Carbon and nutrient amounts in aboveground biomass, understory and soil in a pine stand chronosequence
Manuel Madeira1, António Fabião2, Fernando Páscoa2, Maria do Carmo Magalhães2, Maria Cristina Cameira2, Carlos Ribeiro2,3
RESUMO
Avaliou-se a biomassa e a quantidade de nutrientes da parte aérea de povoamentos de pinhal bravo da Mata Nacional de Leiria, com idades abrangendo o período da respectiva revolução, bem como da vegetação sob coberto. A massa e nutrientes das camadas orgânicas e a quantidade de nutrientes nas camadas minerais do solo, até um metro de profundidade, foram também avaliadas. Observou-se um acréscimo da biomassa da parte aérea e da quantidade de nutrientes ao longo da revolução, com os máximos no final da mesma. Nas camadas minerais do solo, observou-se semelhante acréscimo da quantidade de C orgânico e de nutrientes com o avançar da revolução, sobretudo nas camadas superficiais do solo (0-20 cm). A quantidade de C e nutrientes na vegetação sob coberto e nas camadas orgânicas representou uma fracção muito inferior à acumulada nas componentes anteriores. O C orgânico encontrase distribuído em proporção semelhante pelo solo e biomassa aérea, enquanto o N e o Ca estão sobretudo acumulados no solo. O P apresenta-se em elevada proporção nas camadas orgânicas e nos resíduos de abate; o Mg e o K apresentam situação intermédia. Os resultados sugerem que as perturbações decorrentes da exploração dos povoamentos (abate, exploração, remoção de biomassa e modificações de microclima) provocam fortes variações na quantidade de C orgânico e nutrientes no sistema. A gestão apropriada dos resíduos de abate e das camadas orgânicas é crucial para minimizar a perda de matéria orgânica e de nutrientes e assegurar a sustentabilidade do sistema.
Palavras-chave: pinheiro bravo, biomassa, carbono, nutrientes, Mata de Leiria.
ABSTRACT
]]> Biomass and nutrient amounts accumulated in the above ground biomass along the rotation period of maritime pine stands of the Leiria National Forest were evaluated. Amounts of biomass and nutrients of understory were also assessed. Furthermore, mass and nutrients in organic soil layers, as well as nutrient amounts in mineral soil layers (up to 100 cm depth) were evaluated. A strong increment of above ground biomass and accumulated nutrients was observed along the rotation period, with maximum at the end. A similar increment of organic C and nutrients was also observed in the mineral soil layers in the later rotation stage, being mostly allocated to the top soil layers (0-20 cm). Organic C was equally distributed by the above ground biomass and mineral soil, while N and Ca were mostly allocated to the soil. Phosphorus was largely allocated to the organic soil layers and harvest residues, and Mg and K showed an intermediate pattern. Results suggest that system disturbances associated with stand exploitation (harvesting, log removal, harvest residues removal, microclimate changes) may provoke strong variations on the organic C stock and nutrient availability. Proper site management to avoid organic matter and nutrient losses are crucial to assure system sustainability.Key-words: maritime pine, biomass, carbon, nutrients, Leiria National Forest.
INTRODUÇÃO
Os povoamentos de pinheiro bravo (Pinus pinaster Ait.) ocupam em Portugal uma área de cerca de 710 000 ha (área de povoamentos puros, mistos dominantes e jovens actualizada a 2005/2006 pelo Inventário Flores-tal Nacional, disponível em http://www.afn.min-agricultura.pt/portal/ifn/ifn-resultadospreliminares-julho-2006), distribuindo-se sobretudo pelas regiões NUT II Centro (cerca de 400 000 ha) e Norte (192 000 ha). A sua ocorrência no litoral do País e, em especial, nos sistemas dunares, reveste-se de grande significado ecológico, não só pelas funções de estabilização destes sistemas, mas também por poderem considerar-se como uma forma de restauro de um coberto presente naturalmente nestas áreas durante e após a última glaciação (Aguiar et al., 2007). Nas dunas terciárias e paleodunas podem mesmo constituir habitats classificados no âmbito do Plano Sectorial da Rede Natura 2000 (designadamente os Habitats 2180 Dunas arborizadas das regiões atlântica, continental e boreal e 2270 Dunas com florestas de Pinus pinea ou Pinus pinaster subsp. atlantica; cf. Plano Sectorial da Rede Natura 2000 Habitats Naturais, em http://www.icn.pt/psrn2000/caract_habitat.htm#habitats_2).
Nas regiões NUT III do Baixo Vouga, Baixo Mondego e Pinhal Litoral, que abrangem em larga medida a faixa arenosa ocupada por esta espécie na região Centro, os pinhais de pinheiro bravo ocupam uma área de cerca de 130 000 ha, localizando-se no litoral cerca de 48 000 ha de Matas Nacionais, às quais corresponde uma área florestada, maioritariamente por pinheiro bravo, de aproximadamente 30 000 ha (Rego, 2001). Apesar da importância deste sistema no panorama florestal português, ao contrário do observado para outras espécies (e.g. Eucalyptus globulus), são reduzidos e/ou de âmbito limitado os estudos relativos aos povoamentos de pinheiro bravo no que respeita à acumulação e distribuição de nutrientes da biomassa e à acumulação de carbono e nutrientes no solo.
As plantações florestais exploradas intensivamente determinam a acumulação de grandes quantidades de nutrientes na biomassa aérea e nas camadas orgânicas durante a respectiva rotação, o que pode acarretar a transferência de elevadas quantidades de nutrientes do solo (Fisher & Binkley, 2000; Spangenberg et al., 1996) e afectar as características do mesmo (Madeira et al., 2007). As plantações de eucalipto, as melhor estudadas em Portugal, são disso um bom exemplo (Madeira et al., 2007), dada a exploração intensiva em regime de talhadia (Jones et al., 1999; Shammas et al., 2003) e a elevada quantidade de nutrientes alocada nos respectivos resíduos de abate e camadas orgânicas (Madeira et al., 1995; Jones et al., 1999). Este padrão depende naturalmente das espécies florestais e da intensidade e regime da respectiva exploração, incluindo a duração da revolução e as perturbações inerentes à instalação e exploração. Porém, não se conhecem os padrões referentes ao pinheiro bravo, que tem crescimento muito mais lento e é explorado em alto fuste e com revoluções muito mais longas (Correia & Oliveira, 2003), localizando-se as respectivas plantações em solos frequentemente de textura muito grosseira e com baixa disponibilidade em nutrientes; além disso, a massa das respectivas camadas orgânicas, devido à lentidão da decomposição da folhada (Ribeiro et al. 2007), pode ser muito elevada e acumular grandes quantidades de nutrientes.
Para fundamentar a gestão das áreas de povoamentos florestais e dos nutrientes acumulados na respectiva biomassa e resíduos, com vista ao reforço da produtividade e garantia da respectiva sustentabilidade, é crucial conhecer as quantidades de nutrientes nas componentes do ecossistema (Smethurst & Nambiar, 1990; Proe & Dutch, 1994; Spangenberg et al., 1996; Nzila et al., 2002). Além disso, a remoção ou manutenção dos resíduos de abate é objecto de polémica devido não só à relação entre a gestão dos resíduos e a qualidade do solo e a sustentabilidade, mas também à necessidade urgente de novas formas de energia que possam substituir os combustíveis fósseis (Cowie et al., 2006; Stupak et al., 2007). Urge ainda conhecer a dinâmica da acumulação de carbono e a sua distribuição nas componentes dos sistemas florestais para avaliar a eficiência e evolução do respectivo sequestro (Lal et al., 1998).
