









Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPortuguese Journal of Nephrology & Hypertension
versión impresa ISSN 0872-0169
Port J Nephrol Hypert vol.28 no.1 Lisboa mar. 2014
REVIEW ARTICLE
Is renalase a new regulator of renal dopaminergic activity?
Será a renalase um novo regulador da atividade renal dopaminérgica?
Janete Quelhas-Santos1
1 Nephrology and Infectious Diseases Research & Development Group, INEB, University of Porto, Portugal.
ABSTRACT
Dopamine (DA) of renal origin behaves as an endogenous natriuretic hormone decreasing tubular sodium reabsorption as a paracrine/autocrine substance and contributing to blood pressure control. The availability of renal DA is determined by the interplay between the rate of synthesis and the degree of degradation of the amine. The degradation of renal DA is well recognized to be mediated by two main enzymes – types A and B monoamine oxidases (MAO-A and MAO-B) and catechol-O-methyltransferase (COMT). In studies aimed at reducing the degradation of renal DA, both the selective and combined inhibition of MAO-A and COMT, did not change renal DA excretion or natriuresis, notwithstanding the marked decrease observed in the urinary excretion of the corresponding deaminated and methylated metabolites. This observation suggested that other metabolic pathways in addition to COMT and MAO might be involved in the regulation of renal dopamine output.
Renalase, a new kidney-secreted catecholamines-degrading enzyme, is endowed with potent hypotensive effects when administered in vivo and may represent a new player in the regulation of the renal dopamine system. Key-Words: Chronic kidney disease; natriuresis; renal dopamine system; renalase.
RESUMO
A dopamina (DA) de síntese renal funciona como uma hormona natriurética endógena, que diminui a reabsorção tubular de sódio e contribui assim para o controlo da pressão arterial. A disponibilidade da DA de síntese renal é determinada pelo balanço entre a sua síntese e a sua degradação local. A degradação da DA de síntese renal é reconhecidamente assegurada por duas enzimas presentes nos tecidos renais – os tipos A e B de mono-aminoxídase (MAO-A e MAO-B) e a catecol-O-metiltransferase (COMT). Em estudos que procuraram reduzir a degradação da DA de síntese renal através da inibição seletiva e/ou combinada da MAO-A e da COMT, não se verificaram modificações na excreção urinária de DA nem na natriurese, apesar de se terem observado reduções marcadas na excreção urinária dos correspondentes metabolitos da DA, desaminados e metilados. Estes achados sugeriram a existência de vias metabólicas alternativas a nível renal para além da MAO e da COMT, com envolvimento potencial na regulação dos níveis urinários de DA. A renalase, uma enzima recentemente descrita que é segregada pelo rim e que foi implicada na metabolização de catecolaminas, exerce efeitos hipotensores quando é administrada in vivoPalavras-chave: Doença renal crónica; natriurese; renalase; sistema renal dopaminérgico.
RENAL DOPAMINERGIC SYSTEM
ACTIVITYThe kidney plays an important role in the regulation of blood pressure, nutrient and electrolyte reabsorption, as well as drug and metabolite excretion.
Because the kidney is important in the long-term regulation of blood pressure and is the principal organ involved in the regulation of sodium homoeostasis, many studies have focused on the abnormal renal handling of sodium in the pathogenesis of hypertension. Several studies have shown that human essential hypertension is associated with increased sodium transport in the renal proximal tubule1.
Dopamine (DA) of renal origin behaves as an endogenous natriuretic hormone decreasing tubular sodium reabsorption as a paracrine/autocrine substance and controlling blood pressure2,3.
At least 90% of urinary free DA has its origin in the kidney4, being mainly synthesized in renal proximal tubular cells through the decarboxylation of circulating or filtered L-3,4-dihydroxyphenylalanine (L-DOPA) by aromatic L-amino acid decarboxylase (AADC)5. The rate-limiting step of renal DA synthesis was suggested to be tubular uptake of L-DOPA and this was found to be an active transport process5.
The candidate transport systems for L-DOPA in renal proximal tubular cells, include sodium-dependent B(0), B(0,+), and y(+)L and sodium-independent systems L (LAT1 and LAT2) and b(0,+)6.
Dopamine of renal origin is also well recognized to undergo extensive deamination to 3,4-dihydroxyphenylacetic acid (DOPAC), O-methylation to 3-metoytyramine (3-MT) and deamination plus O-methylation to homovanillic acid (HVA) and the high levels of metabolic enzymes, such as types A and B monoamine oxidases (MAO-A and MAO-B) and catechol-O–methyltransferase (COMT) have been considered important determinants in the overall availability of renal DA2,7. In rat renal cortical slices incubated with L-DOPA, deamination by MAO was found to be the major metabolic pathway for renal DA7.
