









Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Nutrícias
versión On-line ISSN 2182-7230
Nutrícias no.17 Porto jun. 2013
ARTIGO DE REVISÃO
Conjugated Linoleic Acid and Exercise may Share One Biochemical Pathway to Induce Fat-loss
Ácido Linoleico Conjugado e Exercício podem Partilhar Uma Via Bioquímica para Induzir a Perda de Gordura
Hiskias G. Keizer1
1Board certified toxicologist, senior scientist, Stepan Specialty Products
ABSTRACT
Conjugated linoleic acid is a functional ingredient which induces fat-loss in various high quality studies in humans. Although the effect is usually moderate in size, both in vitro studies with conjugated linoleic acid and studies on mice suggest that this effect is real and has a functional basis. Evidence, from in vitro studies and from studies in mice, shows that conjugated linoleic acid affects the expression of a number of genes or gene products which are involved in fat accumulation in adipose tissue. Based on scientific literature we hypothesize that these genes are part of an AMP-kinase dependent body composition regulating (ABC) network that functions both in adipocytes and muscle cells. This network may affect not only adipose tissue but also, lean body mass and exercise endurance and is probably also activated by doing exercise. The consequence of this model is that exposure to conjugated linoleic acid and exercise together may help to enhance fat-loss since they share a common biochemical pathway.
Keywords: Conjugated linoleic acid, Sports nutrition, Exercise, Fat-loss, Biochemical pathway
RESUMO
O ácido linoleico conjugado é um ingrediente funcional, que induz a perda de gordura, indicado em vários estudos em humanos de elevada qualidade. Embora o efeito seja geralmente moderado em dimensão, tanto os estudos in vitro com o ácido linoleico conjugado como os estudos em ratinhos sugerem que este efeito é real e tem uma base funcional. Evidências, de estudos in vitro e de estudos com ratinhos, demonstram que o ácido linoleico conjugado afecta a expressão de um número de genes ou produtos de genes que estão envolvidos na acumulação de gordura no tecido adiposo. Com base na literatura científica sugerimos a hipótese de que esses genes são parte de uma rede de regulação da composição corporal dependente da AMP-quinase, que funciona tanto em adipócitos como em células musculares. Esta rede pode afectar não só o tecido adiposo, mas também, a massa magra e resistência ao exercício e, provavelmente, é também activada pela realização de exercício. A conseqüência desse modelo é que a exposição ao ácido linoleico conjugado e exercício em conjunto podem ajudar a melhorar a perda de gordura, desde que partilhem uma via bioquímica comum.
Palavras- Chave: Ácido linoleico conjugado, Nutrição desportiva, Exercício, Perda de gordura, Via bioquímica
INTRODUCTION
Conjugated linoleic acid (CLA) is known to consistently induce fat loss in humans, but to a limited extent (1,2). However, both in vitro studies with CLA and studies with CLA in mice suggest that this effect is real and has a functional basis. Recently Kennedy et al published a review on the mechanism by which CLA induces fat-loss (3). This work describes that CLA may reduce obesity by at least 5 different mechanisms. However it appears puzzling how a simple fatty acid can have so many different body fat reducing mechanisms of action at the same time. Therefore we looked for a unifying hypothesis. This article describes such a hypothesis and discusses the consequences of this hypothesis if it was correct.
The Adiposity Related Genes Affected by CLA Treatment
CLA affects the expression and or functional activity of a number of genes or gene products (proteins) which are involved the regulation of fat accumulation in adipose tissue. These genes include AMPK (4-7), CPT1 (3, 8-13), Sirt1 (5), PPAR-gamma (13,14), PGC1 (7), and UCP 1/2/3 (11, 15-17).
Of all these genes, AMPK appears to have a central role in the mechanism of fat-loss by CLA as the gene expression observed in cells treated with Metformin, a pharmacological tool to activate AMPK (18), is very similar to the gene expression as induced by exposure of cells to CLA (5) and since AMPK is activated rapidly after exposure to CLA (6).
