiginosus (Mart. ex DC.) Mattos (Bignoniaceae) é uma espécie arbórea nativa do Brasil que pode atingir de 20 a 35 m de altura, a qual é conhecida principalmente como ipê-roxo, pau-darco-roxo e ipê-roxo-da-mata (Martins et al., 2011; Silva et al., 2015). A sua madeira é utilizada na indústria de móveis (marcenaria) de luxo e a casca apresenta propriedades farmacológicas (Lima et al., 2014), no entanto o aspecto visual da copa proporciona efeito de rara beleza durante a abundante floração, destacando-se em projetos paisagísticos (Gemaque et al., 2002).
Os aspectos morfológicos da germinação das sementes são importantes para a compreensão dos mecanismos de propagação da espécie, interpretação e padronização de testes de germinação, e também na identificação de plântulas em campo (Abud et al., 2010), uma vez que o reconhecimento de espécies no estádio juvenil é completamente diferente da planta adulta devido às diferenças morfológicas existentes entre as mesmas (Amorim et al., 2008).
Estudos morfológicos de plântulas pouco descrevem seus órgãos estruturais e adaptações que se relacionam com o ambiente de ocorrência da espécie (Oliveira e Paoli, 2016), e por isso, ainda faltam estudos direcionados para a morfologia de sementes (Freitas et al., 2013; Silva et al., 2014) e plântulas de espécies florestais (Silva et al., 2008).
Atualmente, ainda há dificuldade na identificação e distinção de espécies do género Handroanthus Mattos (anteriormente classificado como Tabebuia Gomes ex A.P. de Candolle) da família Bignoniaceae, o qual apresenta grande similaridade morfológica entre sementes e plântulas (Oliveira et al., 2006).
Assim, trabalhos focados na descrição interna e externa de sementes e morfologia de plântulas visando a identificação de espécies e grupos taxinómicos possuem importância especial (Barretto e Ferreira, 2011; Pinto et al., 2016; Rios et al., 2016), de modo a que seja possível distinguir espécies de géneros e famílias botanicamente próximas (Silva et al., 2008), além de ser possível fazer o reconhecimento de espécies em banco de sementes ou em banco de plântulas, bem como reconhecer as estruturas morfológicas de espécies florestais em trabalhos de silvicultura (Duarte et al., 2016). Além disso, é possível também inferir sobre a estratégia de propagação das sementes e desenvolvimento de plântulas em condições naturais de floresta (Guerra et al., 2006), as quais muitas vezes estão em situação de elevada pressão antrópica, principalmente relacionados as espécies nativas (Oliveira et al., 2012).
Face ao exposto, o objetivo deste trabalho consistiu em descrever a morfologia de sementes e plântulas de H. impetiginosus.
MATERIAL E MÉTODOS
O lote de sementes de H. impetiginosus utilizado foi coletado em 2015, em árvores localizadas no campus central da Universidade Federal do Rio Grande do Norte, e mantidos em Banco de Sementes (17 ± 2 °C e 40 ± 10% de umidade relativa do ar) da Floresta Nacional de Nísia Floresta, Nísia Floresta/RN, Brasil.
Inicialmente, realizou-se a avaliação biométrica (comprimento, largura e espessura) de 100 sementes com e sem asas, bem como o peso de mil sementes (PMS) e grau de umidade das sementes (Brasil, 2009). Para dimensões e PMS, utilizou-se respectivamente, paquímetro digital (0,01 cm) e balança analítica de precisão (0,001 g), sendo os resultados da biometria de sementes expressos em centímetros (cm) para média, desvio padrão, mínimo, máximo, mediana e moda para cada dimensão de semente, além de assimetria e curtose.
Em seguida, fez-se a análise da morfologia interna (hidratação em água destilada por 24-h) e externa (sem intumescimento) da semente (Barroso et al., 1999), avaliando-se os aspectos de coloração do tegumento, forma e características do tegumento e do embrião.