Dopamine excreted in urine is mainly derived from intrarenally produced DA. Because the daily urinary excretion of both DOPAC and HVA is several fold that of the parent amine, these two amine metabolites are used as useful parameters for assessment of the renal dopaminergic system activity, as well as for assessment of the activities of the corresponding enzymes8.
In mammals, the effects of DA are mediated by two DA receptor subtypes, D1-like (D1 and D5) and D2-like (D2, D3 and D4) receptors, which differ in their primary structures showing distinct affinities for DA receptor agonists and antagonists9-10. All DA receptor subtypes have been shown to regulate, directly or indirectly, sodium transport in the proximal and distal nephron and blood pressure9,11.
The mechanisms through which renal DA is thought to produce natriuresis involve mainly the inhibition of both apical (NHE exchange, Cl-/HCO3- exchange and Na+-Pi cotransport) and basolateral (Na+/K+-ATPase and Na+-HCO3- cotransport) transporters12,13.
The D1-like receptors couple to Gs-proteins and activate the adenylate cyclase-cAMP- protein kinase A signaling pathway14. In the kidney and other tissues, D1-like receptors can also couple to Gq/11 and activate the phospholipase C-diacylglycerol-protein kinase C pathway2,14.
During moderate sodium intake, renal DA is responsible for ~50% of the sodium excretion3,15 and several clinical observations, as well as studies on animals with various forms of genetic hypertension, suggested that defects in the renal dopaminergic system may contribute to the development of hypertension, by causing sodium retention9. Renal LAT2 is overexpressed in the prehypertensive spontaneously hypertensive rat (SHR), which might contribute to enhanced L-DOPA uptake in the proximal tubule and increased DA production, as an attempt to overcome the defect in D1 receptor function16.
The effects of renal dopaminergic regulation on sodium handling have been found to be more pronounced during high salt intake17-19 or volume expansion20,21.
In wild-type rats with normal renal function, high salt intake or volume expansion induced by isotonic saline infusion has been shown to increase the production of renal DA, as evidenced by increases in the urinary excretion of DA and DA metabolites17,20, tubular L-DOPA uptake22 and AADC activity19, with concomitant increases in natriuresis and diuresis23,24.
On the basis of the previous considerations, renal dopaminergic system seems essential for the maintenance of sodium homeostasis, and deficiencies in renal dopaminergic activity may be of particular relevance in the pathogenesis of conditions characterized by disturbances in renal sodium handling.
Renal dopaminergic system in Chronic Kidney Disease
Patients suffering from chronic kidney disease (CKD) present a reduced renal DA output evidenced by decreased urinary excretion of DA and metabolites, the extent of which is related to the degree of renal failure25.
The renal dopaminergic system activity was also found compromised in ¾nx rats in a time-dependent manner after renal mass ablation, suggesting that this may contribute to compromise sodium excretion and increase blood pressure in this CKD rat model26.
On the other hand, plasma norepinephrine levels are increased in CKD and this can be accounted for by both heightened sympathetic nerve activity and reduced catecholamine clearance27. Given that norepinephrine and renal DA exert antagonist effects on tubular sodium reabsorption, it was suggested that heightened sympathetic activity and the accompanied reduced renal dopaminergic tonus in CKD may contribute both to compromise sodium excretion and increase blood pressure in this population28,29. The participation of disturbances in sympathetic and renal dopaminergic activities in the pathophysiology of hypertension in CKD has become more complex with the recent discovery of renalase, a flavin adenine dinucleotide (FAD) -dependent oxidase potentially involved in the inactivation of catecholamines and in the regulation of blood pressure and cardiac function30.
RENALASE
Structure and characterization
In 2005, Desir and co-workers reported the discovery of a new enzyme called renalase, also termed MAO-C30. According to information available in Gen-Bank, the human renalase gene (RNLS, C10orf59) is located on chromosome 10 at q23.31, contains 9 exons spanning 311,000 base pairs (NC_000010.10) and encodes four alternatively spliced isoforms (hRenalase 1-4). The longer mRNA variant encodes a protein with 342 amino acid (aa) residues (NM_001031709.2) with a predicted molecular mass of 37.85 kDa, whereas the shorter variant Mrna encodes a 315-aa protein (NM _0018363.3) with a predicted molecular mass of 34.95 kDa.
Primary analysis of RNLS gene expression in human tissues has shown preferential expression in kidney, but also in heart, intestine, liver and skeletal muscle30.
Further studies also revealed renalase expression in peripheral nerves, adrenal glands, and the central nervous system, as well as in human adipose tissues and in rat embryos31-34. A recent study showed that renalase is also highly expressed in the reproductive system, suggesting its function in maturation of germ cells and steroid hormone regulation.
