









Services on Demand
Journal
Article
 Portuguese (pdf)
Portuguese (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkGOT, Revista de Geografia e Ordenamento do Território
On-line version ISSN 2182-1267
GOT no.8 Porto Dec. 2015
https://doi.org/10.17127/got/2015.8.008
ARTIGO ORIGINAL
Organização geossistêmica e invasões por Acacia sp. (Fabaceae: Mimosoideae) na Bacia Hidrográfica do Rio Arouce. Uma visão das invasões biológicas à escala do geossistema.
Geosystems and invasions by Australian Acacia Species (Fabaceae, Mimosoideae) in the Rio Arouce’s watershed area. An overview of biological invasions at the geosystem scale.
Oliveira-Costa, Jorge1; Souza, Reginaldo2
1Faculdade de Letras/Universidade de Coimbra. Largo da Ponte Férrea, 3004-530, Coimbra, Portugal. oliveiracostajorge@gmail.com
2Colegiado de Geografia, Universidade Federal da Fronteira Sul - Erechim/Rio Grande do Sul. ERS 135 - Km 72, nº 200, Erechim/RS, Brasil. regionaldogeo@hotmail.com
RESUMO
Vários fatores influenciam os alcances espaciais e temporais das invasões biológicas. A estrutura da paisagem é um dos fatores cruciais, que influenciam os alcances, assim como os elementos de risco. Por isso, a avaliação de áreas invadidas por espécies exóticas implica a análise da estrutura da paisagem. O presente estudo é abordado por meio da análise da estrutura da paisagem no passado (1960) e no presente. O método aplicado inclui análise de imagens orbitais e de cartografias do uso do solo. O período de tempo abrange 50 anos, 1960-2010. A área de estudo está localizada na Serra da Lousã (Portugal). Os resultados mostraram claras mudanças no uso da terra, como estímulo para aumento das áreas invadidas, em contextos de propensão à invasão, com destaque para as áreas invadidas por Acácias Australianas, que parecem aumentar de forma constante.
Palavras-chave: Mudanças no uso do solo, Invasão biológica, Análise espacio-temporal, Superfícies invadidas, Bacia Hidrográfica do Rio Arouce, Género Acácia.
ABSTRACT
Various factors influence the spatial and temporal spreads of biological invasions. Land structure is one of the crucial factors, which influence the spreads, but also risk elements. Therefore, the assessment of invaded areas by alien species implies the analysis of land structure. This study is approached by analyzing the past and present of the land structure. The applied methods include analysis of orbital images and land cover maps. The timespan covers 50 years, 1960-2010. The study area is located in the ‘Serra da Lousã’ (Portugal). The results show clear changes in the land use, with trends to increase in invaded areas. A prone to invasion was detected, where are highlighting Acacia areas that seem to increase steadily.
Keywords: Land use change, Biological invasion, Spatial-temporal analysis, Invaded surfaces, ‘Bacia Hidrográfica do Rio Arouce’ (Portuguese watershed), Acacia genus.
1. Introdução
Avaliar parâmetros geográficos da invasão biológica é essencial para ajudar a explicar porque algumas espécies exóticas tornam-se invasoras (Williamson, 1996; Lockwood et al., 2007; Simberloff & Rejmànek, 2011). O homem tem investigado o fenómeno das invasões biológicas há mais de 100 anos (Vavilov, 1992; Cronk & Fuller, 1995; McNelly, 2001; Davis, 2006; Lockwood et al., 2007; Vilà et al., 2008), desde um primeiro experimento em 1882 sobre a invasão por Mangifera indica na Jamaica (Espínola et al., 2007). Entretanto, desde este pioneirismo, o número de espécies invasoras, os graus de magnitude dos processos das invasões biológicas, e os caminhos teórico-metodológicos dos trabalhos, têm mudado significativamente, sobretudo nas últimas cinco décadas, após publicação de Charles Elton (Simberloff & Rejmànek, 2011), um dos pioneiros no estudo das relações de invasão, que em 1958 organizou as idéias anteriores definindo a invasão biológica como um processo de degradação dos ecossistemas resultado do estabelecimento de populações exóticas autônomas, após a tranferência de indivíduos (por ação humana) geralmente para áreas que guardam semelhanças (clima, solo, relevo, altitude, hidrografia) com o local de origem (Cronk & Fuller, 1995; Pyšek et al., 1995; Williamson, 1996).
O fenómeno das invasões biológicas é considerado pela IUCN (World Conservation Union) (1999) uma das principais causas de perda da diversidade biológica, apresentando-se atualmente como um dos principais desafios à gestão do território, principalmente pelas elevadas perdas económicas ou custos associados ao controlo que este processo pode implicar (Pimentel et al., 2005). A Ciência da Invasão é hoje componente importante da teoria ecológica, que, por sua vez, é componente importante da teoria geográfica, sendo importantes aportes para explicação de porque o comportamento das espécies fora do seu alcance original é tão diverso e porque sua organização espacial é complexa.
As vias com que diferentes espécies exóticas invasoras têm sido introduzidas e o comportamento destas espécies no novo ambiente tem se tornado altamente complexos e mudam com o tempo e o espaço (Shigesada & Kawasaki, 1997; Vitousek et al., 1997; Cadotte et al., 2006). Desse modo, o fenómeno das invasões congrega graus (espécie exótica – exótica casual – naturalizada – invasora – transformadora) e etapas (introdução – colonização/estabelecimento – naturalização/expansão) no contexto de uma fase inicial lenta de estabelecimento dos indivíduos, seguida de uma fase exponencial, do que pode resultar no sucesso ou no insucesso da invasão, tornando as espécies invasoras ou não (Pyšek et al., 1995; Williamson, 1996). O sucesso da invasão, em geral, acarreta na modificação dos ecossistemas e na exclusão de espécies nativas como consequência do fenómeno da homogeneização biótica (McNelly, 2001). Este estado de transição, entre sucesso e insucesso, depende dos caracteres biológicos intrínsecos aos organismos envolvidos (Cronk & Fuller, 1995) e da conjuntura do habitat invadido (Cadotte et al., 2006), sobretudo nos termos da homogeneidade da estrutura espacial do recurso disponível e perturbações, do que podem resultar impactos (Henderson et al., 2006; Vilà et al., 2013).
Em particular, o sucesso da invasão está de perto correlacionado com a potencial pressão de propágulos (Cronk & Fuller, 1995), numa combinação do número de vezes e locais em que a espécie é introduzida e o número de indivíduos ou propágulos em cada introdução, onde uma pequena proporção (cerca de 10%) das espécies introduzidas se naturalizam, e destas, outra pequena proporção (cerca de 10%) se tornam invasoras (Pyšek et al., 1995; Williamson, 1996). A existência de informações apenas do sucesso da invasão tem permitido que os impactos diretos e indiretos causados pelas invasões biológicas ainda não estejam bem esclarecidos (Levine et al., 2003; Henderson et al., 2006; Vilà et al., 2013).