Neste contexto, desenvolveu-se um estudo para avaliar a quantidade e distribuição do C orgânico e de nutrientes nas componentes do ecossistema pinhal bravo, considerando uma cronossequência abrangendo todo o período da respectiva revolução. Para o efeito, escolheu-se para área de estudo o Pinhal da Mata Nacional de Leiria, devido à especificidade dos respectivos solos (arenosos e considerados pobres em nutrientes) e ao período da revolução aí seguido ser muito longo (cerca de 80 anos; Suzana Gomes, com. pessoal). Foi também objectivo deste estudo avaliar, por intermédio do conhecimento da distribuição mencionada, o efeito da remoção do tronco comercial e dos resíduos de abate, no final da revolução, no stock de nutrientes no solo.
]]> METODOLOGIA
Área de estudo
O estudo decorreu na Mata Nacional de Leiria, mais propriamente em parcelas dos talhões 329, 300, 164 e 279, cujos povoamentos apresentavam 14, 25, 42 e 83 anos de idade, respectivamente. A área onde actualmente se situam estes talhões já se encontrava arborizada em 1841 (Silva & Batalha, 1859; Pinto, 1939).
A precipitação média anual varia entre 710 e 909 mm, respectivamente em S. Pedro de Muel e Marinha Grande (INMG, 1991); a precipitação máxima mensal ocorre no mês de Dezembro (113 e 133 mm, respectivamente), ao passo que o período menos pluvioso tem lugar durante os meses de Julho a Setembro, com precipitações mensais que variam entre 7,7 e 35,3 mm. A temperatura média anual do ar é de 14,1-14,2 ºC, sendo a temperatura média mensal mais elevada nos meses de Julho e Agosto (17,8 e 19,2 ºC, respectivamente em S. Pedro de Muel e Marinha Grande) e a mais baixa no mês de Janeiro (9,9 ºC). A humidade relativa do ar (às 18 horas) é elevada e também decresce de S. Pedro de Muel (84%) para a Marinha Grande (77%).
As formações geológicas da área da Mata Nacional de Leiria são maioritariamente modernas e correspondem essencialmente a dunas e areias de dunas (Morais, 1936; Zbyszewski & Torre de Assunção, 1965). O relevo varia desde aplanado a ondulado, em função do movimento e evolução das dunas. Os solos da Mata Nacional de Leiria foram objecto de vários estudos, sobretudo de natureza morfológica, tendo sido generalizadamente classificados como Podzóis ou Solos Podzolizados (Azevedo, 1944; Cabrita, 1956; Oliveira, 1967; Cardoso et al. 1973). Porém, estudos mais recentes revelam que na realidade são os Arenossolos (sensu WRB, 2006) os solos que aí predominam (Madeira et al., 2007).
Do ponto de vista biogeográfico a Mata de Leiria insere-se no Sector Divisório Português e no Distrito Costeiro Português (Costa et al., 2002), sendo as espécies Erica cinerea, Halimium alyssoides, Halimium ocymoides e Pseudarrhenatherum longifolium consideradas como diferenciais da unidade referida. A vegetação sob coberto na área da Mata de Leiria está representada por inúmeras espécies herbáceas e arbustivas: Arbutus unedo, Erica umbellata, Caluna vulgaris, Phyllirea angustifolia, Smilax áspera, Ulex australis subsp. velwitchianus e Stauracanthos genistoides são espécies características dos pinhais litorais (cf. bioindicadores dos Habitats respectivos, em http://www.icn.pt/psrn2000/caract_habitat.htm#habitats_2), tendo-se também identificado nos locais estudados as espécies Andryala integrifolia, Funaria hygrometrica, Asphodelus lusitanicus, Holcus mollis, Briza maxima, Lithodora prostrata, Cistus salvifolius, Cladonia sp., Rubia peregrina, Crepis capillaris, Simethis planifolia, Cytisus grandiflorus, Daphne gnidium e Erica arborea.
Amostragens
Biomassa aérea da vegetação arbórea
As determinações de biomassa foram efectuadas em parcelas com 40x40 m2de povoamentos de pinheiro bravo da Mata Nacional de Leiria, da classe de qualidade de 20 m (povoamentos com altura dominante de 20 metros aos 50 anos). As parcelas foram sempre delimitadas em áreas aplanadas para garantir a comparabilidade dos resultados. As determinações foram efectuadas antes do desbaste, nas idades em que o modelo de silvicultura adoptado o recomenda. Assim, as amostragens efectuaram-se imediatamente antes dos desbastes aos 14, 25 e 42 anos, e durante o corte final, aos 83 anos. Estimou-se a biomassa no momento do inventário (antes do desbaste), a biomassa exportada pelo des-baste e a biomassa restante após o desbaste. Determinou-se a concentração de nutrientes no lenho, casca, ramos (do último ano, do penúltimo ano e restante parte dos ramos), agulhas (do último ano, do penúltimo ano e restantes agulhas) e frutos (do último ano, do penúltimo ano e restantes frutos)
]]> Para avaliação da biomassa seleccionaramse as árvores de acordo com as classes de DAP e de altura total previamente definidas, utilizando-se uma suta e um hipsómetro de grande sensibilidade. Foi abatida pelo menos uma árvore por classe de diâmetro, obtendose a respectiva biomassa por amostragem directa. As árvores seleccionadas (identificadas por um anel de tinta fluorescente a 1,30m de altura e numeradas) eram isentas de malformações de fuste e copa e estavam em bom estado fitossanitário. Após o abate determinaram-se os seguintes parâmetros: alturas, medidas com fita métrica, total da árvore, do início da copa (primeiro ramo vivo), do início da bicada (aos 7 cm de diâmetro), do cepo e da face inferior de corte das amostras do tronco (rodelas), diâmetro do cepo (todas as medições de diâmetro foram efectuadas sobre casca e sempre nos sentidos Norte-Sul e Este-Oeste, sendo mais tarde efectuada a sua média quadrática), sobre casca e sobre pau (por diferença para a espessura da casca, determinada em dois pontos com medidor de espessura de casca), diâmetro do tronco a 1,30 m, sobre casca e sobre pau, diâmetro do tronco de 2 em 2 m de altura, a partir de 1,30 m até aos 7 cm (bicada), sobre casca e sobre pau (1,30 m, 3,30 m, 5,30 m, etc...), número de verticílos (andares de ramos) e número de ramos por verticílo.Colheram-se (e pesaram-se, se necessário) amostras das componentes da biomassa da árvore ao longo do tronco da mesma e, em cada verticílo, foi seleccionado o ramo que apresentava maior verticalidade (o mesmo método foi usado para a bicada). Nesse ramo separaram-se as seguintes componentes: crescimentos do presente ano, crescimentos do ano anterior e restante material. A primeira fracção colhida foi a correspondente ao crescimento do ano em que se efectuou a amostragem. Os materiais foram seccionados com tesoura de poda e embalados em sa-cos de plásticos. Precedeu-se de igual modo para o crescimento do ano anterior e para o restante material. Esta última fracção, devido à sua elevada quantidade, foi pesada no local, colhendo-se uma sub-amostra de 10 kg, que foi acondicionada em sacos de plástico. Procedimento semelhante foi seguido para a bicada. A colheita de amostras do tronco da árvore (vulgo rodelas) efectuou-se em três pontos ao longo do tronco: na base (ou no cepo), a meio da árvore e imediatamente acima do último toro com aproveitamento. As rodelas tinham uma espessura mínima de 3 cm em todo o seu perímetro e foram acondicionadas em sacos de plástico. No laboratório obtiveram-se os pesos secos a 85 ºC de todos os componentes da biomassa arbórea.
