Genome size estimates for six rust (Pucciniales) species
Estimativa do tamanho do genoma de seis espécies de ferrugens (Pucciniales)
Pedro Talhinhas1,2, Ana Paula Ramos2, Daniela Tavares3*, Sílvia Tavares1,* and João Loureiro3
1 Centro de Investigação das Ferrugens do Cafeeiro, BioTrop, Instituto de Investigação Científica Tropical, 2780-505 Oeiras, Portugal
2 LEAF-Linking Landscape, Environment, Agriculture and Food, Instituto Superior de Agronomia, University of Lisbon, 1349-017 Lisboa, Portugal
3 CFE, Centre for Functional Ecology, Department of Life Sciences, University of Coimbra, 3001-401 Coimbra, Portugal. E-mail: sagtavares@gmail.com, author for correspondence
RESUMO
]]> Os fungos que causam ferrugens caraterizam-se pela especialização relativamente ao hospedeiro, pela biotrofia, por possuírem ciclos de vida complexos e grandes genomas. Neste trabalho a citometria de fluxo foi empregue para determinar o tamanho do genoma de seis espécies de fungos Pucciniales (Basidiomycota), Melampsora pulcherrima, Puccinia behenis, P. cichorii, P. pimpinellae, P. vincae e Uromyces dianthi, agentes causais de ferrugem em Mercurialis annua, Silene latifolia, Cichorium intybus, Pimpinella villosa, Vinca major e Dianthus caryophyllus, respetivamente. Com resultados entre 182,1 e 566,4 Mpb/1C, este estudo contribuiu para o conhecimento do tamanho dos genomas na ordem Pucciniales, reforçando a posição deste táxone como o que engloba os fungos com maior tamanho médio de genoma (335,6 Mpb/1C). Este estudo contribui para uma melhor compreensão dos padrões de distribuição de tamanhos de genoma ao longo da filogenia dos fungos, sugerindo uma ligação entre caraterísticas biológicas e o tamanho do genoma. Em particular, os tamanhos dos genomas de fungos Pucciniales variam fortemente dentro do género, mas também diferem de forma vincada dos genomas de outras ordens em Pucciniomycotina que não Pucciniales, sugerindo que a variação do tamanho do genoma possa ser um elemento ativo na evolução dos agentes causais de ferrugens.Palavras-chave: citogenómica, citometria de fluxo, ferrugem, Fungi, tamanho de genoma.
ABSTRACT
Rust fungi (Basidiomycota, Pucciniales), one of the largest groups of phytopathogenic organisms, are characterised by host specialisation, biotrophy, complex life cycles and enlarged genomes. In this work we have used flow cytometry to determine the genome size of six rust species, Melampsora pulcherrima, Puccinia behenis, P. cichorii, P. pimpinellae, P. vincae and Uromyces dianthi, the causal agents of rust on Mercurialis annua, Silene latifolia, Cichorium intybus, Pimpinella villosa, Vinca major and Dianthus caryophyllus, respectively. With results ranging between 182.1 and 566.4 Mbp/1C, this study has contributed to the knowledge of genome sizes in the order Pucciniales, reinforcing this group as the one with the largest average genome size among fungi, with 335.6 Mbp/1C. By comparing genome sizes and their variability across the fungal kingdom, this study has contributed for understanding the patterns of genome size distribution along the fungal phylogeny, suggesting links between biological specificities and genome sizes. In particular, genome sizes of rust fungi vary greatly within genera, but also differ sharply from other non-Pucciniales orders in the Pucciniomycotina, suggesting that genome size variation may be an active element shaping the evolution of rust fungi.
Keywords: cytogenomics, flow cytometry, rust, Fungi; genome size.
Introduction
Rust fungi (Basidiomycota, Pucciniales) are responsible for very important plant diseases, namely rusts on cereals, soybean, coffee, poplar, pines, legumes, rose and other ornamental plants. They comprise one of the largest groups of phytopathogenic organisms, most of them with very narrow host ranges. Such pathological specialisation into such a wide botanical range of hosts (including mosses, ferns, conifers, monocotyledons and dicotyledons) follows elaborated host-pathogen interaction patterns according to the gene-for-gene theory, which in fact was developed based on the flax-rust pathosystem. Nutrients are obtained by the fungus from living host plant cells (biotrophy) through specialised feeding structures (haustoria). Rust life cycles are among the most complex in fungi, with up to five different spore types and often requiring two botanically distinct hosts to complete the life cycle (as revised by Fernandez et al., 2013). Rust fungi are also characterised by unusually large genomes (average 334 Mbp, while the overall average for Fungi is 44.2 Mbp), a trait that may favour adaptation and pathological specialisation (Tavares et al., 2014).