Por sua dimensão global, o fenómeno das invasões biológicas tem recebido maior atenção, com interesse de pesquisadores de diversas áreas (ecólogos, biólogos, zoólogos, geógrafos), e estudos que compreendem desde a estrutura da paisagem (Henderson et al., 2006), até exame das características fenotípicas e funcionais da espécie exótica (Pyšek et al., 1995). Mesmo que os impactos diretos e indiretos causados pelas invasões biológicas ainda não estejam bem esclarecidos (Levine et al., 2003; Vilà et al., 2013), tem emergido certezas como: (i) espécies invasoras podem reduzir as comunidades nativas com interferências no crescimento, nos valores funcionais e fenológicos das espécies nativas (Shigesada & Kawasaki, 1997; Vitousek et al., 1997; Cadotte et al., 2006); (ii) espécies invasoras podem alterar o funcionamento dos ecossistemas, com interferências na ciclagem de nutrientes, no balanço hídrico, na produtividade, nos processos sucessionais, e na diversidade biológica (Henderson et al., 2006; Lockwood et al., 2007). Cronk & Fuller (1995) ligam as seguintes características das espécies invasoras a seus impactos em áreas não nativas: (i) a larga produção de sementes com alta viabilidade e longevidade, (ii) o rápido crescimento do sistema radicular, (iii) a adaptação ao fogo, (iv) a habilidade de interferir no crescimento de plantas vizinhas, (v) a alta produção de propágulos, (vi) a similaridade morfológica e fisiológica com as espécies nativas, (vii) a autopolinização pelo sistema reprodutivo, e (viii) a habilidade de fixação de nitrogênio por bacterias simbiontes.
Assim como a teoria ecológica, a teoria geográfica também proporciona oportunidades para avaliação dos estímulos que promovem as invasões biológicas (Oliveira-Costa, 2014). O geossistema (Sochava, 1977) e seu caráter de zoneamento com determinação da escala geográfica inscrita nos padrões espaciais da distribuição dos fenómenos, pode constituir numa importante ferramenta para avaliação da complexidade e dinâmica sistêmica do fenómeno da invasão, além dos padrões espaciais e as relações desse fenómeno com os caracteres estruturadores da paisagem. A aplicação de estudos geossistêmicos para diagnósticos ambientais está bem representada a nível mundial (Chorley & Haggett, 1974; Stoddart, 1974; Sochava, 1977; Tricart, 1977; Bertrand, 1968), entretanto, sua aplicação no estudo da invasão por espécies exóticas invasoras é inédita, o que pode constituir numa importante contribuição para esse campo de estudo. O geossistema (Bertrand, 2002) foi proposto como modelo teórico integrador para o estudo das dinâmicas da natureza, a partir de uma leitura geográfica do modelo ecossistêmico no âmbito dos estudos ecológicos.
O modelo geossistêmico (Bertrand, 1968) compareceu como possibilidade teórica e metodológica para analisar as transformações da paisagem através do tratamento das diversas características de sua estrutura e funcionamento, desde o embasamento rochoso à cobertura vegetal e interações climáticas – de modo vertical – às distribuições de fisionomias que compõem a tessitura do espaço geográfico e que são influenciadas por impactos da ação do homem – de modo horizontal. Desta forma, coexistem duas frentes analíticas no modelo geossistêmico: uma vertical e outra horizontal (Stoddart, 1974). Os fluxos de matéria e energia entre estas duas dimensões são considerados como ativadores das transformações paisagísticas no espaço e no tempo (Bertrand, 2009).
Mesmo com evidente inspiração no conceito de ecossistema (Tricart, 1977), o geossistema (Sotchava, 1977) resguarda algumas particularidades que fazem com que ele seja mais interessante do ponto de vista de uma abordagem geoecológica da natureza. Exemplos disso seriam a escala de análise e a definição mais objetiva de unidades de paisagem (Bertrand, 2002). Para o desenvolvimento de uma análise geoecológica das dinâmicas da natureza e dos impactos motivados pela ação antrópica, tal dimensionamento teórico se apresenta como contributo aos estudos transversais do meio ambiente e de diferentes problemáticas que contemplam a questão ambiental na contemporaneidade, como é o caso do fenómeno das invasões biológicas (Oliveira-Costa, 2014).
No presente estudo nós apresentamos uma estrutura para convergência entre o Modelo Teórico do Geossistema (Bertrand, 1968) e o Modelo Teórico da Invasão (Williamson, 1996). Este artigo se apresenta como proposta metodológica que visa o estabelecimento de uma base teórica para o estudo da paisagem invadida, na perspectiva, portanto, daquilo que compreendemos como ‘Geoecologia’. Nosso aporte empírico foi desenvolvido com base num estudo de caso em área invadida na Região Centro de Portugal. Com este artigo buscamos proporcionar oportunidades para aplicabilidade do modelo teórico da invasão combinado ao modelo do geossistema: 1) as escalas espacio-temporais nas quais a invasão se apresenta dentro do geossistema e como estas mudam sob diferentes circunstâncias; 2) os caracteres da estrutura da paisagem como estímulos para diferenças nos graus temporo-espaciais das infestações; 3) a contribuição dos dois modelos teóricos para entendimento da complexidade do fenómeno. A múltipla dimensão desse esboço teórico-metodológico poderá sugerir novas abordagens para o jovem campo da Invasão Biológica.
2. Estudo de caso e aplicação da metodologia
2.1. Área de estudo
A unidade de estudo é uma bacia hidrográfica, com extensão de 7.300 hectares. A Bacia do Rio Arouce (figura 1) está concentrada no Concelho da Lousã (Região Centro de Portugal), na justaposição entre as bacias hidrográficas dos rios Mondego e Tejo. A topografia é caracterizada por transição de cotas altimétricas entre 200-1000m, e graus de declividade entre 0-48 graus, com dominância de superfícies declivosas concentradas espacialmente nos setores intermédio e cabeceiras da bacia, e superfícies aplainadas em erosão (quer natural quer humana) concentradas no setor terminal da bacia, no limite com o Rio Ceira.
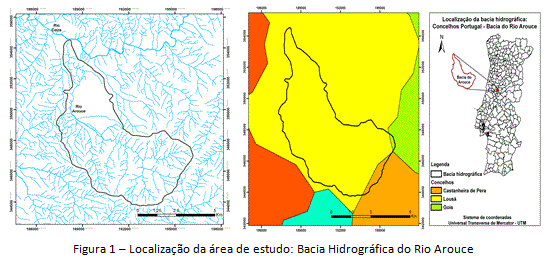
O principal tipo de solo na área de estudo é o cambissolo, do tipo húmico. A litologia é composta de formações sedimentares-metamórficas (areias, cascalhos, arenitos, argilas) distribuídas pelos setores intermédio, cabeceiras e terminal da bacia. Geologicamente, é verificada dominância de rochas do câmbrico-précâmbrico (xistos e grauváques), período de formação do complexo xisto-grauváquico. A estrutura da paisagem é dominada por culturas arvenses e formações florestais, inscritas num contexto de regime hídrico de surperávit para o ano inteiro, com clima do tipo húmido. As áreas sociais (urbanas e rurais) estão concentradas principalmente ao longo do setor da calha central do Rio Arouce, com as áreas residenciais, prédios públicos e áreas industriais distribuindo-se pela planície sedimentar, e as áreas rurais e áreas urbanas (descontínuas) distribuindo-se pelo Vale do Rio Arouce.