Biomassa da vegetação sob coberto
A avaliação da biomassa da vegetação espontânea (matos) foi efectuada em 15 locais (1x1m2), distribuídos aleatoriamente em cada uma das parcelas de estudo (as mesmas utilizadas para o estudo da biomassa arbórea). A vegetação herbácea e arbustiva localizada dentro do quadrado de 1x1 m2 foi cortada junto à superfície do solo e colocada em saco de plástico. No laboratório pesou-se o conteúdo do saco e retirou-se uma amostra de 200 g para obtenção do peso seco a 85ºC. Após secagem, o material foi moído num moinho centrífugo, reduzindo-se a dimensões inferiores a 0,5 mm.
Camadas orgânicas
A massa das camadas orgânicas de cada uma das parcelas objecto de estudo foi avaliada a partir de amostragens em 15 locais (1x1 m2) aleatoriamente distribuídos, igualmente nas mesmas áreas em que se amostrou a biomassa da vegetação espontânea. O material de cada local de amostragem foi seco a 85 ºC até peso constante. Do material seco foi retirada uma subamostra representativa e que foi moída num moinho centrífugo, sendo reduzida a partículas <0,5 mm.
Camadas minerais do solo
]]> As camadas minerais do solo de cada parcela de estudo consideradas foram amostradas também em 15 locais (os mesmos em que se amostrou as camadas orgânicas), até à profundidade de 1 m. As amostragens incluíram amostras não disturbadas (para determinação da massa volúmica aparente) e disturbadas (para determinação de características químicas do solo). As amostragens incidiram sobre as profundidades de 0-10, 10-20, 20-40, 40-60, 60-80 e 80-100 cm. Esta amostragem não seguiu naturalmente a diferenciação do perfil do solo, dado que foi efectuada com fins meramente quantitativos.As amostras disturbadas foram secas a 45ºC e depois crivadas num crivo com malha de 2mm.
Metodologia analítica
O teor de cinza nos materiais orgânicos foi determinado pela incineração de 1 g de amostra a 450 ºC, durante 6 horas. O teor de N total foi obtido pelo método de Kjeldahl. Considerou-se que o teor de C orgânico desses materiais era de 50%. O P, Ca, Mg e K foram extraídos a partir da cinza por digestão nitro-perclórica. A determinação do P efectuou-se por colorimetria (Watanabe & Olsen, 1965), tendo os demais elementos sido determinados por espectrofotometria de absorção atómica (EAA).
As metodologias utilizadas na caracterização das camadas minerais dos solos são as descritas em Póvoas & Barral (1992). O pH foi determinado numa suspensão de solo em H2O e em KCl 1M (razão 1:2,5) por intermédio de um potenciómetro Metrohm 605. O C orgânico foi determinado por via húmida pelo método descrito por De Leenheer & Van Hoven (1958). O teor de N total foi obtido pelo método de Kjeldahl. As bases de troca foram extraídas por uma solução de acetato de amónio 1M, ajustada a pH 7,0, e determinadas por EAA. O P e o K extraíveis foram obtidos pelo método de Egnér-Riehm, sendo o P determinado por colorimetria e o K por EAA.
Cálculos e métodos estatísticos
O cálculo da biomassa arbórea (e respectivas componentes) por hectare foi extrapolado a partir das árvores medidas e da distribuição do povoamento por classes de diâmetro. A biomassa de cada árvore de cada classe de diâmetro foi extrapolada para as árvores da mesma classe. A biomassa total por hectare foi, assim, obtida pelo somatório da biomassa das árvores pertencentes a cada classe de diâmetro. Quantificou-se também a quantidade de nutrientes por unidade de área (ha) a partir da massa das componentes do ecossistema e da respectiva concentração de nutrientes. Foram estabelecidas correlações entre as concentrações de carbono e nutrientes do solo e a idade dos povoamentos.
]]> RESULTADOS E DISCUSSÃO
Biomassa e mineralomassa arbórea da parte aérea
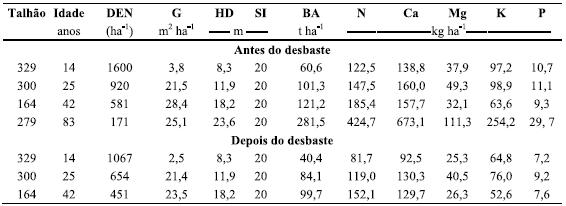
O número de árvores apresentou um forte decréscimo com o avanço da idade dos povoamentos, mas a biomassa da parte aérea aumentou sucessivamente com a idade dos mesmos (Quadro 1). No termo da revolução (abate), aos 83 anos, não obstante o reduzido número de árvores e a ligeira redução da área basal, a biomassa aérea total era mais do dobro da estimada no povoamento com 42 anos de idade (Quadro 1).
Quadro 1 Idade, densidade (DEN), índice de qualidade (SI), altura dominante (HD), área basal (G), biomassa aérea total (BA) (peso seco a 85ºC) e nutrientes acumulados na biomassa dos povoamentos de pinheiro bravo dos talhões 329, 300, 164 e 279, imediatamente antes do desbaste (talhões 329, 300 e 164) e do corte final (talhão 279).
No final da revolução, cerca de 78 % da biomassa da parte aérea correspondia ao lenho (tronco comercial sem casca), enquanto aos ramos correspondia uma percentagem de 11 % e as agulhas correspondiam apenas a 2,8% da biomassa total (Quadro 3). A biomassa arbórea aérea (assim como a quantidade de carbono) no termo da revolução era da ordem de grandeza da estimada para a primeira rotação de plantações de eucalipto, com idades compreendidas entre os 4 e os 20 anos (Fabião, 1986; Madeira, 2007).
A diferença observada para a biomassa, ao longo da idade dos povoamentos, também foi constatada para os nutrientes considerados. Com efeito, a quantidade de N (424,8 kg ha-1) e de Ca (673,1 kg ha-1) era mais do dobro no final da revolução, comparativamente à determinada para um período de 42 anos. As diferenças observadas para o Mg, K e P foram ainda mais acentuadas (Quadro 1). A quantidade de nutrientes foi também semelhante à estimada para plantações de eucalipto (Cortez, 1996).
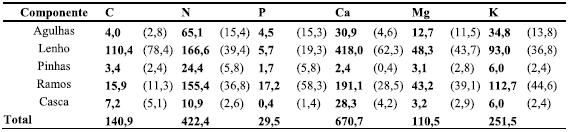
A quantidade de nutrientes na biomassa removida pelos desbastes representou cerca de 25% da quantidade de nutrientes existente na biomassa no final da revolução (Quadros 1 e 2). Trata-se pois de quantidade não negligenciável, indicando a necessidade de ponderar sobre as consequências de manutenção do material de desbaste na estação, no que toca à sustentabilidade do sistema.