Para a avaliação do processo germinativo, procedeu-se à instalação de testes de germinação em substrato de areia e papel, com sementes desprovidas de asas e desinfestadas em solução de hipoclorito de sódio a 2,5% durante 5-min, e semeadas com a posição do hilo voltada para baixo. Em areia, realizou-se a semeadura de 100 sementes a 1,0 cm de profundidade em bandejas plásticas preenchidas com areia esterilizada e umedecida com 60% da capacidade de retenção de água; outras 100 sementes foram distribuídas em papel toalha (Germitest®) umedecido com 2,5 vezes o seu peso seco, organizados na forma de rolos e colocados em sacos plásticos transparentes. Os rolos de papel e as bandejas plásticas foram mantidos em câmara de germinação regulada à temperatura de 25 °C (Brasil, 2013), com fotoperíodo de 8-h, durante 33 dias.
Procedeu-se diariamente a avaliação do processo germinativo e dos aspectos morfológicos de plântulas até à formação do segundo par de eófilos (o primeiro par de folhas com limbo) (Gonçalves e Lorenzi, 2007), descrevendo o tipo de germinação (Miquel, 1987), formação de plântulas normais por meio do substrato areia (Brasil, 2013) e emissão de raiz primária por meio o substrato papel.
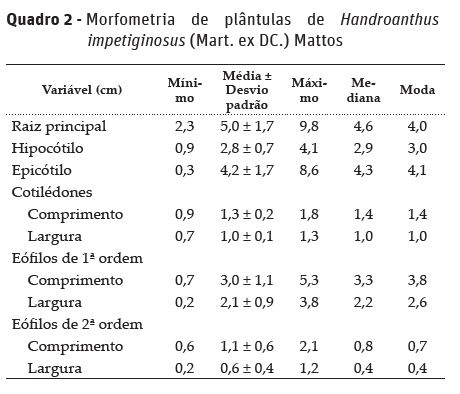
No final do período de germinação, realizou-se a morfometria da raiz principal, hipocótilo, epicótilo, cotilédones, eófilos de primeira e segunda ordem das plântulas normais (Brasil, 2013), cuja medição foi feita com paquímetro digital (0,01 cm) e os resultados foram expressos em centímetros (cm) para média, desvio padrão, mínimo, máximo, mediana e moda para cada morfometria da plântula.
RESULTADOS E DISCUSSÃO
A formação de sementes em espermatófitas envolve uma série de eventos citológicos, bioquímicos e fisiológicos a partir da fecundação do óvulo que culmina com a formação do embrião e do endosperma, o primeiro é composto por eixo hipocótilo-radícula e um ou mais cotilédones, enquanto que o segundo pode ser degradado durante a embriogênese ou permanecer até o momento da maturidade fisiológica da semente (Bewley et al., 2013).
Sementes de H. impetiginosus são ortodoxas (Martins et al., 2011) e apresentam formato comprimido (ou estreito) com ligeira curvatura no seu núcleo seminífero, expandidas em asas duplas laterais assimétricas e hialinas do tipo membranáceo (Figura 1), concordando com a morfologia externa de sementes de espécies circunscritas na tribo Tecomeae (Barroso et al., 1999). O tegumento possui coloração creme sem brilho e o pleurograma fechado é visível em ambos os lados da semente, não sendo possível diferenciar testa de tegma (Figura 1). As asas presentes na semente favorecem a dispersão pelo vento (anemocoria) a longas distâncias (Oliveira et al., 2006) e o peso relativamente leve facilita o transporte, embora o estabelecimento em campo seja prejudicado por possuir menos reservas (Oliveira et al., 2008).
A rafe enegrecida é longitudinal à semente, o hilo é visível, linear na base da semente e de coloração castanho escuro, o micrópilo possui formato puntiforme (Figuras 2A e B). O endosperma é escasso e rodeia completamente o embrião maduro, o qual possui dois cotilédones delgados e foliáceos (Figura 2C). O embrião é classificado em axial do tipo espatulado de acordo com a forma, tamanho e posição que o mesmo ocupa na semente. O eixo hipocótilo-radícula é pouco visível a olho nu, possui forma reta, a plúmula é conspícua e a radícula, glabra (Figura 2D). Características morfológicas do embrião da semente desta espécie não permitiram verificar diferenças marcantes dentro do género Handroanthus Mattos (Oliveira et al., 2006).
