









Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkPortuguese Journal of Nephrology & Hypertension
Print version ISSN 0872-0169
Port J Nephrol Hypert vol.29 no.4 Lisboa Dec. 2015
REVIEW ARTICLE
Lipotoxicity and kidney
Lipotoxicidade e rim
Ana Rita Martins1, Sebastian Mas2
1Nephrology Department, Hospital Santa Cruz, Carnaxide, Portugal.
2Renal, Vascular and Diabetes Research Laboratory. IIS-Fundación Jiménez Díaz, Madrid, Spain.
ABSTRACT
Lipotoxicity could be defined as the deleterious effects exerted by lipids on plasma (normally as nonesterified molecules) on cell and tissues. Lipotoxicity is closely linked to obesity, although it can be observed in some lean individuals. Therefore, as overweight becomes more prevalent worldwide, the pathological effects mediated by lipotoxic molecules will have a greater impact in healthcare systems.
Ectopic deposition of lipids into non-adipose tissues, such as the kidney, can lead to accumulation of toxic metabolites that lead to mitochondrial dysfunction, endoplasmic reticulum stress, apoptosis and renal dysfunction. Pro-inflammatory mediators are released by visceral adipose tissue in response to injury: adipocytokines (leptin, resistin, vistatin, adiponectin), chemokines (IP-10, IL-8, RANTES and MCP-1), interleukins (IL-1, IL-6) and growth factors (TNFa).
Visceral adiposity plays an important role in inflammation and insulin resistance in obesity. Oxidative processes are pivotal events in injury to renal tubular and epithelial cells exposed to ox-LDL. In type 2 diabetes, ox-LDL and free fatty acids damage podocyte function, structure and tubulointersticial tissue.
The mechanisms by which obesity causes nephron injury are not fully understood but likely involve a combination of haemodynamic, metabolic and inflammatory changes. Most of the research has been done on its effects in cardiovascular disease, but lipotoxicity is also a major contributor to the development of chronic kidney disease (CKD). The understanding of the adipogenesis should provide new opportunities for therapeutic intervention.
In this paper the current state of causes, consequences and possible therapeutic intervention is summarized, providing an insight on the contribution of lipotoxic molecules to renal disease.
Key-Words: Lipotoxicity, obesity, visceral adipose tissue, adipocytokines, insulin resistance
RESUMO
A lipotoxicidade define-se pelos efeitos deletérios causados pelos lípidos plasmáticos em circulação (usualmente sob a forma de moléculas não-esterificadas) nas células e nos tecidos. A lipotoxicidade está intimamente relacionada com a obesidade, embora possa ser observada em alguns indivíduos magros. Com o aumento da prevalência da obesidade por todo o mundo, também os efeitos patológicos mediados pelas moléculas lipotóxicas terão maior impacto na população.
A deposição ectópica de lípidos em tecidos não adiposos, tais como o rim, pode levar à acumulação de metabolitos tóxicos que levam à disfunção mitocondrial, ao stress do retículo endoplasmático, apoptose e à disfunção renal. Os mediadores pro-inflamatórios libertados pelo tecido adiposo visceral em resposta a lesão são: adipocitocinas (leptina, resistina, vistatina, adiponectina), quimiocinas (IP-10, IL-8, RANTES e MCP-1), interleucinas (IL-1, IL-6) e fatores de crescimento (TNFa).
A adiposidade visceral tem um papel central na inflamação e na resistência à insulina na obesidade. Os processos oxidativos têm papel fulcral na lesão tubular renal e nas células epiteliais expostas a LDL-ox. Na diabetes tipo 2, LDL.ox e ácidos gordos livres causam dano na função e estrutura podocitária e no tecido tubulointersticial.
Os mecanismos pelos quais a obesidade causa lesão renal não estão ainda completamente esclarecidos mas envolvem provavelmente uma combinação de vias hemodinâmicas, metabólicas e inflamatórias. A investigação tem sido, sobretudo, centrada sobre os efeitos da lipotoxicidade na doença cardiovascular, porém tem também um importante impacto na doença renal crónica (DRC). O entendimento da adipogénese pode oferecer oportunidades para novas intervenções terapêuticas.
Neste artigo, os autores apresentam o conhecimento atual das causas, consequências e intervenções terapêuticas, proporcionando uma visão acerca da contribuição das moléculas lipotóxicas na doença renal.
Palavras-Chave: Lipotoxicidade, obesidade, tecido adiposo visceral, adipocitocinas, insulino-resistência
LIPOTOXICITY
Circulating non-esterified fatty acids (NEFA) in obesity and diabetes
Among the lipids that could induce cell damage, non-esterified fatty acids (NEFA) are the most abundant and, therefore, their production and actions are well known, constituting in most cases a synonym of lipotoxicity. NEFA are the preferential energy source of some highly metabolic organs, such as the myocardium or the liver. Physiological lipolysis of triglycerides occurs in adipose tissue (mainly subcutaneous) and its regulation depends on energy requirements.
NEFAs are transported bound to albumin molecules (which have a half-life of 4-6 min)1.
Serum NEFA concentrations are very variable and affected by gender, fasting state and psychological stress (causing concentrations as high as > 1700 μmol/L) and can be supressed by high-carbohydrate diets2.
But in those physiological conditions, plasma NEFA concentrations remain relatively stable as removal by muscle increases to match adipose tissue lipolysis1,3.
In conditions like obesity, the increase in adipose tissue is expected to correlate with NEFA plasma concentration and, although several studies seem to indicate a direct relationship, recent systematic review shows no apparent correlation between both parameters4. But, due to the variation of NEFA concentration with physiological parameters, further studies will require homogenous measurements taking into account cyrcadian rythms, fasting time and gender.