Effects of Adiposity Genes on Endurance Capacity and Lean Body Mass
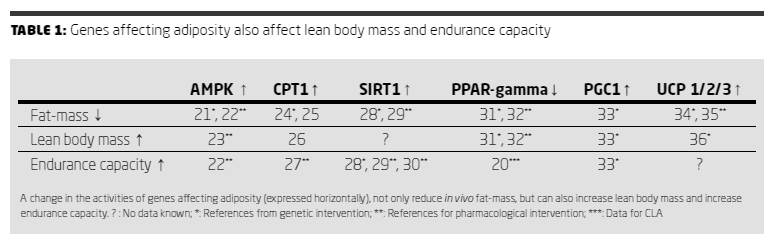
While studying the effects of AMPK, CPT1, SIRT1, PPAR-gamma, PGC1 and UCP1/2/3 on adiposity the attention was drawn to the fact that these genes do not only share effects on adiposity but also on lean body mass and endurance capacity (Table 1). For CLA it has already been shown that it increases lean body mass
in humans (19) and improves endurance capacity in mice (20). However it was not described before that possibly the same genes are involved in improving exercise endurance.
The AMPK-Dependent Body Composition Regulating Network
Since most of the genes mentioned above (AMPK, CPT-1, SIRT-1, PPAR-gamma, PGC-1 and UCP-1/2/3) are involved in regulating metabolism, in fat loss from adipose tissue, in increases in lean body mass and in improving endurance capacity it seems reasonable to suspect that they are all part of one biological network.
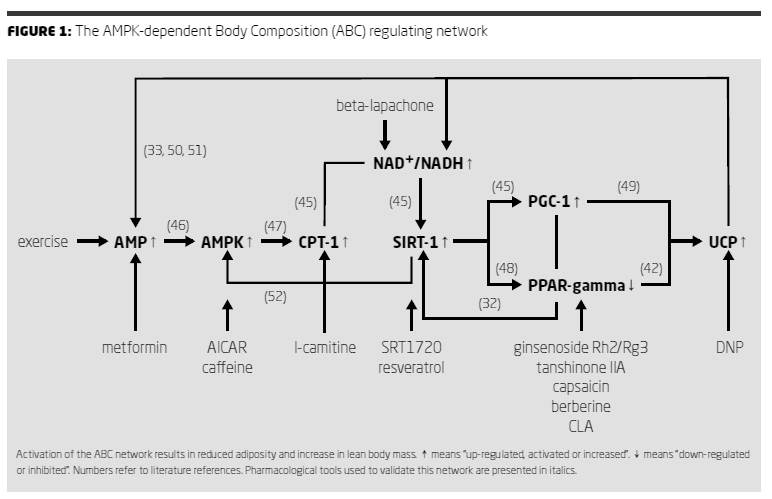
Therefore scientific literature was evaluated with respect to functional inter-relationships between these genes and gene products. This evaluation resulted in the “AMPK-dependent body-composition-regulating network” (ABC-network) of genes as presented in Figure 1. This network is essentially
the same as the energy sensing network as described by Canto (37). For the purpose of this evaluation the potential regulatory role of PPAR gamma was added to Canto’s network. This network appears to work both in adipocytes as in muscle cells as both are relatively rich in PPAR gamma receptors. Exercise probably also activates this network as exercise activates AMPK (by producing AMP from ATP), increases beta-oxidation and down-regulates PPAR gamma (38).
We suspect that CLA mainly affects the AMPK-dependent body composition regulating network by acting as a functional PPAR gamma antagonist (14) since other functional PPAR gamma antagonists like tanshinone IIA (32), ginsenoside Rg3 (39), capsaicin (40), berberine (41) and ginsenoside Rh2 (42) have similar effects on metabolism. Since PPAR gamma is a nuclear receptor involved in gene expression of various genes, inhibition of PPAR gamma signaling by CLA will result in an altered gene expression of various genes. Uncoupling protein-2 (UCP2) is known to be up-regulated by a genetic reduction of PPAR gamma-signaling in muscle (43). Functionally this reduced PPAR gamma signalling is similar to the effect of a functional PPAR gamma receptor antagonist like CLA (14). Uncoupling of the mitochondrial oxidative phosphorylation either by increased expression or activation of uncoupling proteins (44) or by chemical uncoupling by 2,4, dinitrophenol (45) results in increased AMP formation and increased NAD/NADH ratio. These effects are known either directly or indirectly to activate AMPK, CPT1 and SIRT1 (46). This model therefore explains how just one basic mechanism of action of CLA (functional PPAR gamma antagonism) can affect not only body composition but also improve endurance capacity by activating the ABC network.