Renalase was detected in both plasma and urine of healthy individuals, thus suggesting that differently from other MAOs, renalase is secreted into the circulation30.
Only hRenalase1 was detected in human blood samples, which suggests that hRenalase 2-4 probably differ in function from hRenalase 132,35. The splice variants of renalase transcript appear to be tissue-specific and point to be a fine-tuning of renalase function.
In the deduced amino acid sequence of RNLS, a N-terminal signal peptide (residues 1-17), typical for secretory proteins, a dinucleotide-binding site (residues 3-42) and an amine oxidase domain (including residues 75-335), have been recognized30. The general fold of renalase classifies it as a member of the flavoprotein superfamily. Despite being structurally similar to MAO-like enzymes, renalase is not a typical oxidase and its amino acid sequence significantly differs from MAO-A and B (EC 1.4.3.4), sharing less than 14% of amino acid identity30. Renalase was thus classified as a new flavoprotein that functions as a FAD/NADH oxidase and degrades catecholamines with a distinct substrate specificity and inhibitor profile to that of MAO-A and MAO-B30.
Function and mechanism of action
Recombinant renalase was found to acutely lower blood pressure in a dose-dependent manner, this being accompanied by decreased cardiac contractility and heart rate without compensatory changes in peripheral vascular tone30,35,36. More recently, it was shown that subcutaneous administration of the protein has a profound effect on blood pressure and heart rate in an animal model of hypertension (Dahl salt-sensitive), as well as in 5/6 nephrectomized rats37.
Renalase was suggested to exert its effects through the metabolization of circulating catecholamines and probably other unknown substrates30,37. Under basal conditions, plasma renalase was suggested to circulate as an inactive proenzyme that can be converted to the active form by brief surges in catecholamine levels and increases in blood pressure38. In contrast to plasma renalase, urine renalase exhibits robust amine oxidase activity at baseline and the difference in activity between blood and urine renalase at baseline was suggested to be accounted for by the presence of a renalase inhibitor in plasma39.
RENALASE AND HYPERTENSION
Several lines of evidence suggest that disturbances in the renalase pathway may contribute to increase blood pressure. The first genomic connection between renalase and hypertension was demonstrated by Zhao et al. in the Han Chinese population reporting an association between genetic variants in the renalase coding gene and essential hypertension40.
In a study on neurogenic hypertensive subjects, where norepinephrine spillover from adrenergic nerves was significantly increased, renalase secretion by the kidneys was undetectable in most patients41. Also, arterial plasma levels of renalase were reported to be inversely associated with systolic blood pressure in a cohort of patients with resistant hypertension42.
Experimental data also indicate that renalase deficiency, even in the absence of significant kidney disease, is associated with elevated blood pressure. Actually, downregulation of renalase gene expression in the rat using small interfering RNAs, did not affect renal function but increased blood pressure both at rest and during exogenous catecholamines burst43. Moreover, amino acid mutations that decrease renalases in vitro enzymatic activity to varying degrees, similarly affect its capacity to lower blood pressure, suggesting that this could be mediated by its capacity to degrade circulating catecholamines37. Taken together, these results support the theory that renalase may have a role in the development and maintenance of hypertension.
Although it has been suggested that urinary renalase could have an adverse effect on blood pressure by lowering luminal DA levels44, there is virtually no data on urine renalase levels and activity, namely in relation with changes in renal DA system.
RENALASE AND RENAL DOPAMINERGIC SYSTEMIn studies aimed at increasing renal DA availability by reducing the degradation of renal DA, both the selective and combined inhibition of MAO-A and COMT did not change renal DA excretion or natriuresis, notwithstanding the marked decrease observed in the urinary excretion of the corresponding metabolites of the two enzymes45. This observation suggested that other metabolic pathways in addition to COMT and MAO might be involved in the regulation of renal dopamine output.