In type 2 diabetes, data from clinical studies are clearer. The first observation of the correlation between NEFA and diabetes occured in a native American population, in the mid 1990s5. This observation was later confirmed in larger cohorts, such as the Paris Prospective Study, in which fasting NEFA concentration correlated with tolerance to glucose deterioration, independently of previous insulin resistance or insulin secretion defects6. Thus, a high serum NEFA concentration can be considered a pathognomonic sign in type 2 diabetes7 and some authors consider that it is an independent risk factor of insulin resistance in obese patients. A high concentration of NEFA on fasting is related with the amount of visceral adipose tissue.
Adipose tissue dysfunction
Adipose tissue is an important endocrine organ, in addition to its role as an energy storage organ, secreting a large number of cytokines (adipokines). White adipose tissue (WAT) is composed by lipid-laden mature adipocytes, various stromal cells, such as pre-adipocytes, endothelial cells, fibroblasts and immune cells that may be involved in adipose tissue functions. Macrophages represent a major source of proinflammatory cytokines, which contribute to chronic inflammatory response. Brown adipocytes are specialized for energy expenditure, glucose oxidation and insulin sensitivity. Beige adipocytes share many of the oxydative properties of the brown adipocytes1,8.
In the last decade, adipose tissue has been considered to play a central role on insulin resistance and diabetes. The distribution of adipose tissue is also of special importance to both insulin resistance and type 2 diabetes. It is widely accepted that visceral adipose tissue is metabolically highly active and plays a major role in the pathogenesis of insulin resistance8,9.
Since visceral WAT is resistant to the antilipolytic actions of insulin, it releases NEFA via portal circulation. Adipose tissue becomes dysfunctional due to its rapid expansion or contraction10. Several factors have been postulated to play a role on adipocyte dysfunction: the lipotoxicity itself; lipid-mediated Tolllike receptor (TLR) activation11; hypoxia12 and endoplasmic reticulum (ER)-stress on hypertrophic cells which leads to immune recruitment into the stressed tissue13. This inflammation pattern results from the interaction between parenchymal and stromal cells in response to stress leading to adipose tissue remodelling, which can be defined as the dynamic histological changes observed in adipose tissue14. Evidence suggests that the interaction between mature adipocytes and stromal cells plays a crucial role in the regulation of adipocytokine production15.
Under appropriate stimulation adipose precursor cells can acquire some phagocytic capacity and, thereby, become macrophage-like16. Many pro-inflammatory mediators are released by visceral adipose tissue in response to injury: adipocytokines (leptin, resistin, vistatin, adiponectin), chemokines (IP-10, IL-8, RANTES and MCP-1), interleukins (IL-1, IL-6) and growth factors (TNFa)11, 14. There is a paracrine regulation of adipose tissue involving saturated fatty acids and TNFa derived from adipocytes and macrophages, respectively, which establishes a vicious cycle that augments chronic inflammatory changes. The net effect is T cell and macrophage recruitment into the adipose tissue promoting its activation14,16.
Macrophages are of central importance and there are two kinds during the inflammatory process: M1 macrophages, responsible for the chronic inflammation and M2 macrophages in the interstitial space that can be polarized into M1 by stimulation with cytokines. In obese patients, phenotypic change from M2 to M1 macrophages also accelerates adipose tissue inflammation14,16.
Usually in obese individuals, the number of M1 macrophages is higher in visceral fat than subcutaneous fat, one of the reasons for a more proinflammatory profile of visceral fat16. Infiltration and activation of monocytes/macrophages into the adipose tissue not only drives to induction and secretion of proinflammatory chemokines as well as increases fibrosis14,16.
Other immune cells involved in the inflammatory process are neutrophils, eosinophils, lymphocytes, natural killer cells and mast cells14, 16. Two subsets of T cells have been identified: CD8+ T cells, increased in the earlier stages of obesity, their depletion results in improvement of adipose tissue inflammation and insulin resistance. CD4+ T cells and regulatory T cells which are decreased in the advanced stage of obesity16.
An imbalance of T cell subpopulations may also contribute to adipose tissue inflammation.
Some antidiabetic drugs are known to modulate adipocyte-associated inflammation. Thiazolidinedione showed to be capable to improve the unbalanced M1/M2 phenotype of adipose tissue macrophages in dietinduced obese mice17. But still, visceral WAT inflammation may be a promising therapeutic target to prevent or treat obesity-induced adipose tissue inflammation.
Lipotoxicity and insulin resistance
In early stages of obesity, insulin action is still preserved or even enhanced as well as its action on lipogenesis and adipogenesis. In obese individuals, insulin secretion increases with insulin resistance and patients who are predisposed to diabetes fail to compensate for greater insulin requirements and tend to develop type 2 diabetes15, 18. However, even in the absence of diabetes, patients with renal disfunction exhibit insulin resistance with hypertriglyceridemia more frequently than age and BMI-matched control subjects. Insulin resistance is believed to contribute to the burden of CKD on the cardiovascular outcome of these patients18,19.