Interaction of CLA with Exercise, Nutraceuticals and Drugs
As explained above, all genes depicted in the ABC-network of Figure 1 result in
reduced adiposity if their activities are changed in the direction as mentioned in Figure 1. Also exercise (52), treatment with metformin (23), AICAR (22),
carnitine (25), resveratrol (30), SRT1720 (55), tanshinone IIA (32), ginsenoside Rh2 (42), CLA (1), Dinitrophenol (DNP) (35) and beta-Lapachone (56), all depicted in Figure 1, can result in reduced adipogenicity. Of these
treatments at least resveratrol (30), metformin (57), AICAR (22) and CLA (20) have shown to improve endurance. This suggests that all these treatments may enhance the effect of CLA by working on the same network. A potent and early effect of CLA is activation of AMPK. Other activators of AMPK including metformin (57), AICAR (22) and exercise (58, 59) have similar effects on metabolism and endurance capacity. For this reason Narkar positioned AICAR as an “exercise mimetic” (22). The following effects on biology, which are typically caused by exercise, are known to be induced by exposure to CLA: Reduced fat-mass (1), increased lean body mass (19), improved exercise endurance (20, 60), increased strength (60, 61) increased bone mineralisation (62), reduced inflammatory disease (63) and protection against cancer (64). In mice CLA and exercise affect the same biochemical network and share many similar effects on physiology, and therefore CLA may also qualify as an “exercise mimetic”, at least in mice.
Effect of CLA on ABC-Network Activation in Humans
The data as described above are mainly gathered from studies in mice and in vitro systems because studies in humans usually are not very suitable to investigate biochemical networks or mechanisms. However there is evidence to suggest that the ABC-network is also affected by CLA in humans: Effects on adiposity (1) and lean body mass (19) have been confirmed in humans. Mechanistic evidence for activity of the ABC network activated by CLA in humans was provided by Herrmann et al (17): Exposure of humans to a chronic dose of CLA which is sufficient to induce fat-loss resulted in an increase in UCP2 expression as well as a decrease in PPAR gamma expression in adipose tissue. Since these markers in combination with fat loss and increase in lean body mass can be seen as biomarkers for activity of the ABC-network it is tempting to speculate that CLA also activates the ABC-network in humans.
To date two studies were done to measure the effects of CLA on strength. Both these studies (60, 61) showed statistically significant increase in strength. An initial small trial to test the effect of CLA on performance was performed by Colakoglu in 2006 (65). In this trial CLA showed consistent improvement of endurance but the effects were not statistically significant. Statistically significant effects of CLA on endurance have been published by Ha et al in 2010 (60). However more work is needed to quantify this effect.
Although no final proof exists that CLA activates the ABC network in humans, the totality of evidence does suggest that CLA activates the ABC network in humans since evidence suggest that CLA reduces fat-mass (1), increases lean body mass (19) and strength (60, 61) up-regulates UCP2 (17) and down-regulates PPAR gamma (17).
CRITICAL ANALYSIS
A weight-loss claim for CLA has not been approved by EFSA. Also fat-loss as described for CLA in this article is not observed in all human trials (66-69). Therefore when all studies on CLA are included, the effects of CLA on weight-loss or fat-loss are not conclusive. It is therefore important to mention that fat-loss as described in this article is only observed in a selection of studies: These studies were selected as being high quality (conducted according EFSA guidelines) and they were further limited to those using volunteers with a BMI between 25 and 32 (the target population for CLA). Also, only studies were selected which use a dose of ⥠3 grams of mixed isomers of CLA for a period of ⥠12 weeks, to assure adequate dosing.
