










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias vol.36 no.2 Lisboa abr. 2013
Aluminium tolerance in bean traditional cultivars from Madeira
Tolerância ao alumínio em cultivares tradicionais de feijoeiro da Madeira
Ana Maria Domingues1, Emanuel da Silva2, Gregório Freitas2, José Filipe Ganança2, Humberto Nóbrega2, Jan J. Slaski2 and Miguel Ângelo Pinheiro de Carvalho2
1 Instituto de Investigação Científica Tropical/ Tropical Research Institute, Biotrop -Pólo Mendes Ferrão, Fitoquímica, Tapada da Ajuda, Apartado 3014, 1301-901 Lisboa Codex, Portugal. E-mail: dam.portugal@hotmail.com, author for correspondence
2 ISOPlexis Germplasm Bank, University of Madeira, 9000-390 Funchal, Portugal.
3 Bioresource Technologies, Alberta Innovates – Technology Futures, Vegreville, AB, Canada.
ABSTRACT
Common bean (Phaseolus vulgaris L.) is the most important legume crop in the world, providing low-cost, high quality protein, minerals and dietary fiber for human nutrition. The crop was originated from diversity centers in America and exhibits adaptation abilities to different environmental conditions, including soil with low pH. Acid soils occupy 30% of the agro ecosystem areas in the world. In Madeira, acid Andosols and unsatured Cambisols are the dominant groups of soils. Generally, under acidic and infertile conditions, besides of H+ toxicity, soluble aluminium (Al) is the most important abiotic factor limiting plant development and crop productivity. In the field, the hidden roots are also affected and the reduction of root growth under Al stress can be clearly observed in early stages. Seedlings of fifty bean accessions from the Archipelago of Madeira were tested under controlled conditions in the presence of 50 µM Al at pH 4.4. In general, the tested germplasm appeared to be sensitive or very sensitive to Al toxicity. However, fifteen traditional cultivars clearly exhibited elevated Al-tolerance, with an average root relative elongation (RRE) exceeding 50%, while top six accessions surpassed the 60% RRE mark. The Madeira bean germplasm is a valuable resource for sustainable crop production in acid soils and it could be used as parental lines in breeding programs aimed for Al tolerance in common beans.
Keywords: Acid soils, aluminium toxicity, bean, Phaseolus vulgaris
RESUMO
O feijão (Phaseolus vulgaris L.) é a leguminosa mais importante do mundo, fornecendo proteína, sais minerais e fibra dietética de baixo custo e elevada qualidade para a nutrição humana. A cultura foi originada em centros de diversidade do continente americano e exibe capacidade de adaptação a condições ambientais diversas, incluindo a solos com baixo pH. Os solos ácidos ocupam 30% das áreas agro-florestais do globo terrestre. Na Madeira, os Andossolos ácidos e os Cambissolos insaturados constituem os grupos dominantes de solos. Geralmente, em condições de acidez e infertilidade, além da toxicidade do hidrogenião (H+), a presença de alumínio solúvel (Al) na solução do solo constitui um dos factores abióticos mais limitantes do desenvolvimento e rendimento das culturas. No terreno, as raízes ocultas são também afectadas e a redução do crescimento radicular pode ser observada muito precocemente nas plantas jovens. Foram testadas cinquenta cultivares tradicionais de feijoeiro do Arquipélago da Madeira, sob condições controladas, em relação à tolerância a 50 µM Al com pH 4.4. Das cultivares tradicionais estudadas, a maioria mostrou ter sensibilidade moderada ou elevada à toxicidade do Al. Contudo, quinze evidenciaram tolerância à toxicidade do Al, expressa pela média do alongamento relativo da raiz principal (RRE) superior a 50%, enquanto seis alcançaram valores acima de 60%. O germoplasma de feijão da Madeira constitui um recurso valioso em agricultura sustentável em solos ácidos e poderá contribuir para o melhoramento genético da cultura relativamente à tolerância ao Al.