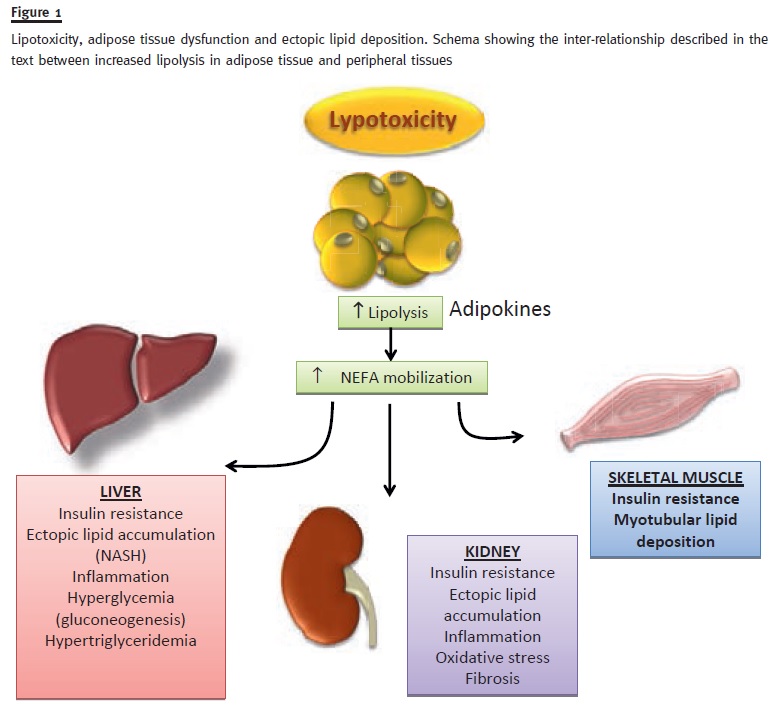
There is a well-kown relation between lipid excess and insulin resistance, particularly regarding Ana Rita Martins, Sebastian Más intramyocellular lipid content1,15. Traditionally, it has been explained via the glucose-fatty acid cycle, also known as Randle cycle. This regulatory pathway, involves the cross-inhibition of glucose and fatty acids over the opposite catabolisms20. Thus, the high level of Acetyl-CoA that results from FA degration acts as an allosteric regulator of pyruvate dehydrogenase, leading to the inhibition of glycolysis20(Figure 1).
Recently, Shulman has reviewed the relationship between lipid accumulation and insulin resistance showing that much of the data gathered in the last 15 years do not fit the classical theory21. On the contrary, magnetic resonance spectroscopy studies have shown a decrease on the level of intramuscular glucose and glucose-6-phospate after i.v. lipid infusion22.
Using lipid-infused rodents to characterize the mechanism involved, these studies have identified transient increases in muscle diacylglycerol (DAG) content23. High intracellular DAG levels trigger the novel protein kinase C theta (PKCΘ) isoform24, which is known to be active in the absence of Ca2+ which, when present in skeletal muscle, drives the serinphosphorylation of several substrates potentially involved in insulin resistance.
The most important substrate described is the insulin receptor substrate 1 (IRS-1) which is phosphorylated at Ser 1101, inactivating the receptor and impairing insulin signaling25. These observations are confirmed in muscle biopsies of patients with insulin resistance26 and type 2 diabetes15.
Lypotoxicity and dyslipidemia
Lipotoxicity is clinically observed as a part of the metabolic syndrome1,15,18. Hepatic dysfuntion with non-alcoholic hepatic steatosis (NAFLD), is associated with insulin resistance lead to de novo lipogenesis and triglyceride esterification and secretion as VLDL8,27. This secretion is responsible, at least partially, for the triglyceridemia observed in those patients.
Strong associations have been reported between amount of liver fat and features of the cardiometabolic risk profile predicting both risk of type 2 diabetes and cardiovascular disease13.
Thus, the relation between circulating NEFA and hipertrygliceridemia is reciprocal as lipotoxicity leads to triglyceride secretion and sustained hypertriglyceridemia leads to adipose dysfuntion1. Also, increased liver fat and visceral adipose tissue are both associated with augmented cardiometabolic risk19.
LIPOTOXICITY AND THE KIDNEY
Lipid metabolism in the kidney
In the last decade, research in pathology has shown that CKD may constitute a state of acquired lipodystrophy, which leads to ectopic lipid deposition.
For decades, CKD has been described as a cause of dyslipidemia with increased plasma triglycerides28.
Recent studies show that adipose tissue distribution may be altered in CKD, resulting in a loss of white adipose tissue and accumulation of lipids in ectopic sites29. The body mass index does not reflect the amount of atherogenic visceral and ectopic adipose tissue, particularly in CKD patients18.
Experimental evidence showed that CKD is associated with reduction of fat deposit sizes and ectopic lipid distribution28,29. This is also common in aging and lipodystrophic syndromes30. Since adipocyte size and number are related to insulin sensitivity, glucose and fatty acid uptake as well as cytokine release, changes in function and cellular composition of white adipose tissue may lead to changes in metabolic state and subsequent complications31. The main function of the white adipose tissue is to store the energy surplus in the form of lipids, by converting circulating cytotoxic NEFA into triglycerides1.
Also in renal disease, profiles of adipokines are altered32. The incubation of adipocytes from visceral adipose tissue in uraemic plasma increases the secretion of leptin33. Also, there seems to exist a crosstalk between white adipose tissue and the kidney through adiponectin. Plasma adiponectin concentration is lower in obese patients, probably due to suppression of its production by macrophages infiltrating adipose tissue in these individuals. Adiponectin has beneficial cardiovascular effects and improves insulin sensitivity.
Sharma et al.34 showed that adiponectin null-mice exhibit proteinuria with foot process disappearance and that treatment with adiponectin normalizes proteinuria through stimulation of AMPK and suppression of an isoform of NADPH-oxidase.
Lipotoxicity in CKD seems to be partly related to adipose tissue dysfunction1. Incubation of human adipocytes in uraemic sera results in an increase in baseline lipolysis and a decreased expression of the messenger RNA that codes perlipin35. The decline in fat mass and ability to store lipids seen in CKD seems to contribute to fat redistribution into other tissues31.