]]>Quadro 2 Quantidade (kg ha-1) de nutrientes existentes no material dos desbastes efectuados no povoamento de P. pinaster com 14, 25 e 42 anos de idade,
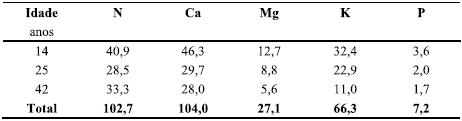
No final da revolução, a quantidade de carbono alocada no tronco (140,9 t ha-1) era quase 80% do total acumulado na biomassa aérea (Quadro 3), como aliás sucede para a generalidade das espécies florestais (Fisher & Binkley, 2000); a proporção respeitante às agulhas, casca e ramos era apenas de 2,8, 5,1 e 11,3%, respectivamente. A quantidade de nutrientes acumulada nas agulhas era inferior à observada para a folhagem de muitas outras espécies, nomeadamente folhosas (e.g. E. globulus; Madeira et al., 2007), e bastante inferior à existente no lenho e nos ramos. A casca, ao contrário do constatado para outras espécies florestais do País (caso do E. globulus; Cortez, 1996) apresentava uma quantidade insignificante de nutrientes, com particular relevância para o Ca.
Quadro 3 Quantidade de C (t ha-1) e de N, P, Mg e Mn (kg ha 1) nas componentes da biomassa da parte aérea do povoamentos de P. pinaster com 83 anos de idade. A percentagem de nutrientes em cada componente está indicada entre parêntesis.
Assim, no caso dos povoamentos de pinheiro bravo, a exportação de nutrientes do sistema está em correspondência sobretudo com a remoção do lenho (a componente económica mais valiosa) e dos ramos; a manutenção destes no sistema significa a retenção de proporção apreciável de nutrientes acumulados na biomassa, nomeadamente no que respeita ao fósforo. O Ca está sobretudo alocado no tronco (62,3%), ao passo que o P se encontra sobretudo nos ramos (58,3%). O N, Mg e K encontravam-se distribuídos em proporção aproximada entre o tronco e os ramos.
]]>Biomassa e nutrientes na vegetação sob coberto
A biomassa da parte aérea da vegetação sob coberto foi bastante apreciável no povoamento mais jovem (8,6 t ha-1), isto é aos 14 anos, mas tendeu a decrescer nos povoamentos mais velhos (4,5-5,4 t ha-1, respectivamente aos 42 e 83 anos), apesar do decréscimo da densidade das árvores (Quadro 4). A mesma tendência foi, aliás, também verificada para qualquer dos nutrientes considerados. A biomassa observada ao longo da revolução foi muito mais elevada do que a determinada no final da primeira e segunda rotação (respectivamente aos 13 e 11 anos) em plantações de eucalipto (Carneiro et al., 2007, 2008) em condições ecológicas similares, nas quais se atingiram valores médios respectivamente de 1,04 e 1,84 t ha-1.
Quadro 4 Biomassa aérea (peso seco a 85ºC) e quantidade de nutrientes da vegetação sob coberto dos povoamentos com 14, 25 e 42 anos, antes de efectuados os respectivos desbastes, e com 83 anos, antes do corte final.
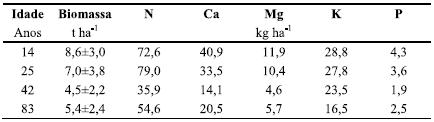
Deve no entanto referir-se que o padrão de variação da biomassa sob coberto ao longo da vida do povoamento foi a normal em povoamentos com boa ocupação do espaço aéreo, com um máximo na fase precoce do seu desenvolvimento, enquanto o índice de área foliar das árvores é sub-óptimo, e novo acréscimo na fase final, com menor densidade das árvores e, consequentemente, menor área foliar. As diferenças observadas em relação às plantações de eucalipto podem atribuir-se ao ritmo de crescimento mais rápido destas (Carneiro et al., 2009) e, eventualmente, a uma menor eficiência das agulhas de pinheiro na intercepção da radiação, quando comparadas com as folhas mais largas dos eucaliptos. Em pinhais de Pinus elliotti da Florida, Gholz & Fisher (1982) assinalaram também máximos de biomassa da vegetação sob coberto em idade mais tardia do povoamento do que se tem observado em Portugal para plantações de eucalipto, nas quais a vegetação espontânea atinge geralmente a maior biomassa antes dos 3 anos, com valores máximos da ordem das 4-5 t ha-1 e decréscimo posterior (Fabião et al., 2002; Carneiro et al., 2009).
Massa e nutrientes das camadas orgânicas
]]> A massa (material isento de cinza) das camadas orgânicas no solo dos povoamentos estudados aumentou claramente com a idade, variando de 9,3 t ha-1, aos 14 anos, para 30,8 t ha-1 no final da revolução. Essa tendência foi contrariada, porém, pelas determinações no povoamento com 42 anos; tal facto deverá estar em correspondência com acções de remoção dos materiais orgânicos da superfície do solo, o que urge averiguar para uma mais correcta interpretação dos dados (Quadro 5).
Quadro 5 Quantidade (média ± 1 desvio padrão) de biomassa e de N, P, Mg, Ca e K nos horizontes orgânicos de povoamentos de P. pinaster com 14, 25, 42 e 83 anos, antes de efectuados os respectivos desbastes ou o corte final.
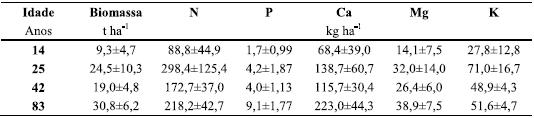
A massa das camadas orgânicas aos 14 anos (8,6±3,0 t ha-1) era mais baixa do que a observada em plantações de eucalipto, tanto na primeira como na segunda rotação (Madeira et al., 1995; Cortez, 1996; Jones et al., 1999; Azevedo, 2000; Magalhães, 2000), isto é, até aos 11-12 anos de idade, o que está em correspondência com a maior rapidez de crescimento desta segunda espécie. A massa observada no final da revolução (83 anos) era muito mais elevada (30,8 t ha-1), enquadrando-se no intervalo (23,552,4 t ha-1) relatado para outros povoamentos de pinheiro bravo, no País, com 56-60 anos (Cortez, 1996; Martins et al., 2007).
A quantidade de nutrientes nas camadas orgânicas, como já foi referido para a massa destas, também aumentou com a idade, atingindo, no termo da revolução, quantidades bastante importantes, como no caso do N (218 kg ha-1) e do Ca (223 kg ha-1); também neste caso a tendência foi contrariada no povoamento com 42 anos de idade (Quadro 5). Não obstante a elevada duração da revolução nos pinhais estudados, a quantidade de nutrientes acumulada nas respectivas camadas orgânicas era da ordem de grandeza ou inferior à determinada nas camadas orgânicas das aludidas plantações de eucalipto; aliás, a grande diferença respeita ao Ca, cuja quantidade era claramente inferior à determinada nas camadas orgânicas no final da primeira rotação de plantações de eucalipto (Jones et al., 1999; Madeira et al., 2007).
Massa volúmica e teor de carbono e nutrientes nas camadas minerais
Algumas das características relevantes dos solos das áreas de estudo estão indicadas no Quadro 6. Os valores dos parâmetros respectivos indicam uma forte variação com a idade dos povoamentos.
]]>Quadro 6 Valores de pH(H2O), massa volúmica (MV) e teores de C e N (g kg-1), valores da razão C/N, concentração debases de troca (cmolc kg-1) e respectiva soma das bases (SB), e de P e K extractáveis (µg g-1) e, no solo, às profundidades de 0-10, 10-20, 20-40, 40-60, 60-80 e 80-100 cm, de povoamentos de P. pinaster com 14, 25, 42 e 83 anos de idade,. Os valores são média ± 1 desvio padrão, n=15.