The genera Puccinia, Uromyces and Melampsora encompass approximately two thirds of all recognised rust species, comprising the most notorious rust pathogens in temperate regions, such as the agents of cereal, legume and poplar rusts. So far, the average genome size for species of the genus Puccinia is 248 Mbp, ranging widely across 17 species from 77 Mbp to 806 Mbp, i.e., spanning nearly the entire range of genome sizes reported so far for rusts. In fact, at 806 Mbp, P. chrysanthemi is one of the largest fungal genomes ever reported, in opposition to the cereal rusts (P. triticina and P. graminis f. sp. tritici, both at 77 Mbp) which fall among the smallest genomes (Tavares et al., 2014), but still twice as large as the average genome size for fungi. Similarly, the genome size of Uromyces species ranges from 277 (U. rumicis) to 712 Mbp (U. vignae), with an average of 467 Mbp obtained over seven species. Genome sizes known for six species in the genus Melampsora range from 118 (M. larici-populina) to 333 Mbp (M. ricini), with an average of 221 Mbp (Tavares et al., 2014).
]]> Such enlarged genomes found in rusts may be important sources of diversity, particularly in the absence of sexual reproduction, due to the activity of transposable elements (abundant in these large genomes), and the events leading to genome size variation (namely polyploidisation and chromosome transfer) are thought to be important factors related with host specialisation and even with specific 'host genotype-pathogen race' interactions (Duplessis et al., 2011; Spanu, 2012; Stuckenbrock & Croll, 2014). The knowledge of rust genome sizes, besides being of interest to the field of Mycology, is therefore of relevance to Plant Pathology in particular and to Agronomy in general. In this work we have determined the genome sizes of six species causing rust diseases in ornamentals and weeds, namely M. pulcherrima (Dog's Mercury rust, on Mercurialis annua), P. behenis (White Campion rust, on Silene latifolia), P. cichorii (Chicory rust, on Cichorium intybus), P. pimpinellae (Wild Anise rust, on Pimpinella villosa), P. vincae (Blue Periwinkle rust, on Vinca major) and U. dianthi (Carnation rust, on Dianthus caryophyllus). For that, flow cytometry, the state of the art method for fungal genome size determination (D'Hondt et al., 2011), coupled with an optimized protocol for the analysis of biotrophic fungi (Loureiro et al., 2007; Tavares et al., 2014), was applied.
Materials and Methods
Plant material exhibiting rust symptoms were collected in the Lisbon area during 2014 and 2015 (Table 1). Rust species were recognised upon identification of host species and complemented by the examination of spore morphology. Herbarium specimens were deposited at the João de Carvalho e Vasconcellos Herbarium (LISI; Lisbon, Portugal). Three samples were collected at the teliosporic stage, two at the urediniosporic stage and one at the aecidiosporic stage (Table 1).
Rust genome sizes were estimated by flow cytometry using a Partec CyFlow Space flow cytometer (Partec GmbH, Görlitz, Germany). For such, nuclei were isolated by simultaneously chopping infected leaves of each sample (listed in Table 1) with leaves of the internal reference standard Raphanus sativus ‘Saxa' (2C= 1.11 pg or 1.086 Mbp; (Doležel et al., 1992) and Solanum lycopersicum ‘Stupické' (2C = 1.96 pg or 1.917 Mbp; Doležel et al. 1992), as previously described (Tavares et al. 2014). The nuclear suspension was then filtered through a 30 μm nylon filter to remove plant and fungal debris, and 50 μg/mL of propidium iodide (Fluka, Buchs, Switzerland) and 50 μg/mL of RNase (Fluka) were added to stain DNA, only. Data were acquired using Partec FloMax software v2.4d (Partec GmbH) as previously described (Tavares et al., 2014). For each sample, fluorescence peaks of fungal nuclei were identified by comparing fluorescence histograms of rust-infected leaves with healthy leaves of the host plant.