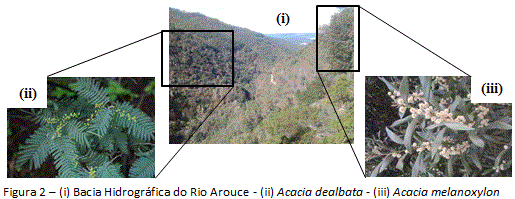
2.2. Espécies focais
As duas espécies de Acácia que ocorrem na Bacia do Rio Arouce foram selecionadas para este estudo (figura 2). Correspondem a plantas leguminosas nativas do sudeste da Austrália, com crescimento que pode alcançar até 15m. Estão entre as espécies do género Acacia (Tourn.) Mill. com maior infestação em Portugal (Marchante et al., 2008). As Acácias foram introduzidas no país há quase 200 anos para controlo da erosão (Fernandes, 2012), entretanto, após poucas décadas no novo ambiente, se dispersaram extensivamente, particularmente por estímulos (ex: uso do solo), com prejuízos para a comunidade nativa. O estabelecimento de simbiose (Rhizobia) com fixação de nitrogênio da atmosfera é uma das principais características das espécies focais (Richardson et al., 2011). A larga produção de sementes com alta viabilidade/longevidade, a adaptação ao fogo, e a autopolinização, são fatores atribuídos ao sucesso das Acácias Australianas em áreas não nativas.
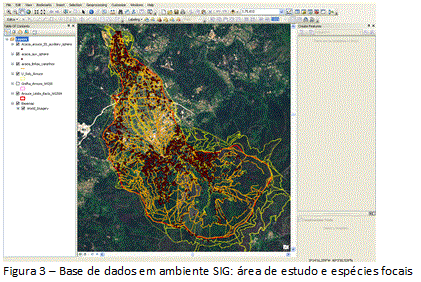
2.3. Tratamento dos dados regionais
Em Sistema de Informação Geográfica, identificou-se a presença/abundância das Acácias, apresentando a distribuição espacial e os graus da infestação (figura 3). A pesquisa foi iniciada em gabinete, com fotointerpretação de imagens orbitais de alta resolução (resolução 60cm) na escala de 1/25000, e preparação das cartografias de uso do solo para diferentes momentos cronológicos, com homogeneização dos produtos cartográficos relativos a 1960 (Carta Agrícola e Florestal de Portugal, 1/25000) e 2010 (imagem orbital BASEMAP, ArcGIS 10.1). Assim, foi produzida cartografia base contendo as unidades geossistêmicas da bacia e os graus de ocupação e sociabilidade das espécies invasoras.
A caracterização do ambiente da Bacia do Rio Arouce foi realizada segundo suas unidades geossistêmicas, identificando a importância dos fatores de ordem antrópica (usos do solo) e natural (hidrologia, tipo de relevo, clima) na condução do processo das invasões. Para avaliação da evolução espacio-temporal da superfície invadida por Acácias, foi estabelecido recorte de 50 anos, a partir das cartografias disponíveis (de 1960 a 2010).
Foram realizadas etapas de campo para posicionamento geográfico com utilização de GPS (sigla em inglês, Global Positioning System) e recolhimento de informações da presença e abundância dos taxa selecionados. Para isso se utilizou uma grelha de referência 200x200m, onde foram atribuídos parâmetros arbitrários quantitativos (1-5) e qualitativos (1-3) relacionados ao grau de ocupação (<5%; 5-25%; 25-50%; 50-75%; >75%) e sociabilidade (indivíduos isolados, pequenas manchas, grandes manchas) das populações invasoras.
2.4. O modelo teórico da Invasão Biológica
A base conceitual desse modelo baseia-se no princípio teórico da tolerância das espécies aos filtros dos habitats, onde são distinguidas duas formas de distribuição dos seres vivos: a distribuição em tipos particulares de ecossistemas por espécies tolerantes aos filtros (=nativa; =autóctona; =indígena); e a distribuição procedente de outras áreas geográficas, a partir da introdução humana (=introduzida; =exótica; =alóctona; =não nativa; =importada). A invasão dependerá de fatores como mecanismos de dispersão, distância do local de origem, e barreiras geográficas, sendo resultado da hierarquia iniciada pelo (a) fator geográfico como filtro biogeográfico, seguida do (b) fator ecológico como filtro fisiológico, do (c) fator ‘caracteres da espécie’ como filtro biótico, e do (d) fator ‘introdução antrópica’.
O modelo congrega, pelo menos, quatro graus, do que pode resultar no sucesso da invasão. Num primeiro grau, o estabelecimento e a reprodução dos indivíduos acontecem sem intervenção humana, mas a persistência destes dependerá da proximação de zonas antropizadas (=adventícias; =subespontâneas; =casuais). Num segundo grau, os indivíduos são capazes de formar populações autônomas no local da introdução (=naturalizadas; =estabelecidas). Num terceiro grau, espécies naturalizadas rapidamente se expandem para longe do foco da introdução, com populações estáveis de numerosos indivíduos (=invasoras). Num quarto grau, espécies invasoras são capazes de alterar os ecossistemas, com danos em infraestruturas, no desaparecimento de nativas, troca do ciclo de nutrientes e na transmissão de doenças (=pestes; =transformadoras; =pragas) (Williamson, 1996).
Estes quatro graus estão inseridos numa sequência de três etapas: 1º introdução (movimentação de espécies potencialmente invasoras pelo homem para outro ecossistema onde não são encontradas); 2º estabelecimento (processo em que uma espécie produz com êxito indivíduos viáveis conforme a probabilidade de sobrevivência); 3º expansão (disseminação e alcance das espécies exóticas no novo ambiente). O modelo teórico define como (i) ‘contaminação biológica’ quando o estabelecimento de exóticas causa mudanças no funcionamento do ecossistema das quais este não consegue se recuperar naturalmente; (ii) ‘capacidade invasora’ ou ‘invasividade’ refere-se aos traços biológicos que potencializam o sucesso do estabelecimento e expansão da espécie; (iii) ‘susceptibilidade à invasão’ ou ‘invasibilidade’ refere-se aos aspectos que caracterizam a fragilidade do ambiente a recetividade de espécies exóticas, na dependência dos caracteres do meio físico.
2.4.1. O sucesso da introdução
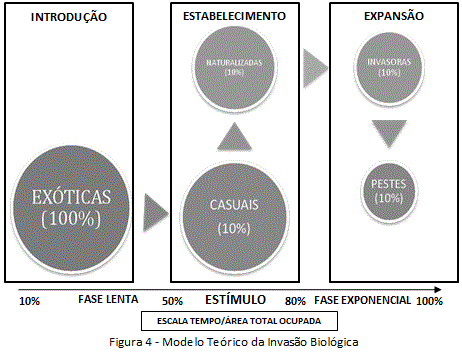
O modelo teórico da invasão biológica é subsidiado por informações detalhadas, sobretudo, no âmbito de introduções que tiveram sucesso. Por isso, foi proposta uma regra geral das condições para ser considerado o sucesso da introdução (figura 4): (1) na fase lenta, ou de dispersão pontual (menos de 50 anos da introdução), a espécie deve estar estabelecida a, pelo menos, 100 metros do foco de entrada, e (2) na fase exponencial, ou de dispersão difusa (mais de 50 anos da introdução), a espécie deve estabelecer-se a mais de 100 metros do foco de entrada (regra nomeadamente para espécies com reprodução por semente). Para o caso de espécies com reprodução por rizomas ou estolões, estas devem se estabelecer a mais de 6 metros, em até três anos após a introdução (Williamson, 1996).