Palavras-chave: Feijão, Phaseolus vulgaris, solos ácidos, toxicidade de alumínio
Introduction
Plant biodiversity offers more than 50000 edible plant species, but from the 7000 domesticated species only 250 are generally consumed by mankind. Many of these crops were domesticated in the Vavilovs centers of diversity (Vavilov, 1987). Common bean was originated in America, from where the crop was disseminated throughout the world by the Portuguese and Spanish settlers. Since the beginning of the Portuguese Discoveries in the XV century, the initially uninhabited Atlantic Islands, including the Archipelago of Madeira, were convergence points for plants from the entire globe, and among them, an undetermined number of bean species and varieties.
Common bean (Phaseolus vulgaris L.) is the most important legume crop in the world, due the excellent nutritional characteristics and inherited ability to adapt to different environmental conditions. The seeds can be easily stored and transported, in dry conditions at room temperature, for extended period of time and thus they provide nourishment during food scantiness. Beans are a low-cost source of high quality protein (20-25% in dry weight basis) and dietary fiber. Dry beans are rich in calcium, iron, potassium, selenium, molybdenum, and other minerals, as well as vitamins. Proteins are rich in the essential amino acids, such as leucine (and isoleucine), phenylalanine, threonine, histidine, valine, and particularly lysine, but have reduced levels of methionine and cysteine. On the other hand, cereals have high levels of these sulphur-rich amino acids, but have low lysine content. Therefore, the beans are the perfect combination with cereals in balanced diets for human nutrition (Ribeiro et al., 2007). More recently, there is renewed interest for the consumption of local pulses, in particular common bean, because it is more ecologically efficient and has lower fat content compared with animal protein sources, especially meat products which mass production releases higher levels of greenhouse gases with negative effect on global warming. On the other hand, beans contain several secondary metabolites that are considered anti-nutritional factors but, generally, these negative effects can be easily neutralized by proper cooking procedures and rational consumption, and are currently marketed as functional foods and nutraceuticals ingredients (Campos-Vega et al., 2010).
The CIAT (Centro Internacional de Agricultura Tropical/ International Center for Tropical Agriculture) maintains the worlds largest collection of Phaseolus germplasm and supports important efforts towards the bean improvement for the small-scale farmers. The use of high-yielding bean varieties bearing resistance to pests and diseases and tolerance to abiotic stress factors, and the efficient use of well adapted symbiotic N2 fixing-bacteria, could overturn the productivity declines and contribute to eradication of malnutrition and poverty, specially in developing countries, within the Millennium Development Goals (Ishitani et al., 2004; Hillocks et al., 2006; Manrique-Carpintero et al., 2007).
Acid soils occupy 30% of the agro-ecosystem areas (von Uexküll and Mutert cited by Domingues et al., 2004). In acid soils, soluble aluminium (Al) toxicity has been identified as one of the most important limiting factors to plant growth and production. In low input subtropical agrosystems, plant performance and yields depend on the genetic mechanisms of Al resistance and the early response to the presence of Al in root medium (Barceló and Poschenrieder, 2002). In common bean, Cumming et al. (1992) found that the three week treatment with 10 µM Al induced shoot and root growth enhancement of the Al tolerant cultivar Dade (111.2% and 126.8% of control, respectively), while decreased the growths in the cv. Romano (70.7 % and 39.0 % of control, respectively). In sustainable agriculture, the approach to cope with Al toxicity should be the use of selected genotypes, within broad gene pool diversity, and the optimization of nutrients cycling (Hillocks et al., 2006). Breeding programs for particular edaphoclimatic conditions must be realized in the proper environmental context, over the course of several years to achieve crop yield stability and to overcome climate changes (Ceccarelli, 2009). Furthermore, the good efficiency of the Mendelian genetic breeding can be greatly enhanced by incorporating functional, comparative and structural molecular genomics (Ishitani et al., 2004), as criteria in marker assisted selection (MAS) of parental germplasm.