Ectopic lipid deposition on the kidney
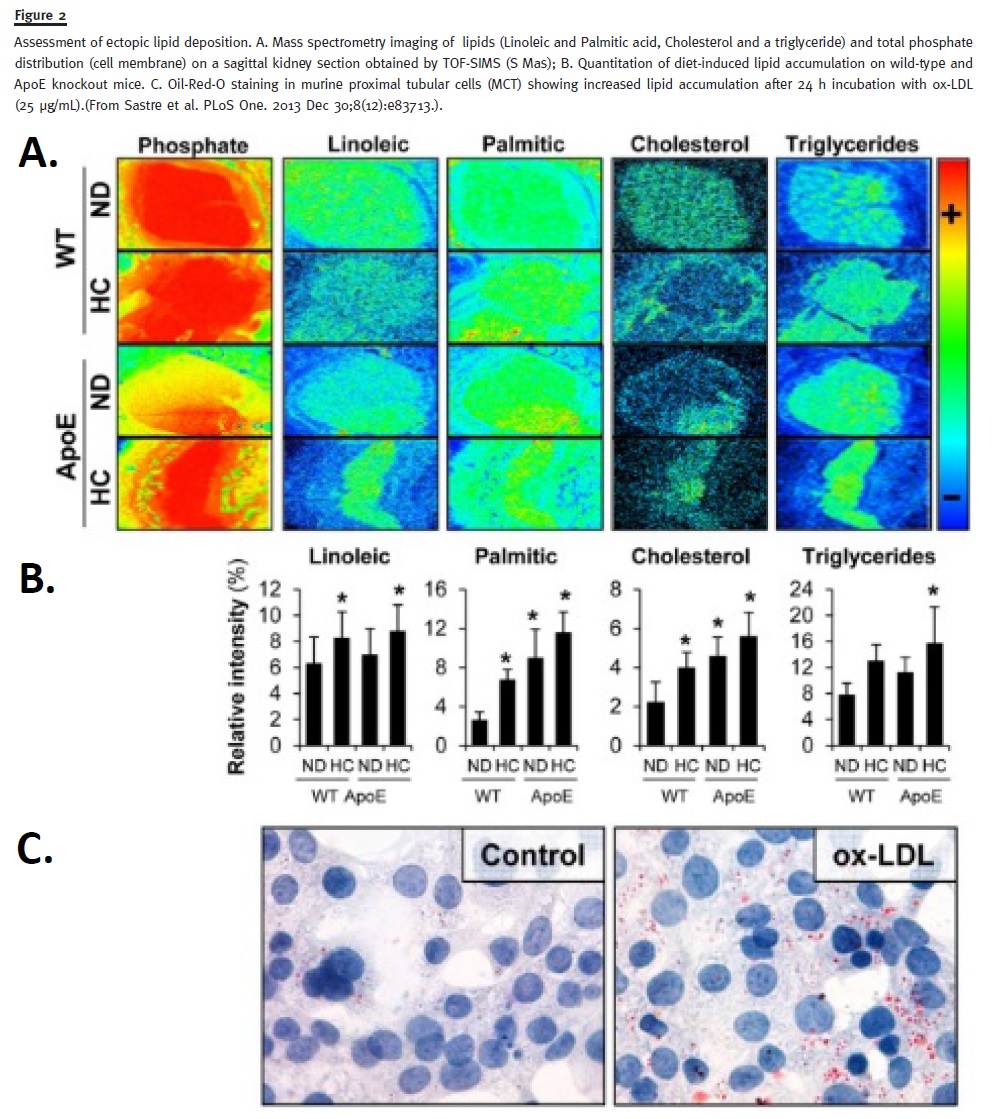
Lipid accumulation occurs, not only in adipose tissue, but also in non-adipose tissues (e.g., ectopic fat deposition and lipotoxicity in kidney – Figure 2)36. When the energy intake exceeds the storage capacity of subcutaneous white adipose tissue, lipid accumulation is induced in ectopic sites37. Triglycerides are not considered toxic per se. On the contrary, they are safe storage forms. The principal determinant of lipotoxicity seems to be excessive intracellular free fatty acid content which leads to the accumulation of potentially toxic metabolites such as fatty acyl-CoA, diacilglycerols and ceramides38.
These metabolites may lead to mitochondrial dysfunction, endoplasmic reticulum stress, apoptosis and eventually to dysfunction and injury. The contribution of 3-hydroxy-3-methylglutaryl-coenzyme A reductase inhibitors to the improvement of proteinuria and the preservation of renal function seems to confirm the role of lipids in renal damage37,39. In the last decades, lipid ectopic accumulation has started to be considered a relevant factor in metabolic disease40. The first and most relevant pathological condition caused by ectopic lipid accumulation is non-alcoholic fatty liver disease, the most common chronic liver disease in the world, which is associated with insulin-resistance37. Nowadays, a high intramyocellular fat level is considered a sign of insulin resistance31, and this process is closely related to visceral fat accumulation32.
Lipid accumulation in ectopic sites is also a characteristic feature of lipodystrophy, a metabolic disorder in which adipose tissue loses its ability to buffer the post-prandial influx of fatty acids28,27. As ectopic fat accumulation damages insulin response, it may lead to impaired insulin synthesis and secretion by pancreatic beta cells8,9. Hence, diabetes may occur, and both lipogenesis and adipogenesis may become stationary while lipolysis is activated. The resultant increase in circulating non-esterified fatty acids contributes to the increased reliance on fat oxidation to provide energy and the generation of fat oxidation by-products supresses glucose oxidation41. Additionally, lipid accumulation in ectopic sites contributes to impaired mitochondrial function30. Evidence suggests that mitochondrial dysfunction, which can accelerate fat accumulation, can be a consequence of insulin resistance rather than a primary defect41.