Results and Discussion
Flow cytometry analyses enabled the separation and identification of nuclei of six rust species by comparison with their host nuclei (except for Mercurialis annua, Silene latifolia and Pimpinella villosa, whose nuclei were out of fluorescence range due to a larger genome size) (Fig. 1). The comparison to the internal reference standard (either R. sativum or S. lycopersicum) permitted the estimation of the genome sizes for the six rust species (Table 1). The genome size of Melampsora pulcherrima (215.6 Mbp), obtained from the Euphorbiaceae Mercurialis annua, is within the range of the genome sizes known for six other Melampsora species (average 221.2 Mbp; Tavares et al., 2014). Among Melampsora spp. infecting Euphorbiaceae, the genome size for M. pulcherrima is lower than that of M. ricini (in Ricinus communis; 332.8 Mbp) and slightly lower than that of M. euphorbiae (in Euphorbia pterococca; 233.8 Mbp). At 418.8 Mbp, the genome size of Uromyces dianthi (from Dianthus caryophyllus) is within the range of the genome size distribution for Uromyces spp. (average 467.5 Mbp; Tavares et al., 2014), and is larger than any of the Uromyces species with non-Fabaceae hosts known so far (as U. transversalis genome size, from Gladiolus sp., is 376.7 Mbp and U. rumicis, from Rumex crispus, is 276.8 Mbp). Ranging from 182.1 to 566.4 Mbp (average 359.4 Mbp), the genome sizes for the four Puccinia species determined in this study are within the range of the 17 Puccinia species with known genome size (average 248.3 Mbp; Tavares et al., 2014). At 321.6 Mbp, the genome size of Puccinia pimpinellae (from Pimpinella villosa) is larger than that of P. smyrnii (at 258.9 Mbp), the other rust from an Apiaceae host with known genome size. At 182.1 Mbp, the genome size of P. behenis (from Silene latifolia) is the smallest determined in this study and, along with U. dianthi, represent the first two genome sizes determined for rusts infecting Caryophyllaceae plants. The genome size of P. cichorii, from Cichorium intybus, is 334.8 Mbp, a value smaller than that determined in rusts found on other Asteraceae, namely Coleosporium inulae (from Dittrichia viscosa; 390.3 Mbp) and P. chrysanthemi (from Dendranthema sp.; 806.5 Mbp). The Vinca major rust, P. vincae, with an estimated genome size of 566.4 Mbp, represents the largest genome in this study, and the first known genome size for a rust infecting an Apocynaceae plant. As before (Tavares et al., 2014), no relationship could be established between the genome size and the type of life cycle of each rust species.
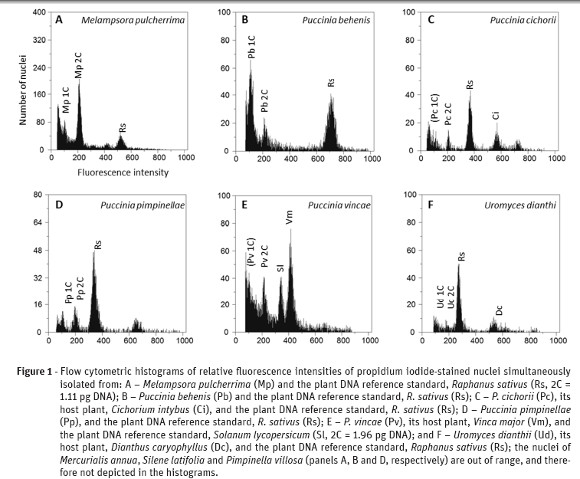 ]]>
]]>
Spanning from 192.2 to 567.1 Mbp, the results obtained in this study raise the number of organisms in the Pucciniales with known genome size to 45, making this the third better documented order in the Basidiomycota for this trait (Fig. 2). The average genome size for rust fungi is now 335.6 Mbp, making this the fungal order with the largest average genome size, namely larger than the Glomeralles (158.6 Mbp) and the Pezizales (89.3 Mbp). Moreover, the variability in genome size values among the Pucciniales is over ten-fold, from 77 to 893 Mbp. Comparing the genome size standard deviation across fungi, the Pucciniales are again the most variable order (Fig. 2). In further detail, the comparison of genome sizes across fungal genera from which there is information available for three or more species (Table 2) shows that Uromyces is the genus with the largest average genome size (461.4 Mbp), followed by the rust genera Puccinia (269.5 Mbp) and Melampsora (219.6 Mbp), and immediately by the Ascomycota genus Octospora (202.7 Mbp), the Basidiomycota genus Gomphidius (199.6 Mbp) and the Glomeromycota genus Glomus (181.2 Mbp). The genus Puccinia is also very variable, with a coefficient of variation across genome size values of 75.7%, second only to the genus Glomus. On the contrary, the genera Melampsora and Uromyces are less variable than many other fungal genera.