There are strong indications that common bean was initially introduced to Madeira in the 17th century from Brazil and other South America countries (da Silva et al., 2010). For centuries, the Madeira farmers cultivated beans or intercropped beans with maize in isolated terraces constructed along the sloppy hills, with acid and very acid, Al and Fe-rich soils. Successive introductions by the settlers and a positive selection for the most adapted individuals to the climatic and edaphic conditions secondarily increased the crop diversity (Freitas et al., 2011). As a consequence, a great number of local bean varieties showing ability to tolerate biotic and abiotic stress, and exhibiting high yield stability, are still found today in the region. Based on the morphological and agronomic characteristics (Freitas et al., 2011) and the phaseolin pattern data (da Silva et al., 2010), the beans accessions from Madeira can be classified as landraces closely related to the Andean races identified by Singh et al. (1991). Probably in the prevalently acid Andosols and unsatured Cambisols of Madeira Archipelago, the Al phytotoxicity effects (Taylor, 1988) might occur, but only when soil organic matter levels are low (Madeira et al., 1994). Accordingly, in low input agriculture, under soil acidic conditions at pH 4.4, the decomposition products of organic residues decreased Al concentration in the soil solution, diminishing Al in exchangeable cations, furnishing plant nutrients (N, P, Ca, Mg, K) and attenuating the metal toxicity effects to bean and maize (Wong et al., 1995). The presence of 50 µM Al at pH 4.5 in nutrient solution allowed the identification of Al resistance in common bean genotypes using the percent inhibition of primary root elongation, percent increase of average root diameter, total root length per plant and total number of root tips per plant (Manrique-Carpintero et al., 2007).
In the present work, we developed an inexpensive, precise, and non-destructive method to test seedlings of bean in diluted nutrient solutions of pH 4.4, under controlled conditions, during 72 hours. This screening test mimics conditions of many unfertile acid soils around the globe and maximizes the discrimination of Al tolerance behavior of the bean traditional cultivars from Madeira based on root elongation under Al stress.
Materials and Methods
Fifty traditional cultivars of Madeira beans (Phaseolus vulgaris L., tribe Phaseoleae, family Fabaceae), accessions from the germplasm collection of the ISOPlexis Genebank at the University of Madeira in Funchal (at 32° 38 N and 16° 54 W), have been screened for aluminium tolerance. The seeds were scarified with soft sand-paper, disinfected for 15 minutes with sodium hypochlorite (a solution containing 5 g active choride per liter), washed extensively with distilled water, and germinated between cotton and moistened filter paper in the dark at 22°C, for 4 days. The seed size categories were grouped, according to the weight of one hundred seeds (Voysest cited by Freitas et al., 2011), as: large (> 40 g/ 100 seeds), medium (25-40 g/ 100 seeds) and small (< 25 g/ 100 seeds). All local cultivars exhibited large seeds, except for the ISOP 480, Feijão Preto, with medium size seeds (Table 1.)
Uniform seedlings with radicle length between 1.5 and 7 cm were selected, placed on the floats and fixed with cotton in the area of the neck. Seedlings were kept for one day in complete nutrient solution (Slaski et al. cited by Domingues et al., 2004). After, floats with plants were transferred to containers with treatment solution containing 1000 µM Ca(NO3)2.4H2O, 300 µM Mg(NO3.6H2O, 300 µM ) 2NH4NO3, with the addition of 50 µM AlCl3.6H2O in the experimental variants. During the treatment period, potassium, phosphorus, sulfate, iron, boron and other micronutrients were not included in the diluted solutions to minimize the direct and indirect reduction of the phytotoxic effect of the Al ions. All nutrient solutions were replaced by fresh ones after 46 hours. Each container with 15 liters of constantly aerated treatment solution housed 120 plants, including 2-4 plants of each accession. Four containers, one with 0 µM Al (control) and three with 50 µM Al, with the pH of solutions strictly maintained at pH 4.40 ± 0.20 by addition of diluted HCl, were used in the 72 hours experiments (Figure 1). The photoperiod was 10 hours of daylight, with constant light intensity at the leaf level, provided by natural light supplemented with fluorescent and incandescent lamps. The temperature of the nutrient solution ranged from 20° ± 2° C during the night and 26° ± 2° C at daytime.