O peso de mil sementes para H. impetiginosus com asas e teor de água de 7,5% foi de 104,3 g, equivalente a aproximadamente 9600 sementes por quilograma, ou seja, dentro do esperado para a espécie que é de 8 a 35 mil sementes por quilograma (Brasil, 2013). Nas espécies florestais há grande diversidade quanto aos caracteres biométricos das sementes o que explica essa ampla variação, como verificado em estudo anterior com esta espécie, em que se obteve um PMS equivalente a 68,4 g em teor de água de 8,6% (Maeda e Matthes, 1984). E também, a não padronização do grau de umidade das sementes no momento da realização do PMS é outro fator que causa variações ainda mais expressivas.
Para os resultados biométricos, constatou-se que as sementes com asas apresentaram 3,9 ± 0,7 cm de comprimento, 1,2 ± 0,2 cm de largura e 0,2 ± 0,1 cm de espessura, enquanto que sementes sem asas possuíam 1,6 ± 0,3 cm de comprimento, 1,1 ± 0,2 cm de largura e 0,2 ± 0,1 de espessura (Quadro 1). Resultados que evidenciam a existência de diferenças biométricas dentro do género quando comparado à espécie Handroanthus aureus Mattos, que apresenta sementes relativamente maiores com (5,8 x 2,1 x 0,3 cm) e sem (1,7 x 1,3 x 0,2 cm) asas e PMS equivalente a 263,6 g (Oliveira et al., 2006). Na espécie Handroanthus chrysotrichus (Mart. ex A. DC.) Mattos foram observadas dimensões relativamente menores, uma vez que as sementes com e sem asas apresentaram, respectivamente, 2,8 x 0,7 x 0,1 cm e 0,6 x 0,5 x 0,1 cm, com PMS equivalente a 4,0 g (Oliveira et al., 2008).
A média e a mediana das sementes com e sem asa são iguais e com baixo desvio padrão, demostrando homogeneidade da amostra. À exceção da largura, a assimetria e a curtose das demais variáveis apresentam valores predominantemente próximos a zero, indicando proximidade à distribuição normal (Zuffo et al., 2014). Em sementes aladas a assimetria é negativa enquanto que sementes sem asas apresentam assimetria positiva. A largura das sementes com asas bem como largura e espessura das sementes sem asas apresentam curtose leptocúrtica e demais variáveis são platicúrticas.
O processo germinativo inicia-se com a retomada do desenvolvimento do embrião após a absorção de água, culminando com emissão da raiz primária rompendo o tegumento da semente (Bewley et al., 2013). No entanto, considera-se semente germinada em termos tecnológicos, a que produz uma plântula com todas as estruturas básicas para a formação de uma planta autossuficiente estruturalmente em campo, com a presença de raiz, hipocótilo, epicótilo, cotilédones e gema apical (Brasil, 2013).
O intumescimento das sementes caracterizado pela mudança de coloração do tegumento e aumento do tamanho ocorreu entre o primeiro e segundo dia após a semeadura (Figura 3A), com emissão da raiz primária através da região hilar a partir do terceiro dia (Figura 3B), alongamento do hipocótilo no quinto dia (Figura 3C), e início da formação de plântulas normais contendo gemas apicais e eófilos em expansão no décimo terceiro dia após a semeadura (Figura 3D).
A germinação é considerada epígea foliáceo fanerocotiledonar (Miquel, 1987), em que o hipocótilo ressurge acima no nível do substrato e o tegumento se desprende dos cotilédones, também foi observado que a areia auxilia na remoção do tegumento nos cotilédone da plântula em desenvolvimento. Os resultados condizem com o fato de que a germinação epígea fanerocotiledonar é comumente mais frequente entre as espécies arbóreas do Brasil (Lobo et al., 2014).