Lipotoxic cellular dysfunctions and injuries occur through several mechanisms, including the generation of reactive oxygen species (ROS); multiple organellar damages and disruption of intracellular signaling pathways; the release of proinflammatory and profibrotic factors; and lipid-induced apoptosis13.
In mice fed with a high-fat diet, the accumulation of lipids in the glomerular and tubulointerstitial cells (Figure 2) with increased expression of sterol regulatory element binding protein (SREBP-1), a transcription factor which plays a key role in lipogenesis and its related genes (ACC, acetyl-coA carboxylase, FAS, fatty acid synthase).
These changes lead to significant glomerulosclerosis and proteinuria41. These data were supported by finding a protection from these effects in mice lacking SREBP-1, when submitted to a high-fat diet42,43.
Excessive lipid droplets can be found in different renal cell types including podocytes. Recent data showed that cholesterol accumulation in podocytes plays a critical role in diabetic nephropathy and contributes to lipotoxicity39,44. Oxidized lipids initiate and modulate the inflammatory cellular events with the formation of macrophages foam cells. Evidence has reported that ox-LDL induces loss of nephrin expression in cultured podocytes44.
Microalbuminuria, a major feature of diabetic nephropathy, is normally considered a consequence of hyperglycaemia and associated glomerular hyperfiltration28,45. However, it also occurs in non-diabetic insulinresistant patients, and studies have shown that lipid accumulation in the kidney may be the cause of renal injury associated with the metabolic syndrome8,9,31.
Renal glucose output is markedly increased in type 2 diabetes, resulting in an increased net renal glucose uptake. It has been proposed that this glucose overload may contribute to the glycogen deposit, which is not associated with an evident fat intrarenal accumulation. In diabetic patients, renal glucose uptake is inversely correlated with FFA uptake, suggesting the existence of a renal glucose-FFA cycle20,33.
In renal, as well as in the overall systemic vascular bed, the generation of oxidized lipids can modulate the formation of macrophage foam cells35. An imbalance between the syntheses of ROS and antioxidants is described as oxidative stress and also as inflammatory state. NADPH oxidase is the enzyme responsible for ROS production and the inhibition of this enzyme weakens the focal adhesion of macrophages within the intimal plaque, restoring their mobility31,34,35.
In inflammatory states, lipids are deposited in tissues and become oxidized. Inflammatorycytokines and growth factors may enhance lipid synthesis, leading to significant intracellular lipid accumulation and foam cell formation both at the systemic and renal levels28,36.
Additionally, ectopic kidney fat deposits have been associated with hypertension8. Thus, renal sinus fat is associated with stage II hypertension37. In the Framigham heart study, individuals with fatty kidneys (high perinephric fat levels) had a higher risk of hypertension (OR 2.12), which persisted after adjustment for body mass index (BMI) and visceral fat40.
Lipotoxicity and podocytes
Podocytes are the cells that modulate the glomerular size-selectivity barrier to proteins in the nephrons. Different slit diaphragm-associated proteins, such as nephrin, CD2-associated protein, podocin and alfa actin 4 contribute to maintain glomerular size-selective permeability30,34.
Gutwein et al. showed that chemokine CXCL-16 is the main scavenger receptor for ox-LDL in human podocytes, and is related with the production of another chemokine, CXCL12, which further impairs nephrin expression44.
Bussolati et al.46 showed that ox-LDL induces loss of nephrin expression in cultured podocytes. It seems that nephrin, when phosphorilated, causes the remodeling of the actin cytoskeleton and an increase in albumin diffusion across podocytes.
Sun et al.43 showed, in animal models of type 1 diabetes, that sterol regulatory element binding proteins (SREBP) are increased in the kidney cortex, resulting in upregulation of the enzymes responsible for FFA synthesis. Even with a normal serum lipid level, animals with SREBP-1 overexpression have elevated renal triglyceride levels, increased mesangial matrix expansion, proteinuria and glomerulosclerosis. These findings indicate that increased triglyceride accumulation in the intracelular mesangium may play a central role in the development of diabetic nephropathy.
Lipotoxicity and tubular cells
Oxidative processes are pivotal events in the injury of renal tubular epithelial cells exposed to ox-LDL (Figure 2c). CD36 are the main receptors for the uptake of ox-LDL in tubular renal cells36,39,41. Heme oxygenase, a redox-sensitive enzyme in proteinuric renal disease, is induced in epithelial cells following an exposure to ox-LDL and is accompained by cytotoxicity, in vitro. In fact, protection of cells with antioxidants resulted in a significant attenuation of cytotoxicity. A similarity was observed between the behaviour of endothelial cells and renal tubular epithelial cells after exposure to ox-LDL, concerning oxidant-mediated toxicity and expression of heme oxygenase. These findings suggest that the paradigm regarding the role of ox-LDL in atherogenesis may apply to tubular injury – ox-LDL causes tubulointerstitial fibrosis, also via atherosclerotic processes, which in turn lead to tubulointerstitial ischaemia7,9,23.
The mechanisms that explain the oxidation of LDL in proteinuric renal disease are not completely understood.
There are two hypotheses: (1) LDL is vulnerable to oxidative attack in the urinary space, from where LDL may be endocytosed by tubular cells or (2) LDL may be oxidized within tubular cells and in the microvascular bed which surrounds the tubulointerstitial tissue7,9,23.
THERAPEUTICS FOR LIPOTOXICITY
Statins
Statins lower cholesterol synthesis by inhibiting 3-hydroxy-3-methyglutaryl CoA (HMG-CoA) reductase and may have additional benefits on vascular function beyond their lipid lowering effect.
Statins also reduce AER and urinary loss of podocytes, a marker of severity of glomerular injury47.