The primary root of each plant was manually measured at 0, 24, 48 and 72 hours of experimental treatment, corresponding to the L0, L24, L48 and L72 lengths, expressed in mm, with accuracy of 0.5 mm. The absolute root elongation (E) was calculated for each plant, in the given time periods and expressed as mm of elongation (E24 = L24-L0; E48 = L48-L0; and E72 = L72-L0). The average of the absolute elongation (AE) of root in control solution was calculated for the time periods considered for the experiments – AE24T0, AE48T0, and AE72T0 (for each accession n = 14).
The individual values of relative growth rate (rgr) were calculated in the three periods and expressed as mm mm-1 day-1 [rgr24 = E24/L24; rgr48 = E48/(L48 x2); and rgr72 = E72/(L72 x3)]. The average of relative elongation rate (RER) of every accession was calculated for the time periods considered (average values RER24, RER48, and RER72), for both control and stressed plants.
The individual values of the Al treatment absolute root elongation were divided by the accession average elongation in the control solution, expressed as percentage, after 24, 48 and 72 hours of treatment (individual values in the time period considered i, as rre i = Ei/AEiT0x100). Data were analyzed and reduced for statistical purposes to twenty plants for each accession by the elimination of extreme values, and the average of the root relative elongation (RRE, n=20) was calculated for each time period – RRE24, RRE48, and RRE72.
The existence of significant differences among accessions was determined by one-way ANOVA using the Statgraphics Program. Homogeneous subsets were established by the LSD test, which sorted the accessions in statistical classes of decreasing Al-tolerance (Table 1).
Results
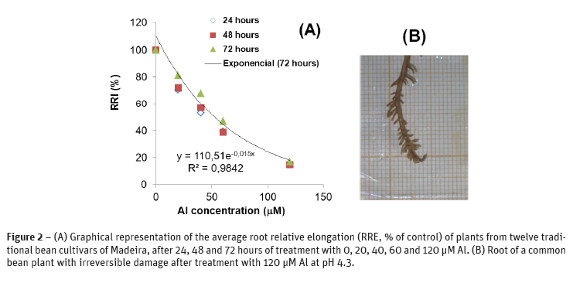
Several previous experiments were performed under similar conditions to establish the best discrimination among the bean accessions. In a preliminary experiment, we determined that bean seedlings were able to increase the initial solution pH adjusted to 4.3 by 0.2 points, after 12 hours in the absence of Al, but the change was smaller under Al stress (data not shown). Another preliminary experiment performed on 12 accessions revealed that the root relative elongation of plants decreased exponentially with increasing concentrations of Al in the root medium after 24 to 72 hours (Figure 2-A) and, the plants exhibited severe root damage in the treatment 120 µM Al at pH 4.3 ± 0.2 (Figure 2-B). In these exploratory experiments, the symptoms of Al toxicity in bean roots were clearly visible as reduced length of the primary and secondary roots, scanty ramification, apices swelling, and brownish color. Without Al, the roots remained white and secondary ramification was abundant at pH 4.3 ± 0.2. The primary root from the stressed plants exhibited significant reduction of total length after 24, 48 or 72 hours in presence of 50 µM Al, when compared with the control plants. These results reflect the sensitivity of the root apices to grow under Al stress, as referred by Taylor (1988), because the use of control with the nutrient solution at the same pH conditions of the experimental variant allow us to discard the influence of H+ toxicity. The root inhibition was reversible after 50 µM Al treatment, because during regrowth in a complete nutrient solutions (approx. at pH = 4.4) for 24 hours, the primary roots continued to grow and developed exuberant secondary ramification (data not shown).