O tipo de germinação também se assemelha ao das espécies H. avellanedae Lor. ex Griseb, H. chrysotrichus (Souza e Oliveira, 2004), H. aureus e H. roseo-albus (Ridl.) Mattos (Lobo et al., 2014), mas diferencia-se da espécie H. serratifolius (Vahl) S.O. Grose, a qual possui plântula epígea criptocotiledonar (Machado et al., 2002). Entretanto, para essas espécies não foi observado desenvolvimento de eófilos de segunda ordem em 30 dias após a semeadura, enquanto que para a espécie em estudo foi possível visualizar o início de desenvolvimento dessas estruturas aos 28 dias após a semeadura (Figura 3E).
Plântulas normais de H. impetiginosus apresentam raiz principal medindo 5,0 ± 1,7 cm, hipocótilo com 2,8 ± 0,7 cm, epicótilo com 4,2 ± 1,7 cm, e cotilédones com 1,3 ± 0,2 cm de comprimento e 1,0 ± 0,1 cm de largura. Os eófilos de primeira ordem possuem 3,0 ± 1,1 cm de comprimento e 2,1 ± 0,9 cm de largura, enquanto que os eófilos de segunda ordem apresentam 1,1 ± 0,6 cm de comprimento e 0,6 ± 0,4 cm de largura (Quadro 2).
A raiz principal é axial pivotante (Figura 3E) de coloração castanho escuro e apresenta pêlos absorventes de coloração esbranquiçada na zona pilífera (Figura 4D), a coifa é pontiaguda de coloração amarelo-marrom escura (Gonçalves e Lorenzi, 2007), visualizada somente com o auxílio de microscópio estereoscópio. A formação de raízes secundárias de coloração mais esbranquiçada ocorre a partir do décimo dia após a semeadura. A presença de pilosidade nas raízes é característica semelhante para as espécies do género Handroanthus.
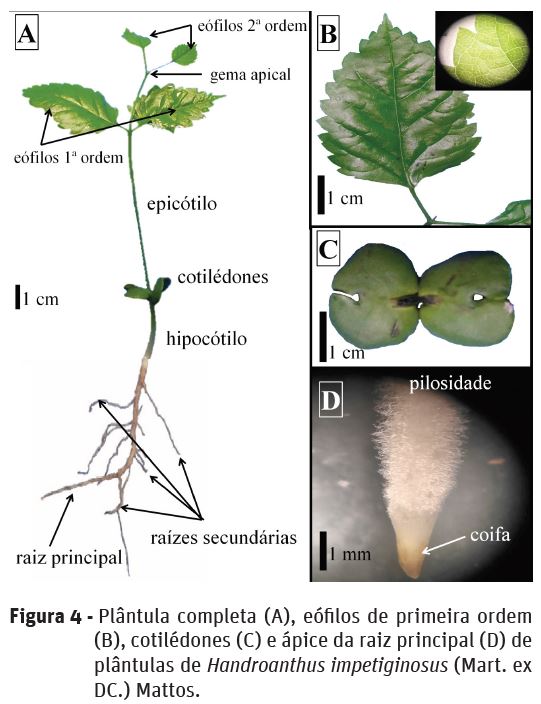
O hipocótilo inicia a diferenciação da raiz principal a partir do oitavo dia, o qual é esbranquiçado, tornando-se verde à medida que se aproxima dos cotilédones, enquanto que o epicótilo é totalmente verde. Os cotilédones são opostos, isófilos, levemente assimétricos e voltados para cima, são persistentes, foliáceos de coloração verde e sésseis (Figura 4C), diferentemente do encontrado para outras espécies do género (H. avellanedae e H. chrysotrichus) que possuem pecíolos evidentes e longos (Souza e Oliveira, 2004), evidenciando uma das poucas diferenças morfológicas entre as espécies na fase de plântula.
Os eófilos de primeira e segunda ordem são simples, peciolados, de filotaxia oposta, glabros e de tamanho semelhantes, iniciando a expansão dos eófilos de primeira e segunda ordem, respectivamente, ao 13 e 28º dia após a semeadura. Os eófilos possuem formato ovado, venação cladódroma, borda serreada, base arredondada e ápice agudo (Figura 4B). A ausência de pilosidade nos eófilos nesta espécie pode ser utilizada para a diferenciar da espécie H. chrysotrichus que apresenta eófilos com pilosidade (Souza e Oliveira, 2004).