2.4.2. Surgimento de estímulos e habitat disponível
Por basear-se em introduções que tiveram sucesso, o modelo impede o entendimento de muitos fatores que controlam a invasão. Este fato tem feito emergir, pelo menos, três certezas: (i) o sucesso da invasão está de perto ligado ao surgimento de estímulos e habitat disponível, no âmbito de aspectos relacionados à espécie (ex: pressão de propágulos - nº de vezes, locais e indivíduos por cada introdução), e a estrutura da paisagem (no âmbito do potencial ecológico, exploração biológica, e ação antrópica); (ii) em geral, apenas 10% das espécies introduzidas se naturalizam, e destas, apenas 10% tornam-se invasoras (Tens Rule); (iii) a invasão caracteriza-se como um processo de rápida expansão e estabelecimento de populações autonômas e numerosas, com impactos significativos (Williamson, 1996).
2.4.3. Os graus da infestação
A Regra dos 10% - Tens Rule (figura 4) caracteriza a invasão como um fenómeno ecológico progressivo, e não unidirecional, na dependência dos caracteres da espécie (invasividade) e fatores ambientais (invasibilidade). Por isso, não há uma proporção constante de espécies que passem de um grau para o outro. A percentagem aproximada de espécies exóticas que se tornam invasoras é de 0,1%, já que, segundo a Tens Rule, 10% das espécies exóticas passam a exóticas casuais, 10% das exóticas casuais se naturalizam, 10% das naturalizadas se tornam invasoras, e 10% das invasoras tornam-se transformadoras (Williamson, 1996).
2.5. O modelo teórico do Geossistema
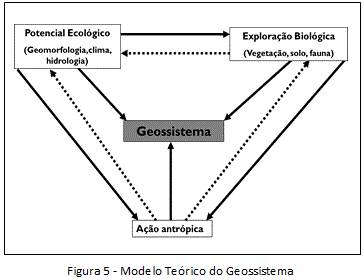
Bertrand (1968) apresentou o geossistema (figura 5) como esboço metodológico para análise das dinâmicas naturais e da degradação antrópica. Aquele pesquisador demonstrou em 1968 as escalas espaciais da análise geográfica, que podem variar de alguns metros a centenas de quilômetros quadrados: geossistema, geofácies, geótopos.
2.5.1. Organização geossistêmica (espacial)
Os geofácies formam um mosaico caracterizado pela presença de diversas unidades de paisagem cujos fluxos de matéria e energia definem as dinâmicas da vegetação – regressão, progressão ou equilíbrio – e do relevo – prevalência da morfogênese ou pedogênese. Por serem definidas como unidades no interior de um quadro paisagístico mais amplo, os estados dos geofácies (degradação ou conservação) refletirão as próprias condições do impacto antrópico no meio ambiente.
Por sua escala, os geótopos não devem ser considerados paisagens, mas, elementos que participam de uma combinação espacial mais ampla. Os geótopos conduzem a análise das microformas presentes no terreno – olhos d’água, vertentes com sulcos erosivos, condições fitofisiomômicas diferentes do entorno. Os geótopos agregam a espacialidade das interações entre biótopos e biocenoses, dos refúgios de biocenoses originais, relictuais ou endêmicas. As dinâmicas dos geótopos são muito particulares. Embora recebam influências das interações em escalas maiores, seu estado pode ser diferente do geossistema.
Ao conjunto dos geofácies e dos geótopos, constantemente modificados pela ação da natureza e/ou da interferência humana, denomina-se geocomplexo. O geossistema (geofácies, geótopos, geocomplexo) foi definido como unidade espacial mais interessante à pesquisa por conter dimensões pelas quais os impactos aparecem mais diretamente ao olhar do pesquisador. A teoria geossistêmica diz respeito a uma entrada – naturalista – na análise da paisagem, de modo que, em sua materialidade de formas, fluxos e relações entre os elementos bióticos, abióticos e ação antrópica, as unidades constituídas em geótopos e geofácies representam os diferentes estágios de evolução dos geocomplexos.
O potencial ecológico se define pela geologia, geomorfologia, clima e hidrografia. Trata-se de uma caracterização do meio ambiente a partir das interações entre estas dimensões, retratando a particularidade do conjunto abiótico do geocomplexo regional. A exploração biológica diz respeito ao modo pelo qual a dimensão biótica se espacializa no potencial ecológico e como se dão as relações. Trata-se de uma entrada naturalista para entender os fatores que engendraram certa distribuição espacial da vegetação, da fauna e suas influências nos processos pedogenéticos, uma vez que os solos são compreendidos como interfaces entre a esfera viva e não viva, seus compostos minerais e a matéria orgânica.
Geossistemas são sistemas abertos nos quais a ação antrópica é um input de matéria e energia nas cadeias de interações entre seus elementos. No estudo integrado das dinâmicas socioambientais, compreende-se que o desempenho dos elementos naturais na elaboração da paisagem sofre intereferências do modo de atuação do homem na criação e recriação de composições híbridas (a cidade, a fazenda, o parque, a área de floresta “protegida”, o controle das águas, entre outras...). Portanto, devem-se compreender as mudanças advindas da antropização da natureza e perturbações desse sistema, para se analisar as dinâmicas de transformação e os diferentes estados de um geossistema.
2.5.2. Organização geossistêmica (temporal)
Os estados de um geossistema também se relacionam com os ritmos desse geossistema, dados a partir de diversas escalas temporais. Pelo fato de geocomplexos, geofácies e geótopos terem suas próprias dinâmicas, todos se encontram em estado de equilíbrio relativo: tanto os processos naturais, quanto aqueles influenciados pela ação antrópica, transformam constantemente a paisagem regional. Bertrand (2009) apresenta uma escala com sete grandezas temporais, conforme o quadro seguinte (quadro 1).
A primeira escala de análise parte dos ritmos de observação direta dos geocomplexos. A última escala permite a análise plurianual. As grandezas intermediárias são relevantes para o conhecimento detalhado dos ritmos naturais - ora mais, ora menos alterados pela ação antrópica - de um determinado espaço geográfico.
Neste momento, parte-se da hipótese de que a convergência desta proposta teórico-metodológica, na sua dimensão espacial e temporal, com o modelo da invasão biológica, pode ser uma interessante estratégia de análise das dinâmicas ambientais que ativaram e ativam as transformações da paisagem na Bacia do Rio Arouce, levando-se em consideração o atual estágio de geossistemas constituidos por geofácies cuja homogeneidade fisionômica é marcada pela presença das espécies invasoras Acacia dealbata e Acacia melanoxylon.
3. Resultados e discussões
3.1. O sucesso da introdução (análise nível generalista I)
A investigação sobre o sucesso da introdução das Acácias na área da Bacia Hidrográfica do Rio Arouce seguiu os encaminhamentos fundamentais do Modelo Teórico da Invasão. Foi estabelecido recorte temporal de 50 anos, em análise com base na fotointerpretação de imagens orbitais e sua homogeneização com cartografias do uso do solo, avaliando as mudanças do início da fase exponencial (1960 - Carta Agrícola e Florestal de Portugal/CAF) à fase exponencial atual das invasões (2010 - imagem BASEMAP, ArcGIS 10.1). Para efeito dessa avaliação, a estrutura da paisagem da Bacia Hidrográfica do Rio Arouce foi sistematizada em nove categorias em nível generalista das opções de uso/ocupação do solo: 1) Área social; 2) Cultura de sequeiro; 3) Cultura de regadio; 4) Olival/Pomar; 5) Floresta de resinosas; 6) Floresta de eucaliptos; 7) Floresta de folhosas; 8) Incultos; e 9) Acacial.