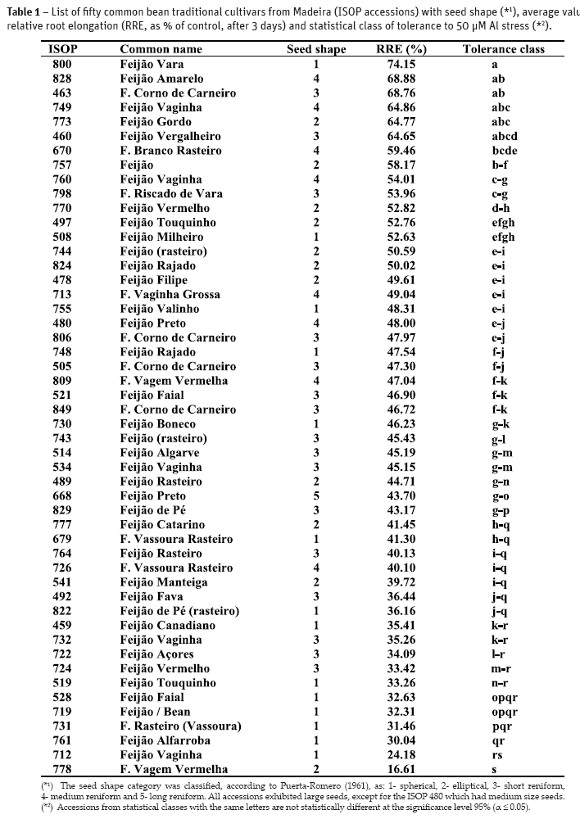
Among the four variables tested for Al tolerance the average of relative root inhibition after 72 hours of treatment (RRE72) permitted the best discrimination of the screened germplasm. The Table 1 shows fifty traditional bean cultivars from Madeira arranged in the decreasing order of Al tolerance evaluated by the inhibition of primary root elongation (RRE72) in presence of Al in the root medium. Additional information about the seed shape and size is also provided to illustrate the variability among the bean accessions.
Fifteen traditional bean cultivars from Madeira clearly exhibited Al-tolerance with RRE72 above 50%. But only six accessions exhibited statistically superior Al-tolerance, with RRE72 above 64% (ISOPs 460, 463, 749, 773, 800 and 828). The majority of the accessions were classified as sensitive with RRE72 ranging from 25 to 50%. A great variability in the response to Al stress was observed among and inside the traditional cultivars, with the most resistant ISOP 800 having RRE72 of 74.15%, while the most sensitive ISOP 778 had merely 16.61% RRE72 (Table 1).
Discussion
In soils of volcanic origin, particularly those located at higher elevations, agriculture in Madeira Island frequently faces challenges associated with unique edaphoclimatic conditions (Ganança et al., 2007).
The Archipelago of Madeira has temperate climate strongly influenced by oceanic and orographic conditions, which define warmer microclimatic spots adequate for cultivation of the cold sensitive bean crop. The high annual precipitation, often exceeding 2000 mm per annum in the mountains, enforces soil lixiviation and water erosion. The majority of FAO soil groups on the Island are acid, characterized by low concentration of exchangeable bases (as K+, Ca2+, and Mg2+), with high levels of Al and iron compounds, and deficit of exchangeable phosphorus, especially in deeper soil.
Al and Fe ions can form complexes to a considerable extent with humic substances, as the superficial horizons are rich in organic matter and continuously developed at least down to 40 cm depth (Madeira et al., 1994). As a result of terrace construction, particularly in Andosols, the resulting fields probably have disturbed pedon with the deeper and very acid soil layers mobilized to the top. Therefore, that anthropogenic activity most likely increased the selection pressure for H+, Al, and Fe tolerance of bean - the orphan crop, traditionally managed by small farmers (Ceccarelli, 2009).