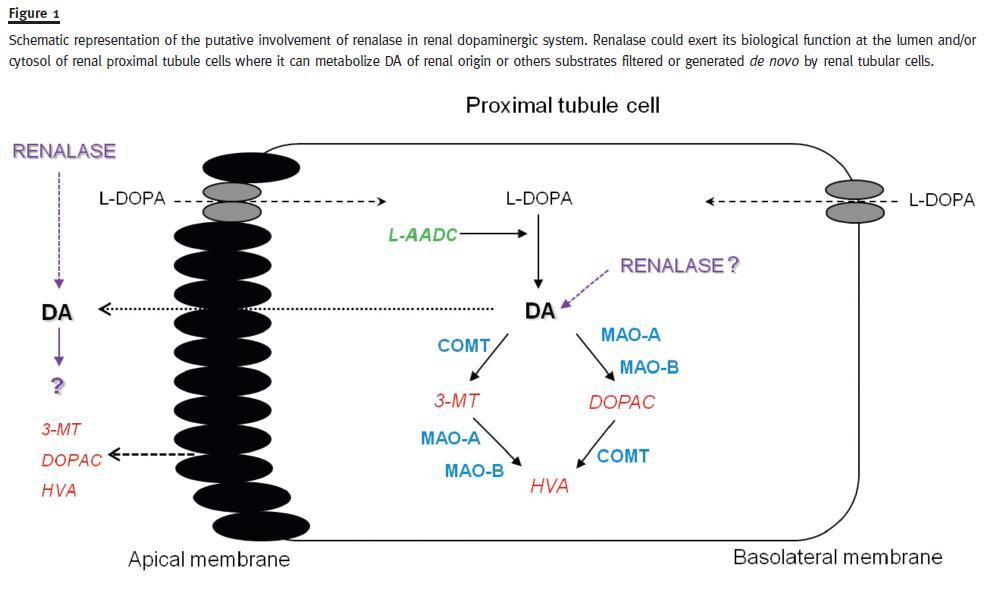
Taking into consideration that renalase is not only secreted in plasma, but is also excreted in urine where it exhibits robust amine oxidase activity at baseline, one can postulate that renalase could be involved in the regulation of renal dopaminergic system activity (Fig. 1).
Renalase deficiency is associated with increased renal DA activityExamination of a global renalase-knockout (KO) mouse model revealed that renalase deficiency is associated with increased plasma catecholamine levels and increased blood pressure46.
For the purpose of examining the influence of renalase on renal DA metabolism, our group assessed renal dopaminergic system activity in the renalase KO mouse model. In these studies, the renalase KO mice presented significant increases in plasma levels of norepinephrine, epinephrine, DA and L-DOPA that were accompanied by a significant increase in urine DA output in comparison in wild-type mice47.
The increased renal DA output in KO mice was observed, notwithstanding an enhanced O-methylation of renal DA and can be explained on the basis of increased renal DA synthesis. The main mechanism underlying the enhanced renal DA synthesis in KO mice appears to be related with increased availability/uptake of L-DOPA in renal proximal tubules (Quelhas-Santos, unpublished observations). Our results also indicated that the signal to increase renal DA synthesis in renalase KO mice is strong as it overcomes a compensatory increase in COMT activity. Because renal DA and norepinephrine exert antagonistic effects on tubular sodium reabsorption2, our findings suggested that the increase in renal DA output in KO mice might behave as a compensatory response induced to maintain sodium homeostasis and control blood pressure, in the presence of enhanced sympathetic tonus47.
The changes induced by recombinant renalase administration, namely in relation with both sympathetic and dopaminergic activities were also evaluated in the renalase KO mouse model. In these studies, we found that the administration of recombinant renalase to KO mice was accompanied by significant decreases in both plasma L-DOPA and catecholamines levels; this was accompanied by marked decreases in both urine DA output and urine DA/L-DOPA ratios48.
Taken together, our findings agree well with the role of renalase as a catecholamine metabolizing enzyme and further reinforce the view that up-regulation of the renal DA synthesis may underly the increased renal DA output observed in renalase KO mice.
Regulation of dopamine D1-like receptors on renalase expression
As mentioned above, renal DA primarily exerts its natriuretic effects via D1-like DA receptors expressed at both the luminal and basolateral membranes of renal tubular cells14,49.
The effect of D1-like receptors on renalase expression and function was recently checked in immortalized renal proximal tubule (RPT) cells from Wistar-Kyoto (WKY) rats and spontaneously hypertensive rats (SHRs)50. In this study, the D1-like agonist fenoldopam increased renalase protein expression and function in WKY RPT cells, but decreased it in SHR cells. Fenoldopam also increased renalase mRNA levels in WKY RPT cells, but not in SHR cells.
These effects were blocked by D1-like receptor antagonist, SCH 23390 in both WKY and SHR cells.
Moreover, inhibition of protein kinase C (PKC), by the PKC inhibitor19-31, blocked the stimulatory effect of fenoldopam on renalase expression, whereas stimulation of PKC, by a PKC agonist (PMA), increased renalase expression, indicating that PKC pathway is involved in this process50.
Additionally, the authors provided evidence that the regulation of renalase by the D1-like receptor was mainly via the D5 receptor because silencing of the D5 but not D1 receptor by antisense oligonucleotides blocked the stimulatory effect of the D1-like receptor on renalase expression in WKY cells. Taken together, these studies suggest that the D5 receptor positively regulates renalase expression in WKY but not in SHR RPT cells and further suggest that aberrant regulation of renalase by the D5 receptor may be involved in the pathogenesis of hypertension50.
Renalase regulates dopamine and phosphate metabolism
The intrarenal dopaminergic system is a key modulator of both renal sodium and phosphate handling51.