Bussolati et al.46 showed, in vitro, that ox-LDL induced apoptosis in human podocytes and that this effect could be prevented by statins. Tonolo et al.48 showed that treatment with simvastatin maintains steady patterns of GFR, AER and nephrin expression.
Fenofibrate (potent PPAR-a agonist)
Hong et al.49 demonstrated, in animal models, the renoprotective effect of fenofibrate by ameliorating lipotoxicity in the kidney. It was already known that in diabetic nephropathy there is an increase in renal lipid accumulation, apoptotic renal injury and oxidative stress. The authors were able to relate these events to a decreased level of PPARa expression in diabetic mice which lead to the inactivation of AMPK-PGC-1a-ERR-1a, subsequently resulting in an increase in oxidative stress in the kidney.
Fenofibrate, by activation of AMPK-PGC-1a-ERR-1a signalling, reverses renal lipid accumulation and ameliorates diabetic nephropathy.
The PPARa activation leads to an increase in fatty acid catabolism and ATP production as well as a decrease in the levels of cytotoxic fatty acid peroxidation products, and promotes cell viability by inhibiting renal epithelium cell death. Furthermore, the authors demonstrated that fenofibrate directly activates AMPK by activation of PPARa, which prevents renal lipid accumulation and renal cell injury both in diabetic mice and in cultured mesangial cells treated with high glucose media49.
Tanaka et al.50 have also shown a protective effect of fenofibrate. It reduces glomerular lipid (mainly triglycerides) accumulation and oxidative stress associated with decreased albuminuria and glomerulosclerosis, on mice under a high-fat diet.
These results suggest that fenofibrate can be a potential therapeutic modality to treat type 2 diabetic nephropathy.
Polyphenols and flavonoids
Naturally occurring molecules, such as polyphenols and flavonoids, present in food and plant extracts, exhibit antioxidant and lipid lowering properties that prevent the effects of lipotoxicity.
Charradi et al.51 demonstrated the protective effect of grape seed extracts on nephrotoxicity induced by a high-fat diet in an animal model. The authors found that a high-fat diet provoked renal injury as evidenced by the rise in plasma urea and uric acid, creatinine clearance and proteinuria. Their data showed an increase of triglyceride deposition, intrarenal oleate and palmitate. Furthermore, there were positive correlations between hypertriglyceridemia and both the susceptibility to chronic kidney disease and the impact of dyslipidemia on the progression of chronic kidney disease. The supplementation with grape seed and skin extract (GSSE)52 resulted in a reduction of triglyceride deposition, copper depletion and kidney disfunction. The authors believe that GSSE acted directly on the kidney, and not indirectly as previously thought, by interfering with the process of weight and fat loss. Histological data also supported that GSSE is protective against proteinuria induced by a high-fat diet.
The specific polyphenols implicated in such protection have not yet been identified. Resveratrol53, quercetin or catechins as well as unsaturated fatty acids found in grape seed oil are potential candidates, although a synergism between these compounds is the most likely option54.
In conclusion, the use of an antioxidant could be beneficial in the prevention and treatment of lipotoxicity.
CONCLUSIONS
There is an epidemic of obesity and the metabolic syndrome across the world. Both are associated with high mortality mainly because of cardiovascular diseases.
Adipose tisse is an important endocrine organ and secretes different molecules that can communicate with every organ in the body.
Obesity is currently seen as a chronic low-grade inflammatory disease, which may spread to remote organs such as the liver, skeletal muscle and the pancreas to cause insulin resistance. Pro-inflammatory cytokines released by activated immune cells and adypocytes can impair insulin staging in insulin-responsive organs, promoting systemic insulin resistance, which increases the risk of developing type 2 diabetes.
Ectopic lipid accumulation in visceral tissues, including the kidneys, is a common finding in models of metabolic diseases but also in kidney diseases.
Increasing evidence suggests that it may contribute to kidney cell dysfunction and organ injury.
A strong relation between glucose and lipid metabolism has been recognized for decades. It has been proved that fatty acids can impair insulin stimulated glucose oxidation in muscles.
Excess intracellular non-esterified fatty acids and their metabolites (FA-CoA, DAG, and ceramides) promote insulin resistance and exert deleterious effects on various organs.