A massa volúmica aparente entre 20 e 100 cm de profundidade apresentou valores elevados e semelhantes nos vários povoamentos considerados. Porém, os valores na profundidade de 0-10 cm foram muito inferiores aos determinados nas camadas subjacentes e, além disso, diminuíram com a idade dos povoamentos, decrescendo de 1,33 g cm-3, no povoamento com 14 anos, para 0,88 g cm-3 no de 83 anos, o que está em correspondência com o acréscimo dos teores de C orgânico (Quadro 6). Contudo no povoamento com 42 anos de idade verificou-se um desvio a essa tendência, quiçá dependente de possíveis perturbações já referidas a propósito das camadas orgânicas.
O teor de C orgânico na camada 0-10 cm aumentou significativamente com a idade dos povoamentos (r=0,9856; p<0,05), atingindo no respectivo termo da explorabilidade os valores mais elevados; esta tendência indica que o teor do solo em C orgânico e nutrientes é tanto mais elevado quanto maior for a longevidade dos povoamentos. Este padrão significativo não se verificou no caso do N (p=0,1044); aliás, o respectivo teor no povoamento com 42 anos era menor do que no de 25 anos, o que parece corroborar a ocorrência de perturbações, já referidas a propósito das camadas orgânicas que, aliás, têm reflexo na respectiva razão C/N, que era mais elevada do que nos outros casos.
Os teores das bases de troca (Ca, Mg, K e Na), como relatado para o C orgânico, também apresentaram um forte acréscimo ao longo da cronossequência estudada, sobretudo até à profundidade de 20 cm. Os respectivos teores, na camada 0-10 cm, apresentaram uma forte correlação positiva com a idade dos povoamentos (r=0,9900; p<0,01; r=0,9883, p<0,05; r=0,9918, p<0,01; r=0,9541, p<0,05, respectivamente para o Ca, Mg, K e Na). O teor da soma de bases de troca na camada superficial do solo (0-10 cm) aumentou consistentemente com a idade dos povoamentos (sobretudo entre os 42 e 83 anos), sendo cerca de onze vezes maior aos 83 do que aos 14 anos (Quadro 6). O teor de qualquer das bases de troca apresentou o mesmo padrão de variação, mas esta foi um pouco mais acentuada no caso do K e do Na do que no do Ca. A variação mencionada reduziu-se fortemente na camada seguinte (10-20 cm) e esbateu-se nas restantes.
Porém, os valores de pH nas camadas minerais do solo do povoamento com 14 anos eram mais elevados do que os determinados nos povoamentos com 25 e 42 anos, em qualquer das camadas consideradas. Tal diferença poderá atribuir-se à acumulação de matéria orgânica nos segundos, cuja acidez não terá sido compensada pela acumulação de bases de troca. A diferença já não era tão acentuada no final da revolução (83 anos), provavelmente devido à grande acumulação de bases de troca que, entretanto, ocorreu na segunda metade da revolução.
Os teores de K e P extraíveis na camada superficial também apresentaram uma forte correlação positiva com a idade dos povoamentos (respectivamente r=0,9636, p<0,05; r=0,9846, p<0,05), seguindo o padrão observado para o C orgânico e as bases de troca.
Quantidade de carbono e nutrientes
]]> As camadas minerais do solo, até à profundidade de 100 cm, continham no final da revolução 10,8 kg C m-2, o que representa mais do dobro do determinado no povoamento com 14 anos (Figura 1), corroborando a tendência de acumulação de carbono na biomassa e na matéria orgânica do solo dos sistemas florestais até à respectiva exploração (Lal et al., 1998). Porém, a razão entre o C do solo e o da biomassa variou com a idade dos povoamentos, sendo no final da revolução de 0,76, ao pas-so que nas outras idades consideradas variou entre 1,48 e 1,59. O acréscimo de C orgânico até 20 cm de profundidade ocorreu até aos 42 anos, mas continuou até aos 83 anos para a profundidade até 100 cm. O acréscimo de C no solo pode atribuir-se à folhada acumulada superficialmente e, sobretudo, às raízes finas das árvores e da vegetação sob coberto, que são consideradas determinantes na acumulação de C orgânico nas camadas minerais do solo (Ågren et al., 1980; Harris et al., 1980; Fogel,1983; Persson, 1983; Santantonio & Hermann, 1985). Esse padrão está em consonância com os fortes acréscimos observados em plantações jovens de eucalipto, sobre solos semelhantes, em resultado da maior quantidade e turnover de biomassa radical correspondente às raízes finas (Katterer et al., 1995; Madeira et al., 2002).
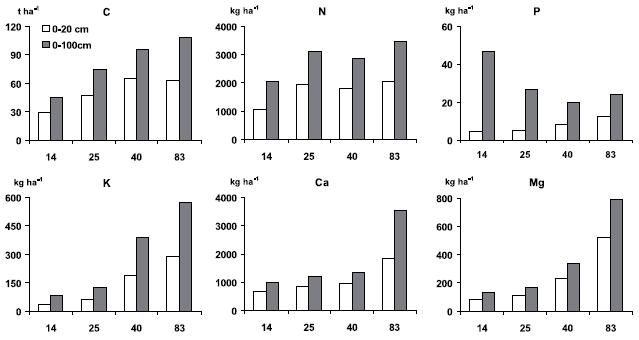
Figura 1 Quantidade de C orgânico (t ha-1) e de N, P extraível e de Ca, Mg e K (kg ha 1) extraíveis nas camadas minerais do solo, até 20 e 100 cm de profundidade, de povoamentos de pinheiro bravo com 14, 25, 42 e 83 anos de idade.
O acréscimo de N no solo não foi tão gradual como o do C orgânico e observou-se sobretudo entre os 14 e os 25 anos (Figura 1), até à profundidade de 20 cm. Não obstante a acumulação de matéria orgânica, a variação da acumulação de N entre os 25 e os 83 anos foi muito reduzida, sugerindo fortes limitações do sistema em N, o que, aliás, está em correspondência com acréscimo da razão C/N daquela (Quadro 6). De facto, as fontes de N durante a revolução foram apenas a deposição atmosférica e a eventual fixação simbiótica do N atmosférico; a primeira deverá ser muito reduzida, como observado noutras áreas próximas ao litoral (Bispo, 2002).
As quantidades das bases de troca, bem como do K extraível pelo método de Egnér-Rihem, também aumentaram com a idade dos povoamentos, sugerindo um forte efeito do ciclo de nutrientes da vegetação arbórea e do sub-coberto na acumulação de nutrientes nas camadas minerais do solo. Ao invés do observado para o C orgânico e o N, o acréscimo das bases de troca ocorreu ao longo da revolução tanto até 20 cm como até 100 cm de profundidade. Tal acréscimo deverá atribuir-se à alteração dos minerais das diversas camadas minerais do solo, a que se deverá somar o efeito dos inputs da atmosfera. O grande acréscimo da quantidade de Ca deverá estar influenciado pelo elevado teor deste elemento relatado para as raízes finas (diâmetro menor do que 2 mm) de pinheiro bravo, em povoamentos instalados sobre solos semelhantes (Ribeiro e Madeira, 1994).
A quantidade de P extraível até 100 cm de profunfidade, pelo contrário, diminuiu com a idade dos povoamentos, sugerindo a transferência, durante o período da revolução, de grande parte da pequena quantidade de P disponível no solo para a biomassa aérea e as camadas orgânicas. O aumento da proporção de P extraível acumulada no solo até 20 cm de profundidade, de cerca de 10% aos 14 anos para cerca de 50%, no final da revolução, corrobora essa transferência.