Experimental data provided evidence that both phosphate and sodium load downregulate renalase expression and increase renal DA output, thus suggesting the involvement of renalase in sodium and phosphate homeostasis52,53.
The mechanisms mediating renalases effect on phosphate excretion were also studied using the renalase KO mouse model54. Compared with wildtype (WT) mice maintained on a regular diet, KO mice showed increased urinary PO4- excretion and hypophosphataemia. However, both WT and KO mice responded similarly to PO4- restriction by increasing renal COMT-1 activity and markedly decreasing PO4-excretion, thus excluding an intrinsic renal defect in the KO. Renal sodium-phosphate cotransporter Npt2a, sodium proton exchanger NHE3 expression, and MAO-A and B activities did not differ between WT and KO under PO4- restriction. Only COMT expression and activity were significantly increased in KO mice under PO4- restriction. Despite that, urinary DA excretion increased, whereas urinary L-DOPA excretion decreased in the KO mouse, indicating an upregulation of the renal DA synthesis54.
Taking into consideration that increased PO4-intake stimulates renal DA synthesis, and luminal DA inhibits proximal tubular PO4-reabsorption, these data suggest that renalase deficiency is associated with increased renal DA synthesis that contributes to inappropriately high urinary PO4- excretion, which compromise PO4- homeostasis54.
Sodium-dependent modulation of renalase in the rat remnant kidney
Increased dietary sodium intake was shown to down regulate circulating and renal tissues renalase levels in salt-sensitive Dahl rats with well preserved renal function35,53. Our group examined the influence of high sodium intake on circulating renalase levels and activity, as well as on heart and kidney tissue levels of the enzyme in the rat remnant kidney55.
We found that the decrease in circulating and renal tissues renalase levels in ¾nx rats during normal sodium (NS) diet was markedly accentuated during high sodium (HS) intake55. This was accompanied by a decrease in circulating renalase activity in ¾nx rats during HS diet, which was correlated negatively with the increase in blood pressure. Taken together, these findings suggested that, by decreasing systemic renalase levels and activity, the excess of sodium intake may contribute to increase blood pressure in the rat remnant kidney55.
In contrast to what was found in plasma, no significant differences were observed in either urine renalase levels or activity between ¾nx and Sham rats during NS diet, notwithstanding the marked decrease in daily urine DA output observed in the rat remnant kidney. Interestingly, urinary renalase levels and activity were markedly increased in both ¾nx and Sham rats during HS diet in comparison with NS intake and this was accompanied by a paralleled sodium-induced increase in daily urinary DA output in both groups55. Further studies are needed to prove or disprove the possible role of renalase on regulation of the renal DA activity.
Renalase in relation with renal function in Humans
In 2005, Desir and co-workers reported that plasma renalase levels detectable by western-blotting were decreased in patients with CKD and ESRD30. Other studies showed that plasma, kidney and cardiac renalase levels were decreased in various rat models of CKD and that this was accompanied by increased levels of epinephrine and norepinephrine in both plasma and heart30,34,39,56. However, a Polish group headed by Mysliwiec reported that plasma renalase levels assessed by a commercially enzyme-linked imunosorbent assay (ELISA), are negatively related with renal function in several populations, namely in kidney transplant recipients57, heart transplant recipients58, patients on peritoneal dialysis59 and patients on haemodialysis60.
In those studies, plasma renalase levels were dependent on residual renal function and were lower in CKD patients with residual renal function when compared with anuric patients61.
Recently, our group examined the link between renal function and the renalase pathway in patients with ESRD, as well as during the recovery of renal function in kidney transplant recipients62. In this study, we found that plasma renalase levels and activity were significantly increased in ESRD patients. This was accompanied in ESRD patients by a marked increase in urinary renalase levels and activity. The recovery of renal function closely predicted decreases in plasma renalase levels, as well as in urine renalase levels and activity. Urine DA levels negatively correlated with urinary renalase levels but did not correlate with urine renalase activity.
These findings, when viewed collectively with the observations that renalase can metabolize DA37, suggest that urinary renalase might play a role in the regulation of renal DA output.
What is new?
Renalase deficiency in renalase KO mice is accompanied by increased renal DA activity that may behave as a compensatory response to heightened sympathetic activity.
The increased renal DA output in renalase KO mice can be explained on the basis of enhanced renal DA synthesis.
In the rat remnant kidney, the excess of sodium intake significantly decreases circulating renalase levels and activity while markedly increasing urinary renalase levels and activity.
In humans, urinary renalase levels and activity strongly depend on renal function.