Espécies de rápida germinação normalmente possuem plântulas epígeas fanerocotilédonares de folhas simples (Gogosz et al., 2015), da mesma forma como o observado para esta espécie na fase de plântula, indicando que H. impetiginosus possui a capacidade de rápido estabelecimento assim que é dispersa da planta mãe. Sabe-se também que a viabilidade das sementes desta espécie supera dois anos de armazenamento a baixa temperatura e reduzida percentagem de umidade (Martins et al., 2011), com boa taxa de germinação em temperaturas próximo à 30 °C sob a presença de luz (Oliveira et al., 2005).
Portanto, a espécie possui sementes ortodoxas que rapidamente germinam e possivelmente formam um banco de plântulas por baixo da copa, aguardando o momento ideal para assumirem a posição de dominantes na floresta, ou para se desenvolver prontamente sob a copa aberta em áreas que fornecem mais luminosidade.
CONCLUSÕES
Sementes de H. impetiginosus pouco se diferenciam de algumas espécies do género Handroanthus, mas apresentam diferenças quanto aos aspectos biométricos da semente. Os aspectos morfológicos da plântula de H. impetiginosus permitem a diferenciação de outras espécies do género, e a germinação de semente de H. impetiginosus é epígea foliáceo fanerocotiledonar, com o início da formação de plântulas normais ao décimo terceiro dia após a semeadura.
Referências Bibliográficas
Abud, H.F.; Gonçalves, N.R.; Reis, R.G.E.; Gallão, M.I. & Innecco, R. (2010) - Morphology of seed and seedling of safflower.Revista Ciência Agronômica, vol. 41, n. 2, p. 259-265. http://dx.doi.org/10.1590/S1806-66902010000200013 [ Links ]
Amorim, I.L.; Davide, A.C.; Ferreira, R.A. & Chaves, M.M.F. (2008) - Morphology of fruits, seeds and seedlings of Senna multijuga var. lindleyana (Gardner) H. S. Irwin & Barneby – Leguminosae Caesalpinioideae. Brazilian Journal of Botany, vol. 31, n. 3, p. 507-516. http://dx.doi.org/10.1590/S0100-84042008000300014 [ Links ]
Barretto, S.S.B. & Ferreira, R.A. (2011) - Morphological aspects of fruits, seeds and seedlings of the leguminosae mimosoideae species: Anadenanthera colubrina (Vellozo) Brenan e Enterolobium contortisiliquum (Vellozo) Morong.Journal of Seed Science, vol. 33, n. 2, p. 223-232. http://dx.doi.org/10.1590/S0101-31222011000200004 [ Links ]
Barroso, G.M.; Morim, M.P.; Peixoto, A.L. & Ichaso, C.L.F. (1999) - Frutos e sementes: morfologia aplicada à sistemática de dicotiledôneas. 1. Ed. Viçosa, MG: Editora/ UFV. 443 p. [ Links ]
Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M. & Nonogaki, H. (2013) - Seeds: physiology of development, germination and dormancy. 3. Ed, New York, NY: Springer, 392 p. [ Links ]
Brasil (2009) - Regras para análise de sementes. Ministério da Agricultura, Pecuária e Abastecimento. Secretaria de Defesa Agropecuária. Brasília: MAPA/ACS, 399 p.
Brasil (2013) - Instruções para a análise de sementes de espécies florestais. Ministério da Agricultura, Pecuária e Abastecimento. Secretaria de Defesa Agropecuária. Brasília: MAPA/ACS, 98 p.