As variações mencionadas para a quantidade da matéria orgânica e dos vários elementos minerais ao longo da revolução sugere que, aquando da exploração dos povoamentos de pinhal bravo, ocorrerá um forte decréscimo de matéria orgânica e nutrientes. Este padrão não pode ser explicado meramente pela remoção da biomassa e dos resíduos de abate, pois as diferenças das quantidades de nutrientes na biomasa e resíduos orgânicos entre os povoamentos mais jovens e os que se encontram no termo da explorabilidade são muito menores do que as estimadas para os horizontes minerais do solo. Tal facto deverá estar em correspondência com as perturbações inerentes à exploração dos povoamentos (remoção de biomassa, camadas orgânicas, perturbação das camadas superficiais do solo) e sobretudo com a forte modificação das condições de temperatura do solo, o que, como tem sido relatado, tem forte impacte nas condições de mineralização da matéria orgânica e na mobilidade de nutrientes (Fisher & Binkley, 2000); aliás, a textura grosseira (arenosa) do solo que se traduz por fraca protecção da MO do solo e reduzida capacidade de retenção de nutrientes constitui um factor facilitador dessa tendência.
]]> Distribuição do C e nutrientes nas componentes do ecossistema
A distribuição de C orgânico e nutrientes sofre variações com a idade dos povoamentos. Porém, neste estudo apenas se considera a distribuição no termo da explorabilidade, dado a mesma dar indicações orientadoras fundamentais para a salvaguarda de recursos da estação e para delinear os sistemas de gestão visando a sustentabilidade do ecossistema.
No final da revolução a biomassa aérea continha maior proporção de C orgânico (52,8%) do que o conjunto da vegetação espontânea (1,0%), camadas orgânicas (5,7%) e camadas minerais do solo (40,5%) até um metro de profundidade (Figura 2). De acordo com as observações de Madeira et al. (2002) para plantações de eucalipto, a proporção alocada nas camadas minerais do solo seria seguramente maior se a biomassa subterrânea (a única componente que, de facto, não foi medida no presente estudo) fosse considerada. De facto, se assumirmos que a biomassa arbórea subterrânea atinge 20% da total, como é geralmente considerado (Keith et al., 1997), então a proporção de C orgânico no solo aumentaria para 47,5%; se considerarmos ainda o C das camadas orgânicas, então subiria para 52,6%. Esta percentagem é ligeiramente superior à relatada por Madeira et al. (2002) para plantações de eucaliptos com seis anos de idade e instaladas em solos semelhantes (50,9%) e por Fabião et al. (1987) para eucaliptais com 12 anos de idade (51,8%); além disso, é ligeiramente inferior à percentagem relatada por Malhi et al. (1999) para sistemas florestais de regiões temperadas (56%).
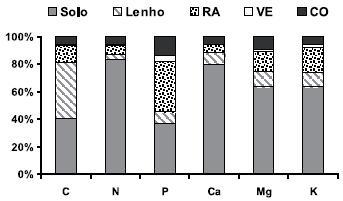
Figura 2 Distribuição do carbono e dos principais nutrientes nos componentes do sistema pinhal bravo, no termo da respectiva revolução, no Pinhal de Leiria.
O N, pelo contrário, estava na sua maior parte alocado nas camadas minerais do solo (83,3%), sendo bastante reduzida a proporção nos resíduos de abate (10,2%) e nas camadas orgânicas (5,2%), seguindo o padrão observado para as plantações de eucalipto (Magalhães, 2000) e de outras espécies (Fisher & Binkley, 2000).
O Ca apresentou um padrão de distribuição semelhante ao azoto (79, 6% alocado nas camadas minerais do solo). O Mg e o K, por seu turno, ocorreram em maior expressão nos resíduos de abate (14,6 e 17,9 % respectivamente) do que no lenho (11,3 e 10,4 %, respectivamente); porém, ainda assim, foi nas camadas minerais do solo que se encontrou a maior proporção (63,4 e 64,0 %, respectivamente) desses nutrientes.
Ao contrário dos nutrientes anteriores, o P sob forma extraível encontrou-se no solo em proporção semelhante (36,8%, Figura 2) à que ocorreu nos resíduos de abate (36,6%); uma proporção apreciável ocorreu no lenho (8,7%), mas ainda assim inferior à observada nas camadas orgânicas (13,9%). Em suma, no sistema em estudo, a retenção dos resíduos de abate e das camadas orgânicas na estação é crucial para minimizar as perdas de P disponível e reduzir o recurso à aplicação de fertilizantes.
]]> A remoção de nutrientes da estação, no final da revolução, através do lenho (Quadro 1) corresponde, exceptuando o Ca, a uma taxa anual inferior aos inputs da atmosfera estimados em áreas próximas do litoral (Bispo, 2002). Aliás, ao contrário do observado para as plantações de eucalipto (Madeira et al., 2007), a remoção do lenho e da totalidade dos resíduos de abate acarreta uma exportação que é da ordem de grandeza ou ligeiramente superior a esses inputs. Assim, tendo em consideração também as entradas devidas à alteração dos minerais do solo, expressas pelos acréscimos de bases com o decorrer da revolução (Quadro 4), os resultados do presente estudo sugerem que a remoção dos resíduos de abate não acarretará perdas sensíveis de nutrientes disponíveis no sistema.Contudo, a remoção da biomassa deve ser equacionada no contexto de um sistema em que, como já foi sublinhado, ocorrem grandes perdas de carbono e de nutrientes devido à perturbação decorrente da exploração florestal, perdas essas facilitadas pelas características do solo. Além disso, se a duração da revolução for reduzida e aumentada a produtividade (Correia & Oliveira, 2003) ocorrerão desequilíbrios entre as entradas anuais e as necessidades das plantas, nomeadamente no caso do P. Em tal cenário, as consequências sobre o stock de nutrientes do solo serão mais acentuadas, sendo a apropriada gestão dos resíduos de abate e dos nutrientes do solo fundamental para garantir a qualidade do solo e a sustentabilidade do sistema.
CONCLUSÕES
No final da revolução dos povoamentos de pinheiro bravo, a biomassa correspondente às agulhas era muito baixa em comparação com a correspondente ao tronco e ramos (cerca de 94%). O lenho e os ramos constituem o grande reservatório de nutrientes na biomassa; a quantidade acumulada nos segundos é de ordem de grandeza ou superior à acumulada no lenho. A quantidade de nutrientes acumulada nas camadas orgânicas e vegetação espontânea é da ordem de grandeza da determinada tanto no lenho, como nos ramos. Verificou-se um elevado acréscimo da quantidade de C orgânico e nutrientes acumulados tanto na biomassa como no solo, sobretudo na segunda metade do período da revolução. A acumulação de N e Mg no solo ocorreu em grande parte até 20 cm de profundidade, enquanto a de C orgânico e Ca teve mais expressão nas camadas restantes. O solo (incluindo as camadas orgânicas) constitui o principal reservatório de nutrientes disponíveis, dependendo, por isso, da sua gestão a fertilidade e a sustentabilidade do sistema; a retenção dos resíduos de abate (sobretudo os ramos) constitui um factor adicional a esses objectivos.
AGRADECIMENTOS
O presente estudo foi efectuado no âmbito do projecto PRAXIS XXI -/3.2/ FLOR/2123/95. Os autores agradecem à Engenheira Susana Gomes, então administradora da Mata Nacional de Leiria, as facilidades na selecção e utilização dos povoamentos, ao pessoal do laboratório de solos do Instituto Superior de Agronomia os processamentos analíticos, aos Bacharéis Paulo Marques e Luís Hilário, do Instituto Superior de Agronomia, bem como às técnicas Rosinda Pato e Maria João Guimarães e aos técnicos Jorge Bandeira e Fernando Santos, da Escola Superior Agrária de Coimbra, o apoio nas amostragens, preparação de amostras e algumas tarefas laboratoriais.