References
1. Abrass CK. Cellular lipid metabolism and the role of lipids in progressive renal disease. Am J Nephrol. 2004;24(1):46-53. [ Links ]
2. Alam MM, Meerza D, Naseem I. Protective effect of quercetin on hyperglycemia, oxidative stress and DNA damage in alloxan induced type 2 diabetic mice. Life Sci 2014;109(1):8-14. [ Links ]
3. Axelsson J, Aström G, Sjölin E, et al. Uraemic sera stimulate lipolysis in human adipocytes: role of perilipin. Nephrol Dial Transplant 2011;26(8):2485-2491. [ Links ]
4. Utzschneider KM, Kahn SE. Review: The role of insulin resistance in nonalcoholic fatty liver disease. J Clin Endocrinol Metab 2006; 91(12): 4753-4761. [ Links ]
5. Krssak M, Falk Petersen K, Dresner A, et al. Intramyocellular lipid concentrations are correlated with insulin sensitivity in humans: a 1H NMR spectroscopy study. Diabetologia 1999;42(1):113-116. [ Links ]
6. Charles MA, Eschwège E, Thibult N, et al. The role of non-esterified fatty acids in the deterioration of glucose tolerance in Caucasian subjects: results of the Paris Prospective Study. Diabetologia 1997;40(9):1101-1106. [ Links ]
7. Nosadini R, Tonolo G. Role of oxidized low density lipoproteins and free fatty acids in the pathogenesis of glomerulopathy and tubulointerstitial lesions in type 2 diabetes. Nutr Metab Cardiovasc Dis 2011;21(2):79-85. [ Links ]
8. Unger RH, Clark GO, Scherer PE, Orci L. Lipid homeostasis, lipotoxicity and the metabolic syndrome. Biochim Biophys Acta 2010;1801(3):209-214. [ Links ]
9. Samuel VT, Petersen KF, Shulman GI. Lipid-induced insulin resistance: unravelling the mechanism. Lancet 2010; 375(9733): 2267-2277. [ Links ]
10. Suganami T, Tanaka M, Ogawa Y. Adipose tissue inflammation and ectopic lipid accumulation. Endocr J 2012;59(10):849-857. [ Links ]
11. Feuerer M, Herrero L, Cipolletta D, et al. Lean, but not obese, fat is enriched for a unique population of regulatory T cells that affect metabolic parameters. Nat Med 2009;15(8):930-939. [ Links ]
12. Exley MA, Hand L, OShea D, Lynch L. Interplay between the immune system and adipose tissue in obesity. J Endocrinol 2014;223(2):R41-48. [ Links ]
13. Bonnard C, Durand A, Peyrol S, et al. Mitochondrial dysfunction results from oxidative stress in the skeletal muscle of diet-induced insulin-resistant mice. J Clin Invest 2008;118(2):789-800. [ Links ]
14. Lumeng CN, Bodzin JL, Saltiel AR. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J Clin Invest 2007;117(1): 175-184. [ Links ]
15. Eckel RH, Grundy SM, Zimmet PZ. The metabolic syndrome. Lancet 2005; 365(9468): 1415-1428. [ Links ]
16. Lumeng CN, DelProposto JB, Westcott DJ, Saltiel AR. Phenotypic switching of adipose tissue macrophages with obesity is generated by spatiotemporal differences in macrophage subtypes. Diabetes 2008; 57(12): 3239-3246. [ Links ]
17. Zhu D, Wang L, Zhou Q, et al. (+)-Catechin ameliorates diabetic nephropathy by trapping methylglyoxal in type 2 diabetic mice. Mol Nutr Food Res 2014;58(12):2249-2260. [ Links ]
18. Eckel RH, Alberti KG, Grundy SM, Zimmet PZ. The metabolic syndrome. Lancet 2010;375(9710):181-183. [ Links ]
19. Gustafson B, Smith U. Regulation of white adipogenesis and its relation to ectopic fat accumulation and cardiovascular risk. Atherosclerosis 2015;241(1):27-35. [ Links ]
20. Hue L, Taegtmeyer H. The Randle cycle revisited: a new head for an old hat. Am J Physiol Endocrinol Metabol. 2009 Sep; 297(3):E578-91. [ Links ]
21. Shulman, G.I. Ectopic fat in insulin resistance, dyslipidemia, and cardiometabolic disease. N. Engl. J. Med. (2014) 371, 2237–2238. [ Links ]
22. Szendroedi J, Yoshimura T, Phielix E, et al. Role of diacylglycerol activation of PKCΘ in lipid-induced muscle insulin resistance in humans. Proc Natl Acad Sci. USA 2014;111(26):9597-9602. [ Links ]
23. Lu J, Wang X, Wang W, et al. Abrogation of lectin-like oxidized LDL receptor-1 attenuates acute myocardial ischemia-induced renal dysfunction by modulating systemic and local inflammation. Kidney Int 2012;82(4):436-444. [ Links ]
24. Jornayvaz FR, Shulman GI. Diacylglycerol activation of protein kinase Cand hepatic insulin resistance. Cell Metab 2012;15(5):574-584. [ Links ]
25. Yu C, Chen Y, Cline GW, et al. Mechanism by which fatty acids inhibit insulin activation of insulin receptor substrate-1 (IRS-1)-associated phosphatidylinositol 3-kinase activity in muscle. J Biol Chem 2002;277(52):50230-50236. [ Links ]
26. Itani SI, Pories WJ, Macdonald KG, Dohm GL. Increased protein kinase C theta in skeletal muscle of diabetic patients. Metabolism 2001;50(5):553-557. [ Links ]
27. Garg A. Clinical review: Lipodystrophies: genetic and acquired body fat disorders. J Clin Endocrinol Metab 2011;96(11):3313-3325. [ Links ]
28. Karpe F, Dickmann JR, Frayn KN. Fatty acids, obesity, and insulin resistance: time for a reevaluation. Diabetes 2011;60(10):2441-2449. [ Links ]
29. Koppe L, Pelletier CC, Alix PM, et al. Insulin resistance in chronic kidney disease: new lessons from experimental models. Nephrol Dial Transplant 2014;29(9):1666-1674. [ Links ]
30. Cartwright MJ, Tchkonia T, Kirkland JL. Aging in adipocytes: potential impact of inherent, depot-specific mechanisms. Exp Gerontol 2007;42(6):463-471. [ Links ]