O primeiro mapeamento das Acácias em Portugal (1980) identificou povoamentos dispersos na Bacia do Arouce (figura 6), 20 anos depois do registro da presença de indivíduos isolados. Segundo as conceituações do Atlas da Distribuição das Acácias e Eucaliptos (SROA), o povoamento disperso não constitui povoamento propriamente dito, pois a densidade de indivíduos não atinge número suficiente. Desse modo, foram identificadas pequenas manchas, o que indica que em 1980 a invasão estava no início da fase exponencial, em etapa de expansão. É identificado o possível local da introdução (unidade intermédia), onde o registro parece indicar o começo da dispersão para longe do foco de entrada (figura 6).
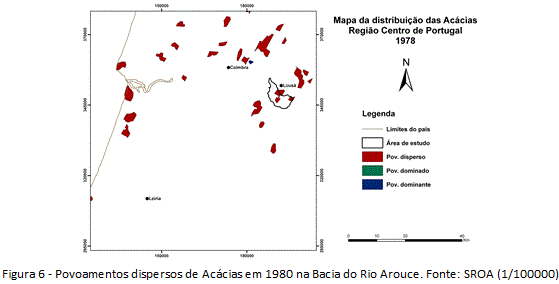
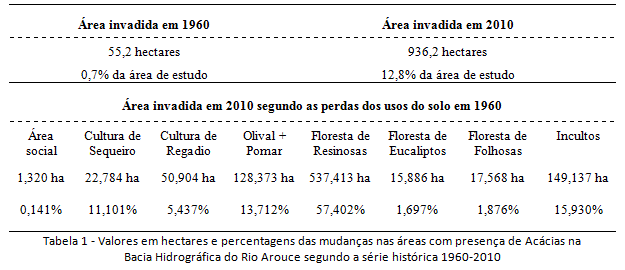
Em razão da indisponibilidade de cartografias para recortes temporais pretéritos, não foi possível estabelecer o momento da introdução das Acácias na área da Bacia do Rio Arouce. Estima-se que tenha ocorrido há mais de 100 anos (seguindo o Modelo Teórico da Invasão), na unidade intermédia dada a densidade elevada das populações nessa unidade da bacia. Ainda, estima-se que as populações, conduzidas pelo surgimento de estímulos, tenham começado a aumentar a partir de 1960, caracterizando o período anterior como a fase lenta e de dispersão pontual das espécies, e o recorte 1960-2010 como a fase exponencial e de dispersão difusa. Tendo a área invadida em 2010 ultrapassado 900 hectares (mais de 12% da área de estudo), enquanto que em 1965 esteve reduzida a apenas 13 hectares (menos de 1% da bacia) (tabela 1), e sabendo da existência em 1980 de povoamentos dispersos, estimam-se três etapas distintas da invasão no período 1960-2010: (i) etapa de estabelecimento (1960 – grau naturalizado), terá sido o final da fase lenta e início da fase exponencial; (ii) etapa de expansão inicial (1980 – grau invasor), terá sido o início da fase exponencial propriamente dita; (iii) e etapa de expansão potencial (2010 – grau transformador), concluindo o sucesso da introdução. Entre as mudanças no uso do solo na área de estudo que estão de perto relacionadas ao sucesso da introdução, destaca-se a diminuição das áreas com Floresta de resinosas (tabela 1). Mais de 50% da área atual de Acácias eram ocupadas por Floresta de resinosas em 1960 (tabela 1).
3.2. Surgimento de estímulos e habitat disponível (análise nível generalista II)
Para avaliação do surgimento de estímulos e habitat disponível entre 1960-2010 na área de estudo, foram homogeneizadas as imagens orbitais de 2010 de alta resolução (BASEMAP), com as cartografias de uso do solo para o recorte temporal de 1960 (CAF), quantificando a evolução das áreas segundo as nove categorizações de opções de uso do solo propostas.
Observado os acréscimos e decréscimos dos tipos de usos entre 1960-2010 na Bacia do Rio Arouce (figuras 7 e 8), o uso do solo foi clarificado como estímulo para a invasão local dada as mudanças significativas que vem ocorrendo na estrutura da paisagem desde 1960. O habitat disponível para as invasões tiveram acréscimo, sobretudo, nas unidades terminal e intermédia da bacia, o que inclinou essas unidades a processos de invasão. Num âmbito geral da estrutura da paisagem, as mudanças significativas foram de perda de usos tradicionais (figura 9). Os decréscimos de 15% nas áreas agrícolas, nomeadamente aquelas com ocorrência nas unidades terminal e intermédia junto ao Rio Arouce (Regadio e Olival/Pomar, por exemplo), permitiram perdas que resultaram em quase 20% da área invadida em 2010 (tabela 1). Já quanto aos usos florestais, representando as maiores mudanças no uso do solo, os decréscimos de 15% nas áreas com Florestas de resinosas (figura 9) terão sido determinantes na disponibilidade de habitat, com perdas da área original que correspondem a quase 60% da área atual ocupada pelas Acácias (tabela 1).
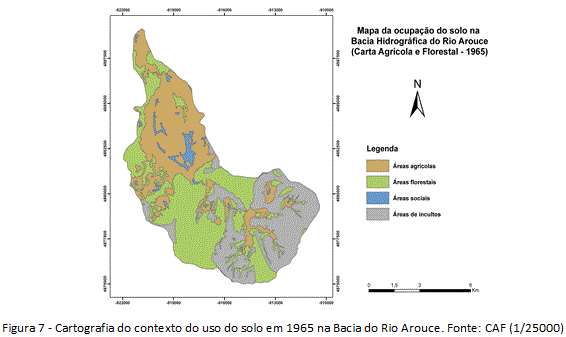
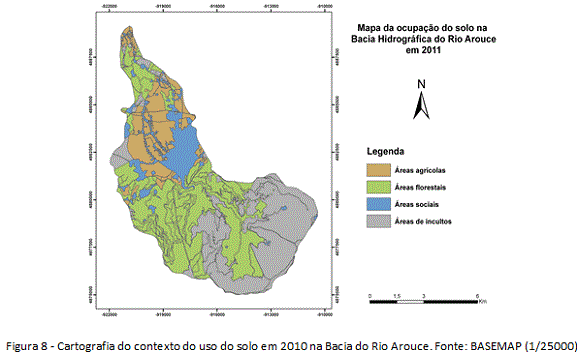
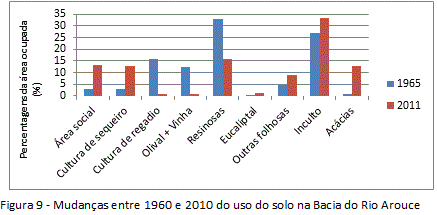
As perdas de usos tradicionais (agrícolas/florestais), que eram maioria em 1960, produziram contextos de inclinação à invasão, estimulando o aumento do processo desde então. Quanto aos usos do solo que ganharam áreas em 2010, destacam-se as culturas agrícolas de sequeiro (+10%) e as áreas sociais (+10%) (figura 9). O aumento das culturas de sequeiro pode estar relacionado ao caráter extensivo desse uso (oposição à cultura de regadio), não seguindo a tendência nacional de diminuição das áreas agrícolas (Nunes et. al. 2007, 2010). Sobre o aumento das áreas sociais, faz necessário destacar o aumento das vias de comunicação (figura 8), que, diferentemente do aumento da cultura de sequeiro, tem implicações diretas como estímulo a invasão. Os incultos de comunidades herbáceo-arbustivas (+6%), além das áreas com outras folhosas e eucaliptos (+5%), variaram com menor representatividade, do ponto de vista da disponibilidade de habitat (figura 9).