REFERÊNCIAS BIBLIOGRÁFICAS
]]> Ågren, G.I.; Axelsson, B.; Flower-Ellis, J.G.K.; Linder, S.; Persson, H.; Staaf, H. & Troeng, E. (1980) -Annual carbon budget for a Scots pine. In: T. Persson (Ed.) Structure and Function of Northern Coniferous Forests. Swedish Natural Science Research Council. Stockholm. Pp. 307-314.Aguiar, C.; Capelo, J. & Catry, F. (2007) -Distribuição dos pinhais em Portugal. In: J. S. Silva (Ed.) Pinhais e eucaliptais: a floresta cultivada. Col. Árvores e Florestas de Portugal, 4, pp. 89-104. Fundação Luso-Americana para o Desenvolvimento / Público / Liga para a Protecção da Natureza, Lisboa.
Azevedo, A. A. (2000) Estudo da dinâmica do azoto e carbono em plantações florestais intensivas. Dissertação de Doutoramento. Instituto Superior de Agronomia; Universidade Técnica de Lisboa, Lisboa.
Azevedo,A. L. (1944) Estudo de alguns perfis de solo da Mata Nacional de Leiria. Agros 27, 3-6: 140-149.
Bispo, R.M.B. (2002) Meteorologia da camada limite e deposição atmosférica numa superfície vegetal (Eucalyptus globulus). Dissertação de doutoramento. Instituto Superior de Agronomia, Universidade Técnica de Lisboa. Lisboa.
Cabrita, C.F.R. (1956) -Contribuição para o estudo dos solos da mata nacional de leiria. Sua cartografia parcial. Relatório Final do Curso de Engenheiro Silvicultor. Instituto Superior de Agronomia, Universidade Técnica de Lisboa. Lisboa.
Carneiro, M.; Fabião, A.; Martins, M. C.; Cerveira, C.; Santos, C.; Nogueira, C.; Lousã, M.; Hilário, L.; Fabião, André; Abrantes, M. & Madeira, M. (2007) - Species richness and biomass of understory vegetation in a Eucalyptus globulus Labill. coppice as affected by slash management. European Journal of Forest Research 126(4): 475-485.
Carneiro, M.; Fabião, A.; Martins, M. C.; Fabião, André; Abrantes da Silva, M.; Hilário, L.; Lousã, M. & Madeira, M. (2008)- Effects of harrowing and fertilisation on understory vegetation and timber production of a Eucalyptus globulus Labill. plantation in Central Portugal. Forest Ecology and Management 255: 591-597.
Carneiro, M.; Serrão, V.; Fabião, A.; Madeira, M.; Balsemão, I. & Hilário, L. (2009) -Does slash management influence biomass and nutrient accumulation in understory vegetation of Eucalyptus globulus Labill. plantations in a Mediterranean environment? Forest Ecology and Management 257: 527-535.
Cardoso, J.V.C.; Bessa, M.T. & Marado, M.B. (1973) -Carta de Solos de Portugal (1:1 000 000). Agronomia Lusitana, XXXIII (I-IV): 481-602
]]> Correia, A.V. & Oliveira, A.C. (2003) -Principais espécies florestais com interesse para Portugal. Zonas de Influência Atlântica. Estudos e Informação n.º 322. Direcção-Geral das Florestas. Lisboa.Cortez, N.R.S. (1996) - Compartimentos e Ciclos de Nutrientes em Plantações de Eucalyptus globulus Labill. ssp. globulus e Pinus pinaster Aiton. Dissertação de Doutoramento. Instituto Superior de Agronomia, Universidade Técnica de Lisboa, Lisboa.
Costa, J. C.; Espírito Santo, M.D.; Lousã, M.; Rodríguez-González, P.; Capelo, J. & Arsénio, P. (2002) - Flora e vegetação do Divisório Português. Excursão Geobotânica ao Costeiro Português, Olissiponense e Sintra-no. Actas do VII Simpósio da Associação Ibero-Macaronésica de Jardins Botânicos. Instituto Superior de Agronomia, Lisboa. pp. 249-340.
Cowie A.L.; Smith P. & Johnson D. (2006) - Does soil carbon loss in biomass production systems negate the greenhouse benefits of bioenergy? Mitigation and Adaptation Strategies for Global Change 11: 979-1002.
De Leenheer, L.; Van Hove, J. (1958) - Determination de la teneur en carbone organique des sols. Étude critique des méthodes titrimétriques. Pédologie 8: 39-77.
Fabião, A.M.D. (1986) - Contribuição para o estudo da dinâmica da biomassa e da produtividade primária líquida em eucaliptais. Região litoral do centro de Portugal. Dissertação de Doutoramento. Instituto Superior de Agronomia, Universidade Técnica de Lisboa. Lisboa.
Fabião, A., Madeira, M. & Steen, E. (1987) - Root mass in plantations of Eucalyptus globulus in Portugal in relation to soil characteristics. Arid Soil Research and Rehabilitation, 1: 185-194.
Fabião, A.; Martins, M.C.; Cerveira, C.; Santos, C.; Lousã, M.; Madeira, M. & Correia, A. (2002) -Influence of soil and organic residue management on biomass and biodiversity of understory vegetation in a Eucalyptus globulus Labill. plantation. Forest Ecology and Management 171: 87100.
Fisher, R.F. & Binkley, D. (2000) - Ecology and management of forest soils (Third Edition). John Wiley & Sons, New York.
Fogel, R. (1983) - Root turnover and productivity of coniferous forests. Plant and Soil 71: 75-85.
]]> Gholz, H.L. & Fisher, R.F. (1982) -Organic matter production and distribution in slash pine (Pinus elliottii) plantations. Ecology 63: 18271839.Harris, W.F., Santantonio, D. & McGinty, D. (1980) - The dynamic belowground ecosystem. In: Waring, R.H. (Ed.) Forests: Fresh perspectives from ecosystem analysis. Oregon State University Press. Corvallis. pp. 119-129.
INMG (Instituto Nacional de Meteorologia e Geofísica) (1991) - O clima de Portugal. Fascículo XLIX, volume 2 - 2.ª região. Normais climatológicas da região de Ribatejo e Oeste correspondentes a 1951-1980. Lisboa.
Jones, H.E.; Madeira, M.; Herraez, L.; Dighton, J.; Fabião, A.; González-Rio, F.; Fernandez-Marcos, M.; Gomez, C.; Tomé, M.; Feith, H.; Magalhães, M.C. & Howson, G. (1999) -The effect of organic matter management on the productivity of Eucalyptus globulus stands in Spain and Portugal: tree growth and harvest residue decomposition in relation to site and treatment. Forest Ecology and Management 122: 73-86.
Kätterer, T.; Fabião, A.; Madeira, M.; Ribeiro, C. & Steen, E. (1995) - Fine-root dynamics, soil moisture and soil carbon content in a Eucalyptus globulus plantation under different irrigation and fertilisation regimes. Forest Ecology and Management 74: 1-12.
Keith, H.; Raison, R.J. & Jacobson, K.L. (1997) -Allocation of carbon in a mature eucalypt forest and some effects of soil phosphorus availability. Plant and Soil 196, 8199.