CLA and exercise appear to share various effects on biochemistry and physiology. However taking CLA is not exactly the same as doing exercise. Exercise activates the ABC pathway by consuming ATP and producing AMP. CLA does so by acting as a functional PPAR receptor antagonist. Exercise is known to increase insulin sensitivity. Since mechanistically CLA has the opposite effect on insulin sensitivity as Rosiglitazone (70) (a PPAR gamma receptor agonist), this may explain why this health benefit is not observed for CLA. Exercise also induces temporally metabolic acidosis, increased body temperature, hypoxia and hormonal changes which may promote health in a way that is not necessarily mimicked by CLA.
Since fat-loss effects of CLA are generally stronger if CLA is taken in combination with exercise and CLA is likely to potentiate the ABC network-related effects of exercise, it is advisable to use CLA in combination with exercise to promote fat-loss, but not instead of exercise.
CONCLUSIONS
The totality of data evaluated in this article, suggest that CLA activates the ABC-network not only in animals but possibly also in humans. This explains several of the physiological effects as observed for CLA. This network is probably also activated by doing exercise.
DISCLAIMER
The author is employed by Stepan Specialty Products B.V., which commercially produces CLA.
REFERENCES
1. Whigham, L. et al. Efficacy of conjugated linoleic acid for reducing fat mass: A meta-analysis in humans. Am. J. Clin. Nutr. 2007: 85: 1203-1211
2. [ Links ] Onakpoya et al, The efficacy of long-term conjugated linoleic acid (CLA) supplementation on body composition in overweight and obese individuals: a systematic review and meta-analysis of randomized clinical trials. Eur. J. Nutr. 2011: 51 (2):127-134
3. [ Links ] Kennedy, A. et al. Antiobesity mechanisms of action of conjugated linoleic acid. J. Nutr. Biochem. 2010: 21: 171-179
4. [ Links ] Jiang, S. et al. Conjugated linoleic acid activates AMP-activated protein kinase and reduces adiposity more effectively when used with metformin in mice. J. Nutr. 2009:139: 2244-2251
5. [ Links ] Jiang, S. T10c12 conjugated linoleic acid causes delipidation in 3T3-L1 adipocytes and mice. 2010: Dissertation, University of Nebraska - Lincoln, Nebraska, Lincoln
6. Jiang, S. et al. Activated AMPK and prostaglandins are involved in the response to conjugated linoleic acid and are sufficient to cause lipid reductions in adipocytes. J. Nutr. Biochem. 2011: 22.7: 656-664
7. [ Links ] Wendel, A.et al. Conjugated linoleic acid induces uncoupling protein 1 in white adipose tissue of ob/ob mice. Lipids 2009: 44: 975-982
8. [ Links ] Martin, J. C. et al. Effects of conjugated linoleic acid isomers on lipid-metabolizing enzymes in male rats. Lipids 2000: 35: 91-98
9. [ Links ] Ferramosca, A. et al. Dietary combination of conjugated linoleic acid (CLA) and pine nut oil prevents CLA-induced fatty liver in mice. J. Agric. Food Chem. 2008: 856: 8148-8158
10. [ Links ] Bouthegourd, J. et al. A CLA mixture prevents body triglyceride accumulation without affecting energy expenditure in Syrian hamsters. Journal of Nutr. 2002: 132: 2682-2689
11. [ Links ] LaRosa, P. C. et al. Trans-10, cis-12 conjugated linoleic acid causes inflammation and delipidation of white adipose tissue in mice: A microarray and histological analysis. Physiol. Genomics 2006: 27: 282-294
12. [ Links ] Park, Y. et al. Effect of conjugated linoleic acid on body composition in mice. Lipids 1997: 32: 853-858