Figure 2 and Table 2 also evidence that while some fungal groups are relatively homogeneous concerning genome sizes of their members (e.g., the Ascomycota orders Eurotiales, Helotiales, Hypocreales, Saccharomycetales and Sordariales and the Basidiomycota families Agaricaceae and Boletaceae), others encompass sharp differences. For instance, in the Ascomycota family Pyronemataceae, the average genome size in the genus Octospora (202.7 Mbp) is approximately 8x larger than that of the genus Cheilymenia (25.5 Mbp), and in the Ascomycota family Xylariaceae, the average genome size in the genus Xylaria (60.5 Mbp) is twice as large as in the genus Hypoxylon (26.2 Mbp). At a higher taxonomic level, mixed scenarios also emerge. In the Ascomycota, a trend for very small genomes in sub-phylla Saccharomycotina and Taphrinomycotina is clear and also below-average values are consistent in the class Eurotiomycetes; still a much higher variability is observed within the classes Pezizomycetes and Sordariomycetes. In the Basidiomycota, sharp differences occur among the Agaricomycotina, with smaller genomes in the orders Agaricales, Polyporales and Tremellales, but also in the family Coniophoraceae of the Boletales, and above-average values in several families of the Boletales. Among the Pucciniomycotina, the large genome size values consistently recorded among the Pucciniales contrast sharply with the low genome size values for non-Pucciniales members of the Pucciniomycotina scattered across different orders (not represented in Figs. 2 nor 3 due to insufficient number of organisms represented in each group): Agaricostilbales (Agaricostilbum hyphaenes, 17.9 Mbp); Classiculales (Naiadella fluitans, 52.7 Mbp); Microbotryales (Microbotryum violaceum, 26.1 Mbp); Mixiomycetales (Mixia osmundae, 13.6 Mbp); Sporidiobolales (Rhodotorula graminis, 21.0 Mbp, and Sporobolomyces roseus, 21.2 Mbp) (as revised by Tavares et al., 2014). The average genome size for these non-Pucciniales members of the Pucciniomycotina is 25.4 Mbp. i.e., over 13x smaller than the average genome size of their sister order Pucciniales (335.6 Mbp after this study).
The mixed scenarios found across fungi regarding genome size variation, in some cases with stable genome size values over large taxonomic groups and in other cases with sharp variations within orders, families and genera (or even intraspecific variation) suggest that genome size is an important player of the speciation processes that shaped the fungal phylogeny, namely through polyploidisation, chromosome transfer and/or the activity of transposable elements (Spanu, 2012; Stuckenbrock & Croll, 2014). Although encompassing over 7000 members, the order Pucciniales is unified by singular biological characteristics, contrasting with other orders in the Pucciniomycotina, a situation that is mimicked by the contrast in genome sizes. The specialization of most rust fungi into single or few host species (frequently requiring two phylogenetically distant host species to complete their life cycles), their strict dependency of life host tissues and their gene-for-gene interaction with their hosts, combined with their large but variable genomes, suggest that genome size variation may be an active element shaping the evolution of rust fungi. Such knowledge is of utmost importance for disease resistance breeding programmes for several crops affected by rust diseases, but also for studies addressing the potential use of rusts as biological control agents of weeds.