Lal, R.; Kimble, J. & Follet, R. (1998) Land Use and Soil C pools in terrestrial ecosystems. In: Lal, R.R.; Kimble, J.M. & Follet, R. F (Eds) Management of carbon sequestration in soil. Advances in soil science. CRC Press, New York, pp.1-10.
Madeira,M.;Araújo,M.C.&Pereira,J.S.(1995) -Effects of water and nutrient supply on amount and on nutrient concentration of litter fall and forest floor litter in Eucalyptus globulus plantations. Plant and Soil 168-169: 287-295.
Madeira, M.; Cortez, N.; Azevedo, A.; Magalhães, M.; C., Ribeiro, C. & Fabião,A. (2007) -As plantações de eucalipto e o solo. In: A. Monteiro Alves, J. Santos Pereira & J. M. N: Silva (Eds) O Eucaliptal em Portugal. Impactes Ambientais e Investigação Científica. ISApress, Lisboa. pp. 137-174.
Madeira; M.; Fabião, A.; Pereira, J. S.; Araújo, M. C. & Ribeiro, C. (2002) - Changes in carbon stocks in Eucalyptus globulus Labill. plantations induced by different water and nutrient availability. Forest Ecology and Management 171: 75-85.
]]> Madeira, M.; Monteiro, F. & Fonseca, M. (2007) -Solos podzolizados em Portugal: ficção e realidade. O Solo a paisagem e o uso da Terra. Encontro Anual da Sociedade Portuguesa da Ciência do Solo, 4-6 de Julho de 2007, p. 27, Vila Real.Magalhães, M.C. (2000) - Efeitos de técnicas de preparação do solo e gestão dos resíduos orgânicos em características físico-químicas do solo de plantações florestais. Dissertação de Doutoramento. Instituto Superior de Agronomia, Universidade Técnica de Lisboa, Lisboa.
Malhi, Y.; Baldocchi, D.D. & Jarvis, P.G. (1999) -The carbon balance of tropical, temperate and boreal forests. Plant Cell Environment 22: 715740.
Martins, A.; Coutinho, J.; Costa, S.; Fonseca, F. & Madeira, M. (2007) -A folhada de quatro povoamentos florestais no Norte de Portugal: Produção e concentração e quantidade de nutrientes devolvidos ao solo. Revista de Ciências Agrárias 30,2: 201-216. [ Links ]
Morais, J. C. (1936) -Geologia e geografia do Pinhal de Leiria. Descrição geológica -geográfica da região do Pinhal de Leiria. Memórias e Notícias (Publicações do Museu Mineralógico e Geológico da Universidade de Coimbra) n.º 9: 4-46. [ Links ]
Nzila J.D.; Boillet J.P.; Laclau J.C. & Ranger J. (2002) - The effects of slash management on nutrient cycling and tree growth in Eucalyptus plantations in the Congo. Forest Ecology and Management 171: 209-221.
Oliveira, A.F. (1967) -Estudo da matéria orgânica nas unidades de solos cartografados em Portugal. Secretaria de Estado da Agricultura, Direcção-Geral dos Serviços Florestais e Aquícolas, Lisboa.
Persson, H. (1983) - The distribution and productivity of fine roots in boreal forests. Plant and Soil 71: 87-101.
Pinto, A. (1939) - O Pinhal do Rei. Subsídios Vol I. Publicado por A. Arala Pinto, Alcobaça.
Póvoas, I. & Barral, M.F. (1992) -Métodos de Análises de Solos. Comunicação do Instituto de Investigação Científica Tropical, série de Ciências Agrárias, 10. Lisboa.
]]> Proe, M.F. & Dutch, J. (1994) -Impact of whole tree harvesting on second-rotation growth of Sitka spruce: the first ten years. Forest Ecology and Management 65: 39-54.Rego, F.C. (2001) Florestas Públicas. Direcção-Geral das Florestas / Comissão Nacional Especializada de Fogos Florestais, Lisboa.
Ribeiro, C. & Madeira, M. (1994) - Root decomposition process in Mediterranean conditions. Transactions of the 15th World Congress of Soil Science. Vol. 4b,pp. 102-103. Commission III: Poster sessions. International Society of Soil Science / Mexican Society of Soil Society, Acapulco.
Ribeiro, C., Cortez, N., Martins, A. Azevedo, A. & Madeira, M. (2007) Decomposição de agulhas de Pinus pinaster e de folhas de Eucalyptus globulus em regiões do interior e do litoral de Portugal. Revista de Ciências Agrárias 30, 2: 142-158. [ Links ]
Santantonio, D. & Hermann, R.K. (1985) -Standing crop, production, and turnover of fine roots on dry, moderate, and wet sites of mature Douglas-fir in western Oregon. Annalles des Sciences Forestières 42: 113-142.
Shammas K.; OConnell A.M.; Grove T. S.; McMurtrie R.; Damon P. & Rance S.J. (2003) -Contribution of decomposing harvest residues to nutrient cycling in a second rotation Eucalyptus globulus plantation in south-western Australia. Biology and Fertility of Soils 38: 228-235.
Smethurst, P.J. & Nambiar, E.K.S. (1990) -Effect of slash and litter management on fluxes of nitrogen and tree growth in a Pinus radiata plantation. Canadian Journal of Forest Research 20: 1498-1507.
Silva F.M.P. & Batalha, M.B.O. (1859) -Memória sobre o Pinhal Nacional de Leiria. Suas madeiras e productos rezinosos.Associação Marítima e Colonial. Imprensa Nacional, Lisboa.
Spangenberg, A.; Grimm, U.; Silva, J.R.S. & Folster, H. (1996) -Nutrient store and export rates of Eucalyptus globulus plantations in eastern Amazonia (Jari). Forest Ecology and Management 80: 225-234.
Stupak I.; Asikainen A.; Jonsell M.; Karltun E.; Lunnan A.; Mizaraité D.; Pasanen K.; Pärn H.; Raulund-Rasmussen K.; Röser D.; Schroeder M.; Varnagiryté I.; Vilkriste L., Callesen I.; Clarke N.; Gaitnieks T.; Ingerslev M.; Mandre M.; Ozolincius R.; Saarsalmi A.; Armolaiti, K.; Helmisaari H.-S.; Indriksons A.; Kairiukstis L.; Katzensteiner K.; Kukkola M.; Ots K.; Ravn H.P. & Tamminen P. (2007) - Sustainable utilisation of forest biomass for energy Possibilities and problems: Policy, legislation, certification, and recommendations and guidelines in the Nordic, Baltic, and other European countries. Biomass Bioenergy 31: 666-684.
]]> Watanabe, F.S. & Olsen, S.R. (1965) Test of an ascorbic acid method for deter-mining phosphorus in water and Na-HCO3 extracts from soil. Soil Society of America Proceedings 29: 677-678.WRB. (2006) - World Reference Base for Soil Resources. 2nd edition. World Soil Resources Reports No. 103. FAO, Rome.
Zbyszewski, G. & Torre deAssunção, C. (1965) -Carta Geológica de Portugal (Escala de 1:50 000). Notícia explicativa da Folha 22-D (Marinha Grande). Serviços Geológicos de Portugal, Lisboa.
1 Instituto Superior de Agronomia, Universidade Técnica de Lisboa, Tapada da Ajuda, 1349-017, Lisboa, Portugal;
2 Escola Superior Agrária de Coimbra, Bencanta, 3040-316 Coimbra;
3 Endereço actual: Universidade Privada de Angola, Campus do Lubango, CP298 Lubango, Angola
Recepção/Reception: 2009.09.04
]]> Aceitação/Acception: 2009.09.15 ]]>