Duarte, M.M.; Paula, S.R.P.; Ferreira, F.R.L. & Nogueira, A.C. (2016) - Morphological characterization of fruit, seed and seedling and germination of Hymenaea courbaril L. (Fabaceae) (Jatobá).Journal of Seed Science, vol. 38, n. 3, p. 204-211. http://dx.doi.org/10.1590/2317-1545v38n3159734 [ Links ]
Freitas, T.P.; Freitas, T.A.S.; Campos, B.M.; Fonseca, M.D.S. & Mendonça, A.V.R. (2013) - Morfologia e caracterização da germinação em função da posição das sementes no fruto de sabiá.Scientia Plena, vol. 9, n. 3, p. 1-9. [ Links ]
Gemaque, R.C.R.; Davide, A.C. & Faria, J.M.R. (2002) - Indicadores de maturidade fisiológica de sementes de ipê-roxo (Tabebuia impetiginosa (Mart.) Standl.).Cerne, vol. 8, n. 2, p. 84-91. [ Links ]
Gogosz, A.M.; Boerger, M.R.T.; Cosmo, N.L. & Nogueira, A.C. (2015) - Morphology of diaspore and seedling of tree species of the Araucaria forest (Subtropical Ombrophilous Forest) in southern Brazil.Floresta, vol. 45, n. 4, p. 819-832. http://dx.doi.org/10.5380/rf.v45i4.35017 [ Links ]
Gonçalves, E.G. & Lorenzi, H. (2007) - Morfologia vegetal: organografia e dicionário ilustrado de morfologia das plantas vasculares. São Paulo: Instituto Plantarum de Estudos da Flora. [ Links ]
Guerra, M.E.C.; Medeiros Filho, S. & Gallão, M.I. (2006) - Morfologia de sementes, de plântulas e da germinação de Copaifera langsdorfii Desf. (Leguminosae-Caesalpinioideae).Cerne, vol. 12, n. 4, p. 322-328. [ Links ]
Lima, P.R.; Horbach, M.A; Dranski, J.A.L.; Ecco, M.; Malavasi, M.M. & Malavasi, U.C. (2014) - Avaliação Morfofisiológica em mudas de Handroanthus impetiginosus (Mart. ex DC.) Mattos durante a Rustificação. Floresta e Ambiente, vol. 21, n. 3, p. 316-326. http://dx.doi.org/10.1590/2179-8087.058813 [ Links ]
Lobo, G.A.; Santana, D.G.; Salomão, A.N.; Renbein, L.S. & Wielewicki, A.P. (2014) - A technological approach to the morphofunctional classification of seedlings of 50 Brazilian forest species.Journal of Seed Science, vol. 36, n. 1, p. 87-93. http://dx.doi.org/10.1590/S2317-15372014000100011 [ Links ]
Machado, C.F.; Oliveira; Davide, A.C. & Guimarães, R.M. (2002) - Methodology for performing germination test of (Tabebuia serratifolia (Vahl) Nicholson).Cerne, vol. 8, n. 2, p. 17-25. [ Links ]
Maeda, J.A. & Matthes, L.A.F. (1984) - Conservação de sementes de ipê.Bragantia, vol. 43, n. 1, p. 51-61. [ Links ]
Martins, L.; Lago, A.A. & Cicero, S.M. (2011) - Physiological quality of Tabebuia avellanedae and Tabebuia impetiginosa seeds submitted to extreme drying.Journal of Seed Science, vol. 33, n. 4, p. 626-634. http://dx.doi.org/10.1590/S0101-31222011000400004 [ Links ]
Miquel, S. (1987) - Morphologie, fonctionnelle de plantules déspèces forestières du Gabon.Bulletin du Muséum National d'Histoire Naturelle, vol. 9, p. 101-121. [ Links ]
Oliveira, L.M.; Carvalho, M.L.M.; Silva, T.T.A. & Borges, D.I. (2005) - Temperatura e regime de luz na germinação de sementes de Tabebuia impetiginosa (Martius ex A. P. de Candolle) Standley e T. serratifolia Vahl Nich. – Bignoniaceae. Ciência e Agrotecnologia, vol. 29, n. 3, p. 642-648. http://dx.doi.org/10.1590/S1413-70542005000300020 [ Links ]