13. [ Links ] Brown, J. et al.. Isomer-specific regulation of metabolism and PPAR-gamma signaling by CLA in human pre-adipocytes. J. Lipid Res. 2003: 44: 1287-1300
14. [ Links ] Kennedy, A. et al. Trans-10, cis-12 conjugated linoleic acid antagonizes ligand-dependent PPAR gamma activity in primary cultures of human adipocytes. J. Nutr. 2008:138: 455-461
15. [ Links ] Kang, K, et al. Evidence that the anti-obesity effect of conjugated linoleic acid is independent of effects on stearoyl-coa desaturase1 expression and enzyme activity. Biochem. Biophys. Res. Comm. 2004: 315: 532-537
16. [ Links ] Ealey, K. N. et al. Effects of dietary conjugated linoleic acid on the expression of uncoupling proteins in mice and rats. Lipids 2002: 37: 853-861
17. [ Links ] Herrmann, J.et al. Isomer-specific effects of CLA on gene expression in human adipose tissue depending on PPARgamma2 P12A polymorphism: a double blind, randomized, controlled cross-over study. Lipids Health Disease, 2009: 8: 35, [ Links ] 2009
18. Zhou, G. et al. Role of AMP-activated protein kinase in mechanism of metformin action. J. Clin. Invest. 2001: 108: 1167-1174
19. [ Links ] Schoeller, D. A.et al. A meta-analysis of the effects of conjugated linoleic acid on fat-free mass in humans. Appl. Physiol. Nutr. Metab. 2009: 34: 975-978
20. [ Links ] Mizunoya, W. et al. Dietary conjugated linoleic acid increases endurance capacity and fat oxidation in mice during exercise. Lipids 2005: 40: 265-271
21. [ Links ] Yang, J., et al. Chronic activation of AMP-activated protein kinase-alpha1 in liver leads to decreased adiposity in mice. Biochem. Biophys. Res. Commun. 2008: 370: 248-253
22. [ Links ] Narkar, V. et al. AMPK and PPAR delta agonists are exercise mimetics. Cell 2008: 134: 405-415
23. [ Links ] Ibanez, L. et al. Anovulation in eumenorrheic, nonobese adolescent girls born small for gestational age: Insulin sensitization induces ovulation, increases lean body mass, and reduces abdominal fat excess, dyslipidemia, and subclinical hyperandrogenism. J. Clin. Endocrinol. Metab. 2002: 87: 5702-5705
24. [ Links ] Abu-Elheiga, L. et al. Continuous fatty acid oxidation and reduced fat storage in mice lacking acetyl-coa carboxylase 2. Science 2001: 291: 2613-2616
25. [ Links ] Owen, K. Q. et al. Effect of dietary l-carnitine on growth performance and body composition in nursery and growing-finishing pigs. J. Anim. Sci. 2001: 79: 1509-1515
26. [ Links ] Blanchard, G. et al. Dietary l-carnitine supplementation in obese cats alters carnitine metabolism and decreases ketosis during fasting and induced hepatic lipidosis. J. Nutr. 2002:132: 204-210
27. [ Links ] Vecchiet, L. et al. Influence of l-carnitine administration on maximal physical exercise. Eur J Appl Physiol Occup Physiol 1990: 61: 486-490
28. [ Links ] Bordone, L. et al. Sirt1 transgenic mice show phenotypes resembling calorie restriction. Aging Cell 2007: 6: 759-767
29. [ Links ] Feige, J. N. et al. The endocrine disruptor monoethyl-hexyl-phthalate is a selective peroxisome proliferator-activated receptor gamma modulator that promotes adipogenesis. J. Biol. Chem. 2007: 282 : 19152-19166
30. [ Links ] Lagouge, M. et al. Resveratrol improves mitochondrial function and protects against metabolic disease by activating sirt1 and pgc-1alpha. Cell 2006:127: 1109-1122
31. [ Links ] Tsai, Y. S. et al. Decreased PPAR gamma expression compromises perigonadal-specific fat deposition and insulin sensitivity. Mol. Endocrinol 2009: 23: 1787-1798