3.3. Os graus da infestação (análise nível de detalhe)
Para determinação dos graus da infestação das Acácias na Bacia do Rio Arouce, foram avaliados os graus de ocupação e de sociabilidade das espécies A. dealbata e A. melanoxylon, tendo utilizado, para efeito dessa avaliação, uma grelha de referência 200x200m, onde foram coletados dados da presença e abundância dos taxa selecionados atribuindo parâmetros arbitrários quantitativos (1-5) e qualitativos (1-3) relacionados aos graus de ocupação (<5%; 5-25%; 25-50%; 50-75%; >75%) e graus de sociabilidade (indivíduos isolados, pequenas manchas, grandes manchas) das populações invasoras.
O mapa da infestação em escala de detalhe (figura 10) desdobrou três graus diferentes das invasoras: (i) Acácias naturalizadas (mesmo grau das espécies na escala generalista em 1960, com presença de indivíduos isolados); (ii) Acácias invasoras (mesmo grau das espécies na escala generalista em 1980, com pequenas manchas), e (iii) Acácias transformadoras (mesmo grau das espécies na escala generalista em 2010, com grandes manchas).
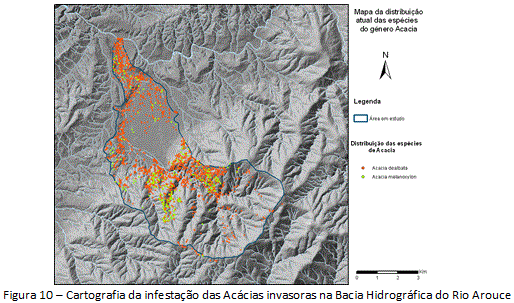
Baseando-se nas considerações acerca da ocupação/sociabilidade, estima-se um gradiente de três contextos distintos da invasão em nível de detalhe na área de estudo: 1) um mais antigo/introdução (unidade intermédia), com os maiores valores de densidade e cobertura total das espécies (nº de espécies >400 por cada área de 200x200m) (etapa de expansão potencial/2010); 2) um mais recente/invasão inconsolidada (unidade cabeceiras), com os menores valores de densidade e cobertura das espécies (nº de espécies <35 por cada área de 200x200m) (etapa de estabelecimento/1960); 3) e um contexto intermediário/invasão em curso (unidade terminal), com valores médios de densidade e cobertura das espécies (nº de espécies <250 e >100 por cada área de 200x200m) (etapa de expansão inicial/1980).Na escala de detalhe, o 1° grau se refere à unidade cabeceiras. Nessa unidade a A. dealbata apresenta-se com indivíduos isolados, com baixo valor de ocupação (valor dominante do grau de ocupação <5%, por cada parcela de 200x200m), enquanto que a A. melanoxylon é menos frequente, com elevado percentual de presença não identificada. O 2° grau se refere à unidade terminal. Nessa unidade a densidade das espécies teve aumento (comparado às cabeceiras), com valor dominante de ocupação entre 5-25%, por parcela de 200x200m. O 3° grau se refere à unidade intermédia, onde a densidade de indivíduos é a mais importante da área de estudo. Nessa unidade foram observados os valores mais elevados para o grau de ocupação da A. dealbata (valor dominante de ocupação >75%, por parcela de 200x200m).
3.4. Organização geossistêmica – espacial (análise integrada I)
A partir da correlação dos dados obtidos durante a pesquisa com o modelo geossistêmico, alguns geofatores são considerados na análise da espacialidade e da temporalidade do fenómeno da invasão por Acácias na Bacia Hidrográfica do Rio Arouce. Carcteriza-se o potencial ecológico e a exploração biológica a partir das três unidades geossistêmicas identificadas por meio de mapeamento (escala 1/25000) (figura 11), com avaliação combinada da distribuição espacial das espécies focais e seus graus de infestação: a unidade terminal, a unidade intermédia e a unidade cabeceiras da referida bacia hidrográfica.
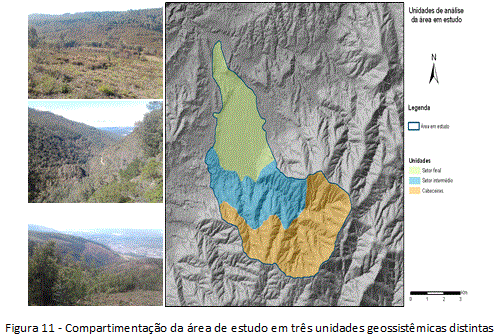
O potencial ecológico da unidade terminal é marcado pela presença dos aluviões do Rio Arouce e as dinâmicas associadas ao balanço hídrico-climatológico para áreas de fundo de vale (foz do Rio Arouce), caracterizadas pelo clima húmido em condições higrófilas (200mm). A evolução geossistêmica dessa unidade ambiental permite sua caracterização em três padrões básicos de identificação: i) área de menor elevação da bacia em níveis topográficos (variante zona litoral); ii) revestimento com areias, cascalhos, argilas e fluvissolos (formação sedimentar); e iii) regadio dominante. A exploração biológica especificamente representada pelas paisagens cuja fitofisionomia é marcada pela presença dos Acaciais, se espacializou no potencial ecológico como resultado de interações ambientais distintas, caracterizando a unidade pela presença de pequenas manchas de Acácias, indicando possível processo de invasão em curso, cuja dispersão, aparentemente, é proveniente da unidade intermédia (unidade mais elevada em termos topográficos).
O potencial ecológico da unidade intermédia divide-a em duas subunidades: 1) fundo de vale (topografia 400m), com areias (fluvissolos) e xistos argilosos, predominantemente agrícola, sob condições higrófilas e/ou mesohigrófilas (200 mm); 2) topo de vale e vertentes (topografia 700m), com xistos, granitos e cambissolos, uso florestal e agrícola, em condições climatófilas (100 mm). A exploração biológica na unidade intermédia é marcada pela presença de grandes manchas de Acácias invasoras, o que indica invasão consolidada, com indivíduos em grau transformador do processo, correspondendo a atual área fonte de propágulos, com dispersão, aparentemente, para o sentido norte (unidade terminal).
O potencial ecológico da unidade cabeceiras, na porção sul da bacia hidrográfica, está caracterizado pelo complexo xisto-grauváquico, área de formações geológicas sedimentares e metamórficas (xistos e granitos). A unidade está localizada na posição topográfica mais elevada da área (900m) e apresenta morfologia ondulada (declives 45 graus), em condições climáticas tipo climatófilas (100 mm) e ocupação do solo predominantemente de incultos de comunidades vegetais arbustivas e abertas. Quanto ao grau de infestação das Acácias, a exploração biológica no potencial ecológico da unidade cabeceiras se define por distribuição pontual de indivíduos isolados, indicando um contexto de invasão inconsolidado. Os indivíduos presentes são jovens, o que aponta para a chegada recente das invasoras, proveninentes provavelmente de algum incêndio ocorrido ou outra atividade antrópica recente. É o setor da área de estudo mais passível de controlo (tipo resposta rápida).
Nesse contexto paisagístico, verificam-se claramente as escalas da análise geossistêmica (Bertrand, 1968) no estudo da espacialização do fenómeno da invasão biológica, com a presença de um mosaico de geofácies definidos pelas diferenças em termos de concentração das espécies, associadas às variações do potencial ecológico entre unidades.