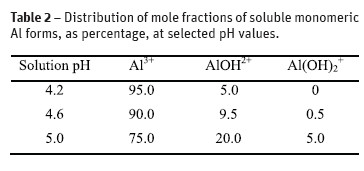
The octahedral hexahydrate trivalent cation (Al (H2O)63+, abbreviated as Al3+) is the most toxic, soluble monomeric form of Al prevailing in aqueous solutions below pH 5.0. As the solution pH rises, Al3+ undergoes successive deprotonations to less phytotoxic Al ions and Al(OH)3, which precipitates near neutral pH. When the solution pH rises from pH 4.2, to 5.0 the distribution of molar fractions (expressed as percentage) of soluble forms Al3+, AlOH2+ and Al(OH)2 + change (Martin, 1988) - Table 2. Several authors (Martin, 1988; Rangel et al., 2005; Poschenrieder et al., 2008) reported that toxicity of the hydrogen ion (H3O+ or, proton, H+) interferes with chemical activities of the Al ions and can modulate their toxic effects on plant roots. Among the edible legume crops (from Fabaceae), the sensitivity to H+ increased in the order pea (Pisum sativum) < soyabean (Glycine max) < common bean (Phaseolus vulgaris), ranging from 84% to 35% of relative root elongation, when solution pH was changed from 6.0 to 4.05 (Lazof and Holland cited by Rangel et al., 2005). The authors suggested that screening of bean for Al tolerance should be done using the root recovery from Al stress approach, whereas others (Rangel et al., 2005; Manrique-Carpintero et al., 2007) proposed the evaluation of root elongation in treatment solution at pH 4.50, in the presence of 5 µM Ca2+ and 0.5 µM K+. However, high concentrations of these cations reduce Al ionic activity in aqueous solutions and its uptake by the roots. Besides, as a consequence of proton toxicity, the primary root is often shorter, but the number of secondary (and tertiary) roots is higher, as a compensation response of the plant to maintain the nutrients uptake. In the present screening, the induction of root ramification was observed both in 0 and 50 µM Al treatments, however secondary roots were much longer without Al presence in the solution. In addition, in the unsatured soils of Madeira Island, the bean plants of any traditional cultivars have to cope simultaneously with high H+ and Al concentrations to produce stable yields. Al phytotoxicity effects could occur only when soil organic matter levels are sufficiently low (Madeira et al., 1994). For these reasons, the present work was performed in diluted solutions with the pH value near 4.4, approximately equivalent to 40 µM H+ in solution.
In common bean (Cumming et al., 1992), the resumption of root elongation was accompanied by increased rates of root respiration and a larger proportion of metabolic energy from root respiration was allocated to maintenance processes against Al toxicity in the Al tolerant genotype Dade. Hence, maintenance of constant aeration of root medium is essential for proper screening for Al tolerance in hydroponics.
The observed brownish color of the roots was probably a result of production and accumulation of phenolic compounds. The formation of these compounds is induced under Al stress, due to their implication in complex formation with Al or stability, after the increase of the Al-organic acid anion complexes used to neutralize metal toxicity (Barceló and Poschenrieder, 2002).
As other cross pollinated crops, beans generally show great genetic variability and phenotypic plasticity assisting its adaptation to different edaphoclimatic conditions. As expected, these traditional bean cultivars were characterized by different morphological, agronomic or biochemical descriptors and markers (Silva et al., 2010; Freitas et al., 2011).
Similar variability was observed with respect to Al tolerance ((Table 1). For example, the ISOPs 724 and 770, both Feijão Vermelho, collected in S. Vicente and Porto Moniz, in the North of Madeira Island, with large size seeds and variegated red coat, mainly composed by the Contender phaseolin type (da Silva et al., 2010), exhibited respectively 33% and 53% RRE72. Among the five Feijão Vaginha cultivars, the Al-tolerant ISOP 749 with approximately 65% RRE72 was statistically superior to the ISOPs 712 and 732 with 24 and 35% RRE72, respectively (Table 1, a = 0.05).
In an earlier work (Domingues et al., 2004), forty four traditional maize cultivars from the ISOPlexis Bank were screened for Al tolerance. When submitted to 100 µM Al treatment, at pH 4.3 the values of RRE72 ranged from 82% to 25%. Maize exhibited higher Al tolerance than beans, but also great variability within the traditional cultivars. In Madeiras traditional agricultural system, the bean intercrops with the maize benefits the bean production, as maize roots might rise the pH and exudate organic acids that complex Al ions more actively than beans. Also, the symbiosis with N2-fixing bacteria becomes more important to satisfy the N needs of both crops. As a consequence of N2 fixation, there is an acidification trend in soils bearing legume species. Therefore, in bean improvement programs selection for proton and Al tolerant Rhizobium spp. should also be conducted (Graham et al., 2003; Hillocks et al., 2006). In well drained fields, the surplus of ammonia (NH4 +) released by the legume roots is oxidized to nitrate (NO3 -), and the anionic N uptake by the roots of other crops induces the pH rise in the rhizosphere, which in turn causes precipitation and detoxification of soluble Al outside of the roots (Taylor, 1988). Moreover, the inclusion of land covering crops and polyculture are advisable, principally in sustainable agro-ecosystems with less inputs and in situations of high water erosion risk, as proven by the ancestral companion plant system of maize-bean-squash (Hart cited by Graham et al., 2003).