32. [ Links ] Gong, Z. et al. The role of tanshinone IIa in the treatment of obesity through peroxisome proliferator-activated receptor gamma antagonism. Endocrinology 2009: 150: 104-113
33. [ Links ] Wenz, T. et al. Increased muscle PGC-1alpha expression protects from sarcopenia and metabolic disease during aging. PNAS USA 2009:106: 20405-20410
34. [ Links ] Gates, A. C. et al. Respiratory uncoupling in skeletal muscle delays death and diminishes age-related disease. Cell Metab. 2007: 6: 497-505
35. [ Links ] Caldeira da Silva, C et al. Mild mitochondrial uncoupling in mice affects energy metabolism, redox balance and longevity. Aging Cell . 2008: 7: 552-560
36. [ Links ] Fuller, P. M. et al. Effects of 2-g exposure on temperature regulation, circadian rhythms, and adiposity in ucp2/3 transgenic mice. J.Appl. Physiol. 2000: 89: 1491-1498
37. [ Links ] Canto, C et al. PGC1, SIRT1 and AMPK, an energy sensing network that controls energy expenditure. Current Opinion. Lipidology 2009: 20: 98-105
38. [ Links ] Tunstall, R. J. et al. Exercise training increases lipid metabolism gene expression in human skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2002: 283: E66-72
39. [ Links ] Hwang, J-T et al. Antiobesity effect of ginsenoside Rg3 involves the AMPK and PPAR gamma signal pathways. Phytotherapy research 2008: 23:262-266
40. [ Links ] Hiromi, Y et al. PPAR gamma partial agonist activity and PPAR alpha inverse agonist activity of capsaicin. J. Pharmacol. Sci. 2006:100 suppl. 1: 236P
41. [ Links ] Huang, C et al, Berberine inhibits 3T3-L1 adipocyte differentiation through the PPAR gamma pathway. Biochem. Biophys. Res. Comm. 2006: 348 (2): 571-578
42. [ Links ] Hwang, J-T et al, Anti-obesity effects of Ginsenoside Rh2 are associated with the activation of AMPK signaling pathway in 3T3-L1 adipocyte. Biochem. Biophys. Res. Comm. 2007: 364: 1002-1008
43. [ Links ] Yamauchi, T. et al. The mechanisms by which both heterozygous peroxisome proliferator-activated receptor gamma (PPAR gamma) deficiency and PPAR gamma agonist improve insulin resistance. J. Biol. Chem. 2001: 276: 41245-41254
44. [ Links ] Fleury, C. et al. Uncoupling protein-2: A novel gene linked to obesity and hyperinsulinemia. Nat. Genet. 1997: 15: 269-272
45. [ Links ] Weinbach, E. C et al.The interaction of uncoupling phenols with mitochondria and with mitochondrial protein. J. Biol. Chem. 1965: 240: 1811-1819
46. [ Links ] Canto, C. et al. AMPK regulates energy expenditure by modulating nad+ metabolism and sirt1 activity. Nature 2009: 458: 1056-1060
47. [ Links ] Carling, D. et al. Purification and characterization of the AMP-activated protein kinase. Copurification of acetyl-coa carboxylase kinase and 3-hydroxy-3-methylglutaryl-coa reductase kinase activities. Eur. J. Biochem. 1989:186: 129-136
48. [ Links ] Park, H. et al. Coordinate regulation of malonyl-coa decarboxylase, sn-glycerol-3-phosphate acyltransferase, and acetyl-coa carboxylase by AMP-activated protein kinase in rat tissues in response to exercise. J. Biol. Chem. 2002: 277: 32571-32577
49. [ Links ] Picard, F. et al. SIRT1 promotes fat mobilization in white adipocytes by repressing ppar-gamma. Nature 2004: 429: 771-776
50. [ Links ] St-Pierre, J. et al. Bioenergetic analysis of peroxisome proliferator-activated receptor gamma coactivators 1alpha and 1beta (PGC-1alpha and PGC-1beta) in muscle cells. J. Biol. Chem. 2003: 278: 26597-26603
51. [ Links ] Zhou, M. et al. Rosiglitazone promotes fatty acyl CoA accumulation and excessive glycogen storage in livers of mice without adiponectin. J. Hepatol. 2010: 53: 1108-1116