References
1. Ortiz PA, Garvin JL. Intrarenal transport and vasoactive substances in hypertension. Hypertension 2001;38(3 Pt 2):621-624. [ Links ]
2. Aperia AC. Intrarenal dopamine: a key signal in the interactive regulation of sodium metabolism. Annu Rev Physiol 2000;62:621-647. [ Links ]
3. Siragy HM, Felder RA, Howell NL, Chevalier RL, Peach MJ, Carey RM. Evidence that intrarenal dopamine acts as a paracrine substance at the renal tubule. Am J Physiol 1989;257(3 Pt 2):F469-477. [ Links ]
4. Lee MR. Dopamine and the kidney: ten years on. Clin Sci (Lond) 1993;84(4):357-375. [ Links ]
5. Soares-da-Silva P, Fernandes MH, Pinto-do-O PC. Cell inward transport of L-DOPA and 3-O-methyl-L-DOPA in rat renal tubules. Br J Pharmacol 1994;112(2):611-615. [ Links ]
6. Pinto V, Pinho MJ, Soares-da-Silva P. Renal amino acid transport systems and essential hypertension. FASEB J 2013;27(8):2927-2938. [ Links ]
7. Fernandes MH, Soares-da-Silva P. Role of monoamine oxidase and catechol-Omethyltransferase in the metabolism of renal dopamine. J Neural Transm Suppl 1994;41:101-105. [ Links ]
8.
9. Hussain T, Lokhandwala MF. Renal dopamine receptors and hypertension. Exp Biol Med (Maywood) 2003;228(2):134-142. [ Links ]
10. Zeng C, Yang Z, Asico LD, Jose PA. Regulation of blood pressure by D5 dopamine receptors. Cardiovasc Hematol Agents Med Chem 2007;5(3):241-248. [ Links ]
11. Zeng C, Wang D, Asico LD, et al. Aberrant D1 and D3 dopamine receptor transregulation in hypertension. Hypertension 2004;43(3):654-660. [ Links ]
12. Felder CC, Campbell T, Albrecht F, Jose PA. Dopamine inhibits Na(+)-H+ exchanger activity in renal BBMV by stimulation of adenylate cyclase. Am J Physiol 1990;259(2 Pt 2):F297-303. [ Links ]
13. Lokhandwala MF, Amenta F. Anatomical distribution and function of dopamine receptors in the kidney. FASEB J 1991;5(15):3023-3030. [ Links ]
14. Jose PA, Raymond JR, Bates MD, Aperia A, Felder RA, Carey RM. The renal dopamine receptors. J Am Soc Nephrol 1992;2(8):1265-1278. [ Links ]
15. Pelayo JC, Fildes RD, Eisner GM, Jose PA. Effects of dopamine blockade on renal sodium excretion. Am J Physiol 1983;245(2):F247-253. [ Links ]
16. Pinho MJ, Serrão MP, Gomes P, Hopfer U, Jose PA, Soares-da-Silva P. Over-expression of renal LAT1 and LAT2 and enhanced L-DOPA uptake in SHR immortalized renal proximal tubular cells. Kidney Int 2004;66(1):216-226. [ Links ]
17. Goldstein DS, Stull R, Eisenhofer G, Gill JR, Jr. Urinary excretion of dihydroxyphenylalanine and dopamine during alterations of dietary salt intake in humans. Clin Sci (Lond) 1989;76(5):517-522. [ Links ]
18. Seri I. Dopamine and natriuresis. Mechanism of action and developmental aspects. Am J Hypertens 1990;3(6 Pt 2):82S-86S. [ Links ]
19. Vieira-Coelho MA, Serrão P, Guimaraes JT, Pestana M, Soares-da-Silva P. Concerted action of dopamine on renal and intestinal Na(+)-K(+)-ATPase in the rat remnant kidney. Am J Physiol Renal Physiol 2000;279(6):F1033-1044. [ Links ]
20. Hegde SS, Jadhav AL, Lokhandwala MF. Role of kidney dopamine in the natriuretic response to volume expansion in rats. Hypertension 1989;13(6 Pt 2):828-834. [ Links ]
21. Sampaio-Maia B, Serrão P, Guimarães JT, Vieira-Coelho MA, Pestana M. Renal dopaminergic system activity in the rat remnant kidney. Nephron Exp Nephrol 2005;99(2):e46-55. [ Links ]
22. Grossman E, Hoffman A, Tamrat M, Armando I, Keiser HR, Goldstein DS. Endogenous dopa and dopamine responses to dietary salt loading in salt-sensitive rats. J Hypertens 1991;9(3):259-263. [ Links ]
23. Seri I, Kone BC, Gullans SR, Aperia A, Brenner BM, Ballermann BJ. Influence of Na+ intake on dopamine-induced inhibition of renal cortical Na(+)-K(+)-ATPase. Am J Physiol 1990;258(1 Pt 2):F52-60. [ Links ]
24. Vieira-Coelho MA, Hussain T, Kansra V, et al. Aging, high salt intake, and renal dopaminergic activity in Fischer 344 rats. Hypertension 1999;34(4 Pt 1):666-672. [ Links ]
25. Pestana M, Jardim H, Serrão P, Soares-da-Silva P, Guerra L. Reduced urinary excretion of dopamine and metabolites in chronic renal parenchymal disease. Kidney Blood Press Res 1998;21(1):59-65. [ Links ]