3.5. Organização geossistêmica – temporal (análise integrada II)
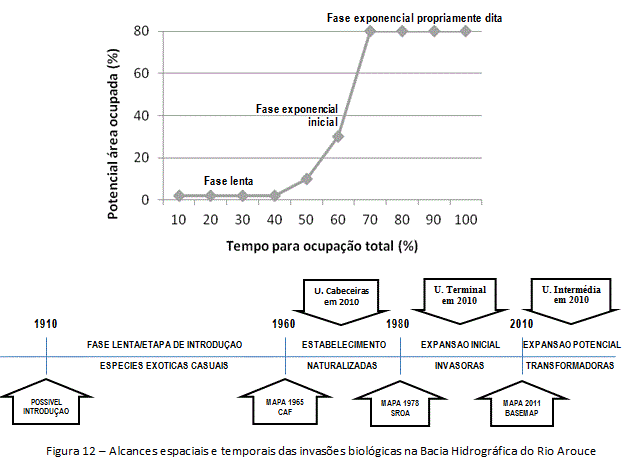
A análise da invasão na Bacia do Rio Arouce à luz dos modelos da invasão e do geossistema levantou a necessidade de uma ilustração da multitemporalidade desde a introdução à consolidação dos Acaciais (figura 12). Por meio dos levantamentos de campo e da pesquisa de gabinete, decodificou-se pixel a pixel e delineou-se a cartografia das espécies focais a fim de apreender os estados das invasões. A inserção do modelo da invasão em uma série evolutiva pode se dar através da proposta de Williamson (1996) e Bertrand (2009). As diferentes propostas configuram um conjunto teórico, simplesmente indicativo (figura 12), que deve ser adaptado em função da problemática levantada na pesquisa.
O estado das variações decenais permite um primeiro balanço cíclico da temporalidade das invasões, além de inserir os impactos ou desastres naturais no ritmo do geossistema. A história da invasão na área de estudo pôde ser delineada, no máximo, a partir de cerca de 50 anos atrás, quando a distribuição das Acácias foi mapeada pela primeira vez através da Carta Agrícola e Florestal de Portugal (CAF/1960). Entre cerca de 1960 e 2010 a Bacia do Rio Arouce apresentou três padrões distintos no âmbito da invasão (figura 12), com acréscimo de cerca de 10% da área invadida, passando de 55 hectares invadidos em 1960 (fase exponencial inicial), para 936 hectares invadidos em 2010 (expansão potencial). O aumento na área invadida esteve mais de perto ligado às transformações nas áreas com florestas de resinosas e culturas agrícolas de regadio e olivais, do que dos incultos e áreas sociais.
A introdução das espécies deverá ter ocorrido há mais de 100 anos. Nos primeiros 50 anos, o processo permaneceu em fase lenta. A partir de 1960, os indivíduos isolados terão começado a aumentar em populações, na transição para a fase exponencial inicial do processo (estabelecimento). A passagem dessa fase para a fase exponencial propriamente dita (expansão inicial) deverá ter ocorrido 20 anos depois, com dispersão para longe do foco da introdução (unidade intermédia). As oscilações nos graus da invasão em 20 anos (1960-1980) aumentaram assustadoramente até 2010 (expansão potencial/sucesso da invasão), estimuladas pelas mudanças na estrutura da paisagem e disponibilidade do habitat.
4. Conclusões
Este estudo mostrou que as mudanças na estrutura da paisagem estimularam a dinâmica espacio-temporal das invasões na Bacia do Rio Arouce. Os resultados das mudanças nos usos do solo entre 1960-2010 mostraram que, de um modo geral, houve uma significativa transformação na estrutura da paisagem da área de estudo, com decréscimo das áreas com culturas agrícolas (culturas de regadio, olivais/pomares) e florestas de resinosas, que perderam seus espaços originais, sobretudo, para áreas com florestas invasoras de Acaciais.
Em análise ao nível de detalhe, nos termos dos graus da infestação atuais das Acácias na Bacia do Rio Arouce, a maior parte dos Acaciais observados na unidade terminal foram reconhecidos como pequenas manchas, enquanto que na unidade intermédia os Acaciais estão particularmente associados a categoria de grandes manchas, e na unidade cabeceiras grande parte das espécies invasoras presentes enquadram-se como indivíduos isolados.
Este resultado indica que as áreas de cabeceiras, com dominância de indivíduos isolados, podem ser caraterizadas, do ponto de vista da invasão biológica, como áreas de chegada recente de espécies invasoras, onde os indivíduos de Acácias ainda não se estabeleceram. Este resultado aponta para a possibilidade de se realizarem com maior êxito atividades de monitorização e controlo nesta unidade, já que entre as unidades geossistêmicas da bacia, é a que apresenta uma ocorrência menos significante das espécies invasoras.
O resultado apontou para a unidade intermédia, onde dominam topografias 400m-700m, uma presença pronunciada de espécies de A. dealbata e A. melanoxylon, o que indica que a chegada dos indivíduos deverá ter ocorrido há bastante tempo, complementando a hipótese de que a introdução inicial deverá ter ocorrido nesta unidade. Para esta unidade, onde o processo da invasão se encontra consolidado, sugerimos que sejam incentivadas atividades de sensibilização, no sentido de alertar para os riscos do aumento das áreas com Acácias, numa tentantiva de mitigar o processo, e a realização de estudos no âmbito da detecção remota das espécies, com vistas a subsidiar atividades de controle/monitorização.
Os resultados dos graus da infestação para as áreas mais baixas topograficamente, apontaram para a presença de pequenas manchas de Acácias. A menor densidade das populações na unidade terminal da Bacia do Rio Arouce comparativamente a unidade intermédia, é indicadora de um processo de invasão em curso dos taxa focais nesta unidade. Nós sugerimos a realização de atividades de sensibilização e educação ambiental, junto aos inúmeros proprietários privados das áreas agrícolas e das residências presentes, para que estes compreendam a gravidade do problema da invasão e possam agir no sentido de não contribuir para com o aumento da infestação e da área invadida por Acácias.
Os dados da presença e abundância das Acácias Australianas na Bacia do Rio Arouce confirmaram o sucesso da invasão. Como conclusão, o fato dos graus da infestação serem mais elevados na unidade terminal da bacia, comparativamente a unidade cabeceiras, e entendendo que foi na unidade intermédia onde ocorreu a introdução inicial, e por isso funciona hoje como fonte de propágulos, isso parece indicar que o avanço da invasão é mais importante no sentido correspondente a unidade terminal, o que está de acordo com o princípio teórico de que a estrutura da paisagem (hidrografia, declividade, ocupação do solo) é importante a considerar no processo da invasão, pois parece determinar o padrão principal em termos da direção das invasões biológicas na Bacia Hidrográfica do Rio Arouce.
5. Referências
BERTRAND, Georges. Paysage et géographie physique globales: esquisse methodologique. Révue de Géographie des Pyrenées et Sud-Ouest. Toulouse, v.39, p.249-72, 1968. [ Links ]
BERTRAND, Claude. BERTRAND, Georges. Une Géographie Traversière. L’environnement à Travers Territoires et Temporalités. Paris : Éditions Arguments, 2002. ISBN 2909109275
BERTRAND, Georges; BRIFFAUD, Serge (dir.). Le paysage: retour d’expériences entre recherche et projet. Bordeaux: Centre Départemental du Patrimoine, 2008.