52. [ Links ] Schrauwen, P. et al. Improved glucose homeostasis in mice overexpressing human UCP3: A role for AMP-kinase Int. J. Obes. Relat. Metab. Disord. 2004: 28: 824-828
53. [ Links ] Hou, X et al. SIRT regulates hepatocyte lipid metabolism through activating of AMP activated protein kinase. J.Biol Chem. 2008: 283.29: 20015-20026
54. [ Links ] Lee, S. et al. Exercise without weight loss is an effective strategy for obesity reduction in obese individuals with and without type 2 diabetes. J. Appl. Physiol. 2005: 99: 1220-1225
55. [ Links ] Feige, J. N. et al. Specific SIRT1 activation mimics low energy levels and protects against diet-induced metabolic disorders by enhancing fat oxidation. Cell. Metab. 2008: 8: 347-358
56. [ Links ] Hwang, JH et al. Pharmacological Stimulation of NADH Oxidation Ameliorates Obesity and Related Phenotypes in Mice. Diabetes 2009: 58 (4): 965-974
57. [ Links ] Benedini, S et al. Improved resistance to exercise in mice treated with metformin. Acta Physiologica 2010: 200 (suppl 681): 98-98
58. [ Links ] Matsakas A. Endurance exercise mimetics in skeletal muscle. Curr. Sports Med. Rep. 2010: 9 (4): 227-232
59. [ Links ] Richter, E. A. et al. AMPK and the biochemistry of exercise: Implications for human health and disease. Biochem. J. 2009: 418: 261-275
60. [ Links ] Ha, Y. et al. Effects of conjugated linoleic acid on body fat reduction and physical exercise enhancement of obese male middle school students. Jrn. Life Science 2010: 20:1844-1850
61. [ Links ] Pinkoski, C. et al., The effect of conjugated linoleic acid supplementation during resistance training. Med. Sci. Sports Exerc. 2006: 38(2):339-348
62. [ Links ] Banu J. et al. Effects of conjugated linoleic acid and exercise on bone mass inyoung male Balb/C mice. Lipids in Health and Disease 2006: 5:7
63. [ Links ] Huebner S.M. et al.Individual Isomers of Conjugated Linoleic Acid Reduce Inflammation Associated with Established Collagen-Induced Arthritis in DBA/1 Mice. J. Nutr. 2010: 140: 8 1454-1461
64. [ Links ] Pariza, M. et al. Conjugated linoleic acid and the control of cancer and obesity. Toxicol. Sci. 1999: 52 (suppl 1): 107-110
65. [ Links ] Colakoglu, S.et al. Cumulative effects of conjugated linoleic acid and exercise on endurance development, body composition, serum leptin and insulin levels. J. Sports Med. Phys. Fitness 2006: 46: 570-577
66. [ Links ] Joseph S.V. et al. Conjugated linoleic acid supplementation for 8 weeks does not affect body composition, lipid profile, or safety biomarkers in overweight, hyperlipidemic men. J.Nutr. 2011: 141(7):1286-1291
67. [ Links ] Sluijs I. et al. Dietary supplementation with cis-9,, trans-11 conjugated linoleic acid and aortic stiffness in overweight and obese adults. Am.J.Clin. Nutr. 2010: 91(1): 175-183
68. [ Links ] Salas-Salvado J. et al. Conjugated linoleic acid intake in humans: a systematic review focusing on its effect on body composition, glucose, and lipid metabolism. Crit.Rev.Food Sci.Nutr. 2006: 46(6): 479-488
69. [ Links ] Wang Y. et al. Dietary conjugated linoleic acid and body composition. Am.J.Clin. Nutr. 2004: 79(6 Suppl.): 1153S-1158S
70. [ Links ] Mayerson et al. The Effects of Rosiglitazone on Insulin Sensitivity,Lipolysis, and Hepatic and Skeletal Muscle TriglycerideContent in Patients With Type 2 Diabetes. Diabetes 2002: 51: 797-802 [ Links ]
Correspondence
Hiskias G.Keizer
Museumlaan 16, LP Koog aan de Zaan, The Netherlands
hkeizer@stepan.com
Received at 23rd April 2013
Accepted at 05th July 2013

{kind=link}
{kind=link}