26. Moreira-Rodrigues M, Sampaio-Maia B, Pestana M. Renal dopaminergic system activity in rat remnant kidney up to twenty-six weeks after surgery. Life Sci 2009;84(13-14):409-414. [ Links ]
27. Koomans HA, Blankestijn PJ, Joles JA. Sympathetic hyperactivity in chronic renal failure: a wake-up call. J Am Soc Nephrol 2004;15(3):524-537. [ Links ]
28. Pestana M, Jardim H, Serrão P, Soares-da-Silva P, Guerra L. Reduced urinary excretion of dopamine and metabolites in chronic renal parenchymal disease. Kidney Blood Press Res 1998;21(1):59-65. [ Links ]
29. Pestana M, Jardim H, Correia F, Vieira-Coelho MA, Soares-da-Silva P. Renal dopaminergic mechanisms in renal parenchymal diseases and hypertension. Nephrol Dial Transplant 2001;16 Suppl 1:53-59. [ Links ]
30. Xu J, Li G, Wang P, et al. Renalase is a novel, soluble monoamine oxidase that regulates cardiac function and blood pressure. J Clin Invest 2005;115(5):1275-1280. [ Links ]
31. Eikelis N, Hennebry SC, Socratous F, McGrane M, Chopra R, Decourten B, et al. Reduction in Renalase Protein Expression in Adipose Tissue after Weight Loss. Journal of Hypertension. 2009 Jun;27:S160-S1. [ Links ]
32. Hennebry SC, Eikelis N, Socratous F, Desir G, Lambert GW, Straznicky N, et al. Central and Peripheral Distribution of Renalase, a Novel Soluble Monoamine Oxidase, in Human Tissue. Journal of Hypertension. 2009 Jun;27:S33-S4. [ Links ]
33. Hennebry SC, Eikelis N, Socratous F, Desir G, Lambert G, Schlaich M. Renalase, a novel soluble FAD-dependent protein, is synthesized in the brain and peripheral nerves. Mol Psychiatry 2010;15(3):234-236. [ Links ]
34. Ghosh SS, Krieg RJ, Sica DA, Wang R, Fakhry I, Gehr T. Cardiac hypertrophy in neonatal nephrectomized rats: the role of the sympathetic nervous system. Pediatr Nephrol 2009;24(2):367-377. [ Links ]
35. Desir GV. Regulation of blood pressure and cardiovascular function by renalase. Kidney Int 2009;76(4):366-370. [ Links ]
36. Desir GV. Renalase is a novel renal hormone that regulates cardiovascular function. J Am Soc Hypertens 2007;1(2):99-103. [ Links ]
37. Desir GV, Tang L, Wang P, et al. Renalase lowers ambulatory blood pressure by metabolizing circulating adrenaline. J Am Heart Assoc 2012;1(4):e002634. [ Links ]
38. Desir GV. Renalase deficiency in chronic kidney disease, and its contribution to hypertension and cardiovascular disease. Curr Opin Nephrol Hypertens 2008;17(2):181-185. [ Links ]
39. Li G, Xu J, Wang P, Velazquez H, Li Y, Wu Y, Desir GV. Catecholamines regulate the activity, secretion, and synthesis of renalase. Circulation 2008;117(10):1277-1282. [ Links ]
40. Zhao Q, Fan Z, He J, et al. Renalase gene is a novel susceptibility gene for essentialhypertension: a two-stage association study in northern Han Chinese population. J Mol Med (Berl) 2007;85(8):877-885. [ Links ]
41. Socratous F, Eikelis N, Hennebry S, Schlaich MP. Altered renalase secretion in neurogenic human hypertension. Hypertension. 2009;53:(1117). [ Links ]
42. Schlaich M, Socratous F, Eikelis N, Chopra R, Lambert G, Hennebry S. Renalase plasma levels are associated with systolic blood pressure in patients with resistant hypertension. Journal of Hypertension. 2010;28(p e437). [ Links ]
43. Ghosh SS, Gehr TWB, Sica DA, Masilamani S, Ghosh S, Wang R, et al. Effect of renalase inhibition on blood pressure. J Am Soc Nephrol. 2006;17:208:A. [ Links ]
44. Desir GV. Role of renalase in the regulation of blood pressure and the renal dopamine system. Curr Opin Nephrol Hypertens 2011;20(1):31-36. [ Links ]
45. Sampaio-Maia B, Moreira-Rodrigues M, Pestana M. Role of chronic inhibition of dopamine-metabolizing enzymes in the regulation of renal sodium and phosphate excretion in the rat remnant kidney. Nephron Physiol 2006;103(1):p14-24. [ Links ]
46. Wu Y, Xu J, Velazquez H, Wang P, Li G, Liu D, et al. Renalase deficiency aggravates ischemic myocardial damage. Kidney Int. 2011 Apr;79(8):853-60. [ Links ]
47. Quelhas-Santos J, Serrão P., Fernandes-Cerqueira C., Simões-Silva L., Soares-Silva I., Sizova D., et al. Renal dopaminergic activity is increased in the renalase knockout mouse model.