Generally, the holistic strategy of several physiological processes contributes to the successful performance of potentially Al tolerant genotypes (Taylor, 1988). The germplasm screened by the RRE72 (Table 1) should be further studied to determine the physiological mechanisms of Al tolerance and to find molecular Al tolerance markers (Ishitani et al., 2004). Verification of the data generated by this screening test ought to be conducted in toxic Al rich soils to evaluate pods or grain yield since for tolerance using root elongation indices as a single criterion might be misleading in bean genotypes, which accumulate high amounts of Al in the shoots (Mas-sot et al., 1992). Due to the potential of allocation of Al into the shoot, a proper selection of the cultivars must be performed to avoid the accumulation of Al in the seeds, as beans are used as staple food. The nutritional characterization of these traditional cultivars might reveal valuable traits in future investigations.
Conclusions
The Al tolerance indices based on the present methodology of root relative elongation discriminated well the traditional bean cultivars of Madeira.
In hydroponics with 50 µM Al at pH 4.4, using the RRE72 index, six traditional cultivars among the selected fifty bean accessions from Madeira Island were identified, with statistically higher Al tolerance (the ISOPs 460, 463, 749, 773, 800 and 828).
This germplasm is a very valuable resource for sustainable crop production in acid and Al toxic soils of the region. These traditional cultivars maintained in the ISOPlexis Gene Bank at the University of Madeira could also serve as donors of tolerance genes in bean breeding programs worldwide.
Bibliographic References
Barceló, J. and Poschenrieder, C. (2002) - Fast root growth responses, root exudates, and internal detoxification as clues to the mechanisms of aluminium toxicity and resistance: a review. Environmental and Experimental Botany, 48, 1: 75-92. [ Links ]
Campos-Vega, R.; Loarca-Piña, G. and Dave Oomah B. (2010) - Minor components of pulses and their potential impact on human health. Food Research International 43, 2: 461-482. [ Links ]
Ceccarelli, S. (2009) - Evolution, plant breeding and biodiversity. Journal of Agriculture and Environment for International Development, 103, 1-2: 131-145. [ Links ]
Cumming, J.R.; Cumming, A.B. and Taylor, G.J. (1992) – Patterns of root respiration associated with the induction of aluminium tolerance in Phaseolus vulgaris L. Journal of Experimental Botany, 43, 253: 1075-1081. [ Links ]
da Silva, E.M.; Correia, A.S.A.; Lopes, N.A.A.; Nóbrega, H.G.M.; Ganança, J.F.T.; Domingues, A.M.; Khadem, M.; Slaski, J.J. and Pinheiro de Carvalho, M.A.A. (2010) -Phytogeographical origin of Madeiran common beans based on phaseolin patterns. Pesquisa Agropecuária Brasileira, 45, 8: 863-871. [ Links ]
Domingues, A.M.; Fernandes, M.L. and Nunes, M.A. (2004) - Biodiversity of maize from Madeira Island to aluminium tolerance. In: Proceedings of 2nd Symposium of Island Ecosystems. CEM, Liberal, Funchal, p. 15-24. [ Links ]
Freitas, G.; Ganança, J.F.T.; Nóbrega, H.; Nunes, E.; Costa, G.; Slaski, J.J. and Pinheiro de Carvalho, M.Â.A. (2011) -Morphological evaluation of common bean diversity on the Island of Madeira. Genetic Resources and Crop Evolution 58, 6: 861-874. [ Links ]
Ganança, J.F.T.; Abreu, I.; Sousa, N.F.; Paz, R.F.; Caldeira, P.; dos Santos, T.M.M.; Costa, G.; Slaski, J.J. and Pinheiro de Carvalho, M.Â.A. (2007) – Soil conditions and evolution of aluminium resistance among cultivated and wild species on the Island of Madeira. Plant, Soil and Environment, 53, 6: 239-246. [ Links ]