31. Lumeng CN, Saltiel AR. Inflammatory links between obesity and metabolic disease. J Clin Invest 2011; 121(6): 2111-2117. [ Links ]
32. Kume S, Uzu T, Araki S, et al. Role of altered renal lipid metabolism in the development of renal injury induced by a high-fat diet. J Am Soc Nephrol JASN 2007;18(10):2715-2723. [ Links ]
33. Kalbacher E, Koppe L, Zarrouki B, Pillon NJ, Fouque D, Soulage CO. Human uremic plasma and not urea induces exuberant secretion of leptin in 3T3-L1 adipocytes. J Ren Nutr 2011;21(1):72-75. [ Links ]
34. Sharma K, Ramachandrarao S, Qiu G, et al. Adiponectin regulates albuminuria and podocyte function in mice. J Clin Invest 2008;118(5): 1645-1656. [ Links ]
35. Montague CT, ORahilly S. The perils of portliness: causes and consequences of visceral adiposity. Diabetes 2000; 49(6): 883-888. [ Links ]
36. De Vries APJ, Ruggenenti P, Ruan XZ, et al. with the ERA-EDTA Working Group Diabesity. Fatty kidney: emerging role of ectopic lipid in obesity-related renal disease. Lancet Diabetes Endocrinol 2014;2(5):417-426. [ Links ]
37. Nielsen S, Guo Z, Johnson CM, Hensrud DD, Jensen MD. Splanchnic lipolysis in human obesity. J Clin Invest 2004;113(11):1582-1588. [ Links ]
38. Mraz M, Lacinova Z, Drapalova J, et al. The effect of very-low-calorie diet on mRNA expression of inflammation-related genes in subcutaneous adipose tissue and peripheral monocytes of obese patients with type 2 diabetes mellitus. J Clin Endocrinol Metab 2011;96(4):E606-613. [ Links ]
39. Merscher-Gomez S, Guzman J, Pedigo CE, et al. with the FinnDiane Study Group. Cyclodextrin protects podocytes in diabetic kidney disease. Diabetes 2013;62(11): 3817-3827. [ Links ]
40. Foster MC, Hwang S-J, Porter SA, Massaro JM, Hoffmann U, Fox CS. Fatty kidney, hypertension, and chronic kidney disease: the Framingham Heart Study. Hypertension 2011;58(5):784-790. [ Links ]
41. Sun L, Halaihel N, Zhang W, Rogers T, Levi M. Role of sterol regulatory elementbinding protein 1 in regulation of renal lipid metabolism and glomerulosclerosis in diabetes mellitus. J Biol Chem 2002;277(21):18919-18927. [ Links ]
42. Yu C, Chen Y, Cline GW, et al. Mechanism by which fatty acids inhibit insulin activation of insulin receptor substrate-1 (IRS-1)-associated phosphatidylinositol 3-kinase activity in muscle. J Biol Chem 2002;277(52):50230-50236. [ Links ]
43. Sun, L., Halaihel, N., Zhang, W., Rogers, T., and Levi, M. Role of sterol regulatory element-binding protein 1 in regulation of renal lipid metabolism and glomerulosclerosis in diabetes mellitus. J. Biol. Chem. (2002) 277, 18919–18927. [ Links ]
44. Gutwein P, Abdel-Bakky MS, Schramme A, et al. CXCL16 is expressed in podocytes and acts as a scavenger receptor for oxidized low-density lipoprotein. Am J Pathol 2009;174(6):2061-2072. [ Links ]
45. Meyer C, Gerich JE. Role of the kidney in hyperglycemia in type 2 diabetes. Curr Diab Rep 2002;2(3):237-241. [ Links ]
46. Bussolati B, Deregibus MC, Fonsato V, et al. Statins prevent oxidized LDL-induced injury of glomerular podocytes by activating the phosphatidylinositol 3-kinase/AKTsignaling pathway. J Am Soc Nephrol 2005;16(7):1936-1947. [ Links ]
47. Lee T-M, Su S-F, Tsai C-H. Effect of pravastatin on proteinuria in patients with wellcontrolled hypertension. Hypertension 2002;40(1):67-73. [ Links ]
48. Tonolo G, Velussi M, Brocco E, et al. Simvastatin maintains steady patterns of GFR and improves AER and expression of slit diaphragm proteins in type II diabetes. Kidney Int 2006;70(1):177-186. [ Links ]
49. Hong YA, Lim JH, Kim MY, et al. Fenofibrate improves renal lipotoxicity through activation of AMPK-PGC-1α in db/db mice. PLoS One 2014;9(5):e96147. [ Links ]
50. Tanaka Y, Kume S, Araki S, et al. Fenofibrate, a PPAR1α agonist, has renoprotective effects in mice by enhancing renal lipolysis. Kidney Int 2011;79(8): 871-882. [ Links ]
51. Charradi K, Elkahoui S, Karkouch I, et al. Grape seed and skin extract alleviates highfat diet-induced renal lipotoxicity and prevents copper depletion in rat. Appl Physiol Nutr Metab 2013;38(3):259-267. [ Links ]
52. Zhang Z, Li BY, Li XL, et al. Proteomic analysis of kidney and protective effects of grape seed procyanidin B2 in db/db mice indicate MFG-E8 as a key molecule in the development of diabetic nephropathy. Biochim Biophys Acta 2013;1832(6):805-816. [ Links ]
53. Xu F, Wang Y, Cui W, et al. Resveratrol prevention of diabetic nephropathy is associated with the suppression of renal inflammation and mesangial cell proliferation: Possible roles of Akt/NF-kB pathway. Int J Endocrinol 2014;2014:289327. [ Links ]
54. Kim MY, Lim JH, Youn HH, et al. Resveratrol prevents renal lipotoxicity and inhibits mesangial cell glucotoxicity in a manner dependent on the AMPK-SIRT1-PGC1α axis in db/db mice. Diabetologia 2013;56(1):204-217. [ Links ]
Drª Ana Rita Mateus Martins
Department of Nephrology, Hospital Santa Cruz,
Centro Hospitalar de Lisboa Ocidental
Avenida Professor Reinaldo dos Santos, 2790-240 Carnaxide
Oeiras, Portugal.
E-mail: anarita.mateus@gmail.com
Conflict of interest statement: None declared.
Received for publication: 25/05/2015
Accepted in revised form: 07/10/2015

{kind=link}
{kind=link}