BERTRAND, Georges. En passant par le paysage... parmi lieux et milieux, environnements et territoires. GÉODOC – Documents de Recherches de l’Institut Daniel-Faucher, nº 56. Toulouse : Université de Toulouse – Le Mirail, 2009.
BERTRAND, Claude; BERTRAND, Georges. Uma geografia transversal e de travessias: o meio ambiente através dos territórios e das temporalidades. PASSOS, Messias Modesto (org.). Maringá: Massoni, 2009. ISBN 9788588905955 [ Links ]
CADOTTE, M. W., MACMAHON, S. M. & FUKAMI, T. Conceptual ecology and invasion biology: reciprocal approaches to nature. Springer, 2006. [ Links ]
CARVALHO, L. M. et al. Disturbance influences the outcome of plant–soil biota interactions in the invasive Acacia longifolia and in native species. Oikos, 119: 1172–1180, 2010.
CHORLEY, R. J. & HAGGETT, P. Modelos físicos e de informação em Geografia. Rio de Janeiro: Livros Técnicos e Científicos S. A. e Ed. da USP, 1974. [ Links ]
CHRISTOFOLETTI, Antonio. Modelagem de sistemas ambientais. São Paulo: Blucher, 1999. ISBN 9788521201779. [ Links ]
CRONK, Q. C. B. & FULLER, J. L. Plant Invaders: the threat to natural ecosystems. Springer, 1995. [ Links ]
DAVIS, M. A. Invasion biology 1958-2005: The pursuit of science and conservation. In: Conceptual ecology and invasion biology: reciprocal approaches to nature, volume 1, chapter 3. 2006.
ESPINOLA, A. L. et. al. Espécies invasoras: conceitos, modelos e atributos. Interciência, v. 32, n. 9, set. 2007. [ Links ]
FERNANDES, M. M. Acacias e geografia histórica: rotas de um percurso global. Cadernos do curso de doutoramento em Geografia. Faculdade de Letras da Universidade do Porto. Março de 2012. [ Links ]
HENDERSON, S. et al. Progress in invasive plants research. Progress in Physical Geography: 30, 1. pp. 25–46. 2006.
IUCN [International Union for the Conservation of Nature]. 1999. IUCN guidelines for the prevention of biodiversity loss due to biological invasion. Species 31/32: 28-42.
LEVINE, J. M. et al. Mechanisms underlying the impact of exotic plant invasions. Phil. Trans. Royal Society, 270: 775-781, 2003. [ Links ]
LOCKWOOD, J. L. et al. Invasion Ecology. Blackwell Publishing. 2007. [ Links ]
MACDONALD, G. Biogeography: Introduction to Space, Time and Life. Willey, 1°edition. 2003. [ Links ]
MARCHANTE, E. et al. Guia prático para a identificação de plantas invasoras de Portugal Continental. Universidade de Coimbra. 2008. [ Links ]
MATEO RODRIGUEZ, José Manuel; SILVA, Edson Vicente; LEAL, Antônio Cezar. Paisaje y geosistema: apuntes para una discusión teórica. Revista Geonorte, Edição Especial, v.1, n.4, 2012. [ Links ]
MCNEELY, J. Invasive species: a costly catastrophe for native biodiversity. Land Use Water Resource, 1: 1-10, 2001. [ Links ]
MONTEIRO, Carlos Augusto de Figueiredo. Os geossistemas como elementos de integração na síntese geográfica e fator de promoção interdisciplinar na compreensão do ambiente. Revista de Ciências Humanas, Florianópolis, v. 14, n. 19, 1996. [ Links ]
NUNES, A.; Figueiredo, A. & Almeida, A. C. Abandono agrícola no interior centro e norte de Portugal: dinâmica da vegetação e impactes hidrogeomorfológicos. VI Congresso da Geografia Portuguesa, Lisboa, 17-20 de Outubro de 2007. [ Links ]
NUNES, A. N. et al. Soil erosion and hydrological response to land abandonment in a central inland area of Portugal. Land Degradation & Development. 21: 260–273. 2010.
OLIVEIRA-COSTA, J. L. P. Os Caminhos da Invasão do Género Acacia Mill. na Bacia do Rio Arouce. Tese de Mestrado. Faculdade de Letras da Universidade de Coimbra. 2014. [ Links ]
PIMENTEL, D. et al. Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecological Economics. 52: 273-288. 2005. [ Links ]
PYŠEK, P., REJMÁNEK, M., PRANCH, K. & WADE, M. Plant Invasions - general aspects and special problems. Netherlands, Academic Publishing Amsterdam, Amsterdam, 713p. 1995.
RICHARDSON, D. M. et al. Humanâmediated introductions of Australian Acacias – a global experiment in biogeography. Diversity and Distribution, 2011.
SHIGESADA, N. & KAWASAKI, K. Biological Invasions – Theory and Practice. Oxford University Express. Oxford, Great Britain: 204p. 1997.
SIMBERLOFF, D. & REJMÁNEK, M. Encyclopedia of Biological Invasions. University of California Press, Berkeley & Los Angeles. 2011. [ Links ]
SOTCHAVA, V. B. O estudo dos geossistemas. Métodos em questão. São Paulo: nº 16, IGEOG – USP, 1977.
SOUZA, Reginaldo. O sistema GTP (Geossistema-Território-Paisagem) aplicado ao estudo sobre as dinâmicas socioambientais em Mirante do Paranapanema-SP. Dissertação (mestrado). Programa de Pós-graduação em Geografia. UNESP: Presidente Prudente, 2010. [ Links ]
SOUZA, Reginaldo. Raia Divisória ou Raia Socioambiental ? Uma (re)definição baseada na análise da paisagem através do sistema GTP. Tese (doutorado). Programa de Pós-graduação em Geografia. UNESP: Presidente Prudente, 2015. [ Links ]
STODDART, D. R. Organismo e ecossistema como modelos geográficos. São Paulo: Livros Técnicos e Científicos S. A. e Ed. da USP, 1974. [ Links ]
TRICART, J. Ecodinâmica. Rio de Janeiro IBGE/SUPREN, 1977. [ Links ]
VAVILOV, N. I. Origin and geography of cultivated plants. Great Britain, Cambridge University express, Cambridge. 497p. 1992. [ Links ]
VILÀ, M. et al. ¿Qué son las invasiones biológicas? Invasiones Biológicas. Consejo Superior de Investigaciones Científicas, CSIC/Madrid. 2008. [ Links ]
VILÀ, M. et al. Ecological impacts of invasive alien plants: a meta-analysis of their effects on species, communities and ecosystems. Ecology Letters, 14: 702-708, 2013. [ Links ]
VITOUSEK, P. M. et al. Introduced species: a significant component of human-caused global change. New Zealand Journal of Ecology, 21: 1-16, 1997. [ Links ]
WATTS, D. Principles of Biogeography: an introduction to the functional mechanisms of ecosystems. McGraw-Hill Publishing Co. Ltd. 1971. [ Links ]
WILLIAMSON, M. Biological Invasions. Chapman & Hall Publishing. London: 244p. 1996. [ Links ]
6. Agradecimentos
Agradecemos a colaboração dos professores Dr. António Campar, Dr. Albano Figueiredo e Dra. Adélia Nunes, pela valiosa orientação nos trabalhos de gabinete (CEGOT/UC) e campo. Ainda, agradecemos a Profa. Dra. Elizabete Marchante, pela assistência no estágio CEF/UC.

{kind=link}
{kind=link}
{kind=link}