Graham, P.H.; Rosas, J.C.; Estevez de Jensen, C.; Peralta, E.; Tlusty, B.; Acosta-Gallegos, J. and Arraes Pereira, P.A. (2003) -Addressing edaphic constraints to bean production: the bean cowpea CRSP project in perspective. Field Crops Research, 82, 2-3: 179-192. [ Links ]
Hillocks, R.J.; Madata, C.S.; Chirwa, R.; Minja, E.M. and Msolla, S. (2006) -Phaseolus bean improvement in Tanzania, 1959-2005. Euphytica, 150, 1-2: 215-231. [ Links ]
Ishitani, M.; Rao, I.; Wenzl, P.; Beebe, S. and Tohme, J. (2004) - Integration of genomics approach with traditional breeding towards improving abiotic stress adaptation: drought and aluminium toxicity as case studies. Field Crops Research, 90, 1: 35-45. [ Links ]
Madeira, M.A.; Furtado, E.; Jeanroy, E. and Herbillon, A.J. (1994) - Andisols of Madeira Island (Portugal). Characteristics and classification. Geoderma, 62, 4: 363-383. [ Links ]
Manrique-Carpintero, G.; Rao, I. and Beebe, S. (2007) -Identification of aluminium resistance in common bean genotypes using a hydroponic screening method. 2007 External Program and Management Review (EPMR), poster 20. (Acessed 2011. 12.8. Disponível em: http://webapp.ciat.cgiar.org/epmr_ciat/pdf/poster_20_epmr07.pdf. [ Links ]
Martin, R.B. (1988) -Bioinorganic chemistry of aluminium. In: Sigel, H. and Sigel, A. (Ed.) – Metal ions in biological systems: Volume 24: Aluminium and its role in biology. New York, Marcel Dekker, p. 1-57. [ Links ]
Massot, N., Poschenrieder, C. and Barceló, J. (1992) – Differential response of three bean (Phaseolus vulgaris) cultivars to aluminium. Acta Botanica Neerlandica, 41, 3: 293-298. [ Links ]
Poschenrieder, C.; Gunsé, B.; Corrales, I. and Barceló, J. (2008) - A glance into aluminium toxicity and resistance in plants. Science of the Total Environment, 400, 1-3: 356-368. [ Links ]
Puerta-Romero, J. (1961) - Variedades de judías cultivadas en España. Nueva classificación de la espécie Phaseolus vulgaris (L ex. p.) Savi. Madrid, INIA, 798 p. (Monografías INIA, 11). [ Links ]
Rangel, A.F.; Mobin, M.; Rao, I.M. and Horst, W.J. (2005) – Proton toxicity interferes with the screening of common bean (Phaseolus vulgaris L.) genotypes for aluminium resistance in nutrient solution. Journal of Plant Nutrition and Soil Science,168, 4: 607-616. [ Links ]
Ribeiro, N.D.; Londero, P.M.G.; Cargnelutti Filho, A.; Jost, E.; Poersch, N.L. and Mallmann, C.A. (2007) – Composição de aminoácidos de cultivar-es de feijão e aplicações para o melhoramento genético. Pesquisa Agropecuária Brasileira, 134, 10: 1393-1399. [ Links ]
Singh, S.P.; Gepts, P. and Debouck, D.G. (1991) -Races of common bean (Phaseolus vulgaris, Fabaceae). Economic Botany, 45, 3: 379-396. [ Links ]
Taylor, G.J. (1988) -The physiology of aluminium phytotoxicity. In: Sigel, H. and Sigel, A. (Ed.) – Metal ions in biological systems: Volume 24: Aluminium and its role in biology. New York, Marcel Dekker, p. 123-163. [ Links ]
Wong, M.T.F.; Akyeampong, E.; Nortcliff, S.; Rao, M.R. and Swift, R.S. (1995) -Initial responses of maize and beans to decreased concentrations of monomeric inorganic aluminium with application of manure or tree prunings to an Oxisol in Burundi. Plant and Soil, 171, 2: 275-282. [ Links ]
Vavilov, N.I. (1987) – A origem e geografia das plantas agrícolas. Moscovo, Nauka, p. 440. [ Links ]
Recebido/Received: 2011.11.05 Aceitação/Accepted: 2012.06.05