Oliveira, A.K.M.; Schleder, E.J.D. & Favero, S. (2006) - Morphological characterization, viability and vigor of Tabebuia aurea (Silva Manso) Benth. & Hook. f. ex. S. Moore seeds.Revista Árvore, vol. 30, n. 1, p. 25-32. http://dx.doi.org/10.1590/S0100-67622006000100004 [ Links ]
Oliveira, A.K.M.; Schleder, E.J.D. & Favero, S. (2008) - Morphological characterization, viability, and vigor of Tabebuia chrysotricha (Mart. ex. DC.) Standl seeds.Revista Árvore, vol. 32, n. 6, p. 1011-1018. http://dx.doi.org/10.1590/S0100-67622008000600006 [ Links ]
Oliveira, S.S.C.; Araújo Neto, J.C.; Cruz, S.J.S. & Ferreira, V.M. (2012) - Morphometric characterization of seeds and plantules and Anadenanthera macrocarpa (Benth.) Brenan germination.Ciência Florestal, vol. 22, n. 3, p. 643-653. http://dx.doi.org/10.5902/198050986630 [ Links ]
Oliveira, J.H.G. & Paoli, A.A.S. (2016) - Morphology and development of seedlings of Acalypha gracilis (Spreng.) Müll. Arg, Euphorbia cotinifolia L. and Jatropha gossypiifolia L. (Euphorbiaceae).Arnaldoa, vol. 23, n. 2, p. 443-460. http://doi.org/10.22497/arnaldoa.232.23204 [ Links ]
Pinto, M.B.; Grabias, J.; Hoffmann, P.M.; Velazco, S.J.E. & Blum, C.T. (2016) - Caracterização morfológica de frutos, sementes, plântulas e germinação de Oreopanax fulvus Marchal.Agrária, vol. 11, n. 2, p. 111-116. http://doi.org/10.5039/agraria.v11i2a5366 [ Links ]
Rios, P.A.F.; Araújo Neto, J.C.; Ferreira, V.M. & Neves, M.I.R.S. (2016) - Seed morphometry and germination of Aechmea costantinii (Mez) L. B. Sm. (Bromeliaceae).Revista Caatinga, vol. 29, n. 1, p. 85-93. http://dx.doi.org/10.1590/1983-21252016v29n110rc [ Links ]
Silva, K.B.; Alves, E.U.; Bruno, R.L.A.; Matos, V.P. & Gonçalves, E.P. (2008) - Morphology of fruits, seeds and seedlings of Erythrina velutina Willd., Leguminoseae – Papilionideae.Journal of Seed Science, vol. 30, n. 3, p. 104-114. http://dx.doi.org/10.1590/S0101-31222008000300014 [ Links ]
Silva, K.B.; Pinto, M.S.C. & Souza, N.A. (2014) - Morfometria de frutos e diásporos de Acacia farnesiana (L.) Willd.Revista Verde, vol. 9, n. 2, p. 76-82. [ Links ]
Silva, G.H.; Santos, R.V. & Lucena, R.J. (2015) - Seedlings production of Handroanthus impetiginosus (Mart. ex DC.) Mattos in substrate containing vermiculite co-product. Scientific Electronic Archives, vol. 8, n. 2, p. 22-28. [ Links ]
Souza, A. & Oliveira, J.H.G. (2004) - Morfologia e anatomia das plântulas de Tabebuia avellanedae Lor. ex Griseb e T. chrysotricha (Mart. ex Dc.) Standl. (Bignoniaceae).Acta Scientiarum - Biological Sciences, vol. 26, n. 2, p. 217-226. http://dx.doi.org/10.4025/actascibiolsci.v26i2.1649 [ Links ]
Zuffo, A.M.; Andrade, F.R. & Zuffo Júnior, J.M. (2014) - Caracterização biométrica de frutos e sementes de baru (Dipteryx alata Vog.) na região leste de Mato Grosso, Brasil.Revista de Ciências Agrárias, vol. 37, n. 4, p. 463-471. [ Links ]
Agradecimentos
O primeiro e segundo autores agradecem à Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) pela concessão de bolsas.
Recebido/received: 2018.04.25
Aceite/accepted: 2018.07.02