48. Quelhas-Santos J, Serrão P., Soares-Silva I., Tang L., Sampaio-Maia B., Desir G., et al. Effects of recombinant renalase on renal dopamine activity in the renalase knockout mouse model. NDT Plus. 2012;27(2):ii330-ii2. [ Links ]
49. Hussain T, Lokhandwala MF. Renal dopamine receptor function in hypertension. Hypertension 1998;32(2):187-197. [ Links ]
50. Wang S, Lu X, Yang J, et al. Regulation of renalase expression by D5 dopamine receptors in rat renal proximal tubule cells. Am J Physiol Renal Physiol 2014;306(6):F588-596. [ Links ]
51. Carey RM. Theodore Cooper Lecture: Renal dopamine system: paracrine regulator of sodium homeostasis and blood pressure. Hypertension 2001;38(3):297-302. [ Links ]
52. Weinman EJ, Biswas R, Steplock D, et al. Increased renal dopamine and acute renal adaptation to a high-phosphate diet. Am J Physiol Renal Physiol 2011;300(5):F1123-1129. [ Links ]
53. Ghosh SS, Gehr TWB, Sica DA, Masilamani S, Fakhry I, Wang R, et al. Renalase regulates blood pressure in salt sensitive Dahl rats. J Am Soc Nephrol 2006;17:208:A. [ Links ]
54. Sizova D, Velazquez H, Sampaio-Maia B, Quelhas-Santos J, Pestana M, Desir GV. Renalase regulates renal dopamine and phosphate metabolism. Am J Physiol Renal Physiol 2013;305(6):F839-844. [ Links ]
55.
56. Gu R, Lu W, Xie J, Bai J, Xu B. Renalase deficiency in heart failure model of rats–a potential mechanism underlying circulating norepinephrine accumulation. PLoS One 2011;6(1):e14633. [ Links ]
57. Malyszko J, Zbroch E, Malyszko JS, Koc-Zorawska E, Mysliwiec M. Renalase, a novel regulator of blood pressure, is predicted by kidney function in renal transplant recipients. Transplant Proc 2011;43(8):3004-3007. [ Links ]
58. Przybylowski P, Malyszko J, Kozlowska S, Malyszko J, Koc-Zorawska E, Mysliwiec M. Serum renalase depends on kidney function but not on blood pressure in heart transplant recipients. Transplant Proc 2011;43(10):3888-3891. [ Links ]
59. Zbroch E, Malyszko J, Koc-Zorawska E, Mysliwiec M. Renalase in peritoneal dialysis patients is not related to blood pressure, but to dialysis vintage. Perit Dial Int 2012;32(3):348-351. [ Links ]
60. Zbroch E, Malyszko J, Malyszko JS, Koc-Zorawska E, Mysliwiec M. Renalase, a novel enzyme involved in blood pressure regulation, is related to kidney function but not to blood pressure in hemodialysis patients. Kidney Blood Press Res 2012;35(6):395-399. [ Links ]
61. Malyszko J, Koc-Zorawska E, Malyszko JS, Kozminski P, Zbroch E, Mysliwiec M. Renalase, stroke, and hypertension in hemodialyzed patients. Ren Fail 2012;34(6):727-731. [ Links ]
62. Quelhas-Santos J, Soares-Silva I, Fernandes-Cerqueira C, Simões-Silva L, Ferreira I, Carvalho C, et al. Plasma and Urine Renalase Levels and Activity during the Recovery of Renal Function in kidney Transplant Recipients. Exp Biol Med (Maywood) 2014; Mar 5. [Epub ahead of print]. [ Links ]
Prof. Dr. Janete Quelhas dos Santos
Nephrology and Infectious Diseases Research & Development Group, INEB,University of Porto.
Praça Gomes Teixeira 4099-002
Porto, Portugal
Email: sjanete@med.up.pt
Conflict of interests Statement: None declared
Received for publication: 17/03/2014
Accepted: 19/03/2014

{kind=link}