Conclusions
In this work we used flow cytometry to determine the genome size of six rust species (Melampsora pulcherrima, Puccinia behenis, P. cichorii, P. pimpinellae, P. vincae and Uromyces dianthi), the causal agents of rust on Mercurialis annua, Silene latifolia, Cichorium intybus, Pimpinella villosa, Vinca major and Dianthus caryophyllus, respectively. Ranging between 192.2 and 567.1 Mbp, the values obtained in this study reinforce the order Pucciniales as the one with the largest average genome size across fungi, at 335.6 Mbp, i.e., nearly 8x larger than the average fungal genome. By comparing the genome sizes (and their variability) across the entire fungal kingdom, this study has provided clues for the importance of genome size variation into the shaping of the fungal phylogeny.
Acknowledgments
This work was supported by Fundação para a Ciência e a Tecnologia (FCT, Portugal) through project PTDC/AGR-GPL/114949/2009 and postdoctoral grants awarded to Sílvia Tavares (SFRH/BPD/65965/2009) and Pedro Talhinhas (SFRH/BPD/88994/2012).
]]>References
D'Hondt, L.; Hofte, M.; Van Bockstaele, E. and Leus, L. (2011) - Applications of flow cytometry in plant pathology for genome size determination, detection and physiological status. Molecular Plant Pathology, vol. 12, n. 8, p. 815-828. [ Links ]
Doležel, J.; Sgorbati, S. and Lucretti, S. (1992) - Comparison of three DNA fluorochromes for flow cytometric estimation of nuclear DNA content in plants. Physiologia Plantarum, vol. 85, n. 4, p. 625-631. [ Links ]
Duplessis, S.; Cuomo, C. A.; Lin, Y. C.; Aerts, A.; Tisserant, E.; Veneault-Fourrey, C.; Joly, D. L.; Hacquard, S.; Amselem, J.; Cantarel, B. L.; Chiu, R.; Coutinho, P. M.; Feau, N.; Field, M.; Frey, P.; Gelhaye, E.; Goldberg, J.; Grabherr, M. G.; Kodira, C. D.; Kohler, A.; Kues, U.; Lindquist, E .A.; Lucas, S. M.; Mago, R.; Mauceli, E.; Morin, E.; Murat, C.; Pangilinan, J. L.; Park, R.; Pearson, M.; Quesneville, H.; Rouhier, N.; Sakthikumar, S.; Salamov, A. A.; Schmutz, J.; Selles, B.; Shapiro, H.; Tanguay, P.; Tuskan, G. A.; Henrissat, B.; Van De Peer, Y.; Rouze, P.; Ellis, J. G.; Dodds, P. N.; Schein, J. E.; Zhong, S.; Hamelin, R. C.; Grigoriev, I. V.; Szabo, L. J. and Martin, F. (2011) - Obligate biotrophy features unraveled by the genomic analysis of rust fungi. Proceedings of the National Academy of Sciences of the USA vol. 108, p. 9166-9171. [ Links ]
Fernandez, D.; Talhinhas, P. & Duplessis, S. (2013) - Rust fungi: new advances on genomics and host-parasite interactions. In: Kempken, F. (Ed.) - The Mycota, vol XI - 2nd ed. Application in Agriculture. Berlin, Springer Verlag, pp. 315-341. [ Links ]
]]>Laundon, G.F. (1967) - Terminology in the rust fungi. Transactions of the British Mycological Society, vol. 50, n. 2, p. 189-194. [ Links ]
Loureiro, J.; Rodriguez, E.; Doležel, J. and Santos, C. (2007) - Two new nuclear isolation buffers for plant DNA flow cytometry: a test with 37 species. Annals of Botany, vol. 100, n. 4, p. 875-888. [ Links ]
Spanu, P. D. (2012) - The genomics of obligate (and nonobligate) biotrophs. Annual Review of Phytopathology, vol. 50, May 1, p. 91-109. [ Links ]
Stukenbrock, E. H. and Croll, D. (2014) - The evolving fungal genome. Fungal Biology Reviews, vol. 28, n. 1, p. 1-12. [ Links ]
Tavares, S.; Ramos, A. P.; Pires, A. S.; Azinheira, H. G.; Caldeirinha, P.; Link, T.; Abranches, R.; Silva, M. C.; Voegele, R. T.; Loureiro, J. and Talhinhas, P. (2014) - Genome size analyses of Pucciniales reveal the largest fungal genomes. Frontiers in Plant Sciences, vol. 5, p. 422. [ Links ]
]]>Received/Recebido:2015.02.28
Accepted/Aceite: 2015.05.